










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.49 n.3-4 San José Dec. 2001
in neotropical agricultural landscapes during the dry season
Heather E. Williams 1,2 and Christopher Vaughan 1,3,4
1 Associated Colleges of the Midwest, Apdo. 10265, San Jose, Costa Rica
2 School of Veterinary Medicine, University of Wisconsin, Madison, WI 53706. MrsSoupah@aol.com
3 Regional Wildlife Program, Universidad Nacional, Heredia, Costa Rica.
4 Institute of Environmental Studies, University of Wisconsin, Madison, WI 53706. Fax 608-262-0014; cvaughan@facstaff.wisc.edu
Received 12-IX-2001. Corrected 26-IV-2001. Accepted 30-VIII-2001.
Abstract
Habitat use by a C. capucinus troop was studied in an agricultural landscape during late dry season (March-April 1994) in northwest Costa Rica. Riparian forests, palm canals and living fence rows accounted for 82 % of observations, significantly more than the other six habitats present. The study troop consumed 24 species of plants and five animals. Feeding concentrated on the introduced African oil palm (Elaeis guineensis) (33.6 %) and mango (Mangifera indica) (27.2 %), found mostly in palm canals and mango orchards respectively. The troop rested between 0930-1330 hr and fed and moved between 0530-0930 hr and 1330-1730 hr. Living fence rows were used as travel routes or corridors and less intensively for other activities.Key words: African oil palm, agricultural land, capuchins, Cebus capucinus, diet, habitat use, living fence rows.
Over 70 % of the original Central American forests have been converted into forest fragments separated by agricultural and urban landscapes, threatening many wildlife species (Cornelius 1991, Vaughan 1993). The white-faced monkey (Cebus capucinus) is considered an endangered species throughout most of its range in Central America due to habitat destruction and overhunting (Carrillo and Vaughan 1994). However, this species survives in altered habitats, including agricultural landscapes. We studied the late dry season habitat use, activity patterns, diet and movement of a C. capucinus troop in an agricultural landscape in northwest Costa Rica. We also documented the current wildlife management practices utilized there. Our objective was to utilize the results of the study and management practices to predict how healthy populations of white-faced monkeys could be maintained under similar circumstances.
Materials and methods
Study site and population: Curu Wildlife Refuge (CWR) is a privately owned wildlife refuge and farm located on the southeastern tip of the Nicoyan Peninsula in Puntarenas, Costa Rica (9 o 45 to 9 o 48 N and 84 o 50 to 84 o 57" E) (Fig. 1). It receives 1 600 mm annual precipitation, 90 % falling between May and November (Anonymous 1985). CWR extends over 1 492 ha with 30 % altered environments (pastures, living fencerows, forest plantations and fruit plantations) and 70 % natural forest environments (natural upland dry, lowland evergreen, mangrove and beach- marine) (Schutt and Vaughan 1995). Natural riparian forests (RF), palm canals (PC), and living fence rows (LFR) form vegetation corridors in the agricultural landscape. Ecotourism, cattle, and fruit production provide 90 % of the income for CWR. Because the abundant wildlife in CWR attracts many researchers and tourists, long-term objectives of its owners include combining wildlife/biodiversity conservation and agricultural development (Schutt and Vaughan 1995).
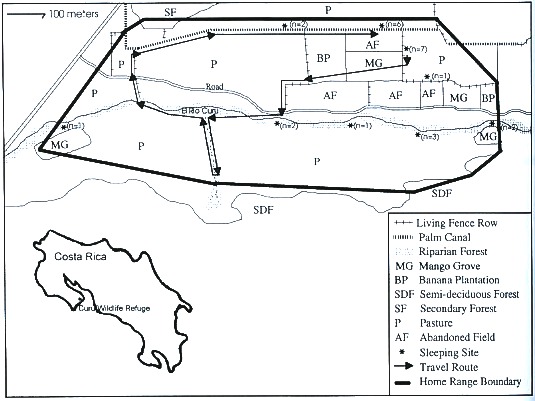
home range of the C. capucinus troop.
Estimated population sizes of 250 white-faced monkeys (17 individuals/km2 ) and 200 howler monkeys (Alouatta palliata) (13 individuals/ km2 ) are high for an area the size of CWR. Populations are probably large because CWR owners enforce a strict no-hunting policy and actually stimulate non-human vertebrate wildlife populations by providing artificial foods (bananas, mangos and coconuts) and allowing wildlife access to all habitats. White-faced monkeys were found in most of the habitats in CWR (A. Schutt pers. comm.). Our study troop inhabited the lowland farm area of CWR, dominated by pasture (P), mango groves (MG), banana plantation (BP), riparian forests (RF), living fence rows and palm canals. The study troop contained 34 monkeys (9 juveniles, 8 young adults and 17 adults). At least eight adults were female.
Research: It was conducted from dawn to dusk (0530-1730 hr) between March-April 1994. Using scan sampling of the nearest individual (Altmann 1974), activity of the nearest monkey to the observer was recorded at 5 min intervals.
Activities: They were classified as: a) moving (traveling through trees without feeding or foraging), b) foraging (actively searching for food), c) feeding (chewing or swallowing food), or d) other (playing, grooming, sleeping, sitting, fighting, copulating, or washing fur with lime fruit). A chi-square was used to analyze differences in feeding, foraging, moving, and resting activity along LRF, PC, and RF.
Home range: It was determined by the minimum convex polygon method (Mohr 1947, Hayne 1949).
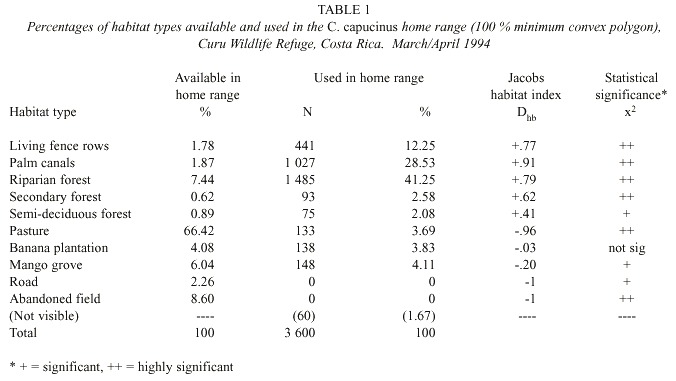
Habitat use: Habitat use (LFR, PC, RF, MG, BP, semi-deciduous forest (SDF), secondary forest (SF), road, abandoned field (AF) and pasture (P)) was recorded every 5 min. To analyze habitat use in proportion to its availability, gross cover type was mapped using aerial photographs (1:1 000, Instituto Geográfico Nacional 1985) and ground surveys. Percent use of each cover type available in the study troops home ranges was determined by overlaying a grid (16 dots/.25 2 ") on the cover map, counting points found in each cover type, and converting these numbers to percentages. The habitat use index (Dhb ) was used to determine cover preferences with the formula Dhb = (r-p)/(r+p-2rp) (Jacobs 1974), where r was the proportion of observations in a specific cover type, and p the proportion of that vegetative type available. Dhb values from –1 to 0 indicated a cover type used in lower proportion than available, while values from 0 to 1 indicated a cover type used in greater proportion than available. Expected and observed daytime locations were used to perform a chi-square goodness of fit test.
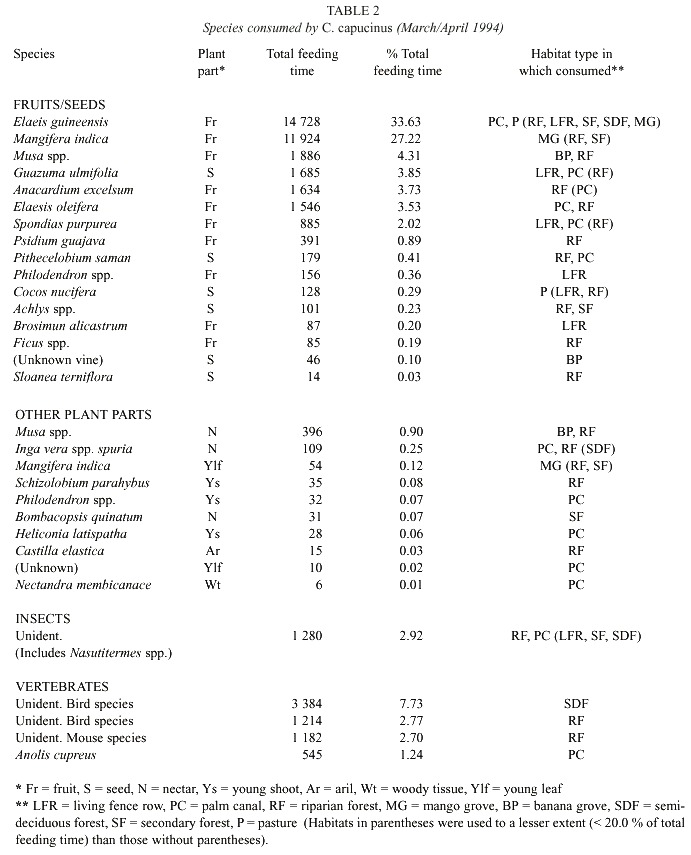
Diet: Feeding time duration was recorded with a stopwatch concurrently with the scan samples. When the focal monkey consumed a plant or animal, the species name and plant parts (seed, fruit, young leaf, young shoot, aril, nectar, or woody tissue) were recorded. Unknown species were collected for identification. Total feeding times on vegetation, insects, and vertebrates by the C. capucinus troop were recorded, and time feeding on each species and between habitats was compared.
Movement: Daily troop movement was mapped, recording site and time of directional change. Daily distances traveled were measured to obtain an average day range length.
Nocturnal sleeping sites: They were located by following the troop to the sleeping tree at sunset (1730 hr) and mapping the site on a study area map.
Results
Research was conducted between 0530 and 1730 hr for 25 days during March-April 1994.
Activity: The study troop spent 26.9 % of total observations moving, 26.5 % feeding, 16.3 % foraging, and 30.3 % in other activities (other activities were dominated by sleeping and will hereafter be referred to as "resting" activities). They rested more between 0930-1330 hr, fed more between 0530-0930 hr and 1330-1730 hr, and moved more between 0530-0930 hr and 1330-1730 hr. Comparing activities in LFR, PC and RF, the troop was more likely to move in LFR (x2 = 45.6, d.f. = 2, p < 0.01), to feed and rest in PC and RF (x2 = 12.5 and 27.8, respectively, d.f. = 2, p < 0.01), and to forage in RF (x2 = 13.7, d.f. = 2, p < 0.01).
Home range: A total of 3 600 observations were used to estimate 100 % home range. The study troop occupied a home range of approximately 37.2 ha (Fig. 1).
Habitat use: Compared to availability was based on 100 % home range size and 3 600 daytime observations over a two-month span. The study troop used habitat types in proportions different from their availability in the home range. The habitat use index showed LFR, PC, RF, SF and SDF used in a higher proportion than available, and P, MG, road and AF used in a lower proportion than available. Banana plantations were used in the same proportion as available (Table 1). The troop spent most time in LFR (0530-0630 hr), PC (0630-0830 hr, 1530-1730 hr) and RF (0830-1530 hr). LFR, PC and RF in the home range area were approximately 800 m, 700 m and 1 500 m long respectively and varied around 1 m, 4 m and 13 m in width respectively.
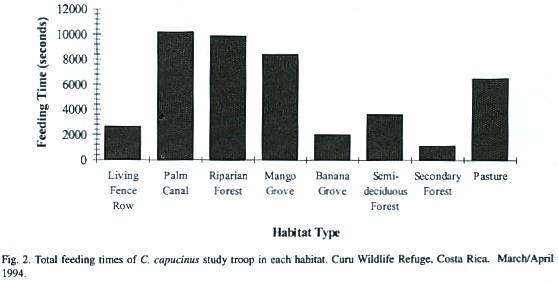
Diet: The study troop consumed 24 species of plants, including four agricultural plants: African oil palm fruit (Elaeis guineensis), mango (Mangifera indica), coconut (Cocos nucifera), and bananas (Musa sp.). They also ate several insect species, three unidentified bird species and a lizard (Table 2). Feeding on African oil palm fruit (31.7 %) and mango fruit (25.0 %) was greater than on other foods. The troop fed more in PC, RF and MG than other habitat types (Fig. 2), consuming especially E. guineensis and E. oleifera in PC, M. indica in MG, and Anacardium excelsum in RF. They also less frequently consumed Spondias mombin fruits and Guazuma ulmifolia seeds along LFR.
Movement: Based on 25 days of observation, the study troop averaged 1 290 m daily (range = 936 m to 1 550 m, S.D. = 200 m) (Fig. 1).
Nocturnal sleeping sites: Nine nocturnal sleeping sites were identified during the study period. LRF, PC and RF were used almost equally as sleeping sites (Fig. 1).
Discussion
The dry season is a difficult period for wildlife species because of potential water and food shortages (Chapman 1988, Chapman and Fedigan 1990, Moscow and Vaughan 1987, Vaughan et al. 1997). Man-planted food sources (African oil palms, mangos, and bananas) during the late dry season turn agricultural areas into important feeding areas for wildlife species (Potus flavus, Procyon lotor, Nasua narica, Odocoileus virginianus), including Cebus (Schutt and Vaughan 1995). Several Cebus troops from surrounding forested environments also feed on these agricultural crops.
Although this was only a two month study in the late dry season, several trends were obvious. The study troop entered semi-deciduous forest and secondary forest habitats only about 2 % each of total observations during the two-month study. For over 82 % of our observations, the troop was in narrow tree strips or corridors of LFR (12.2 %), PC (28.5 %), and RF (41.3 %).
During important feeding hours (0630-0830 hr, 1530-1730 hr), PC (22.9 %), RF (22.2 %), MG (19.0 %) and P (14.6 %) habitats were the most utilized. LFR (6.0 %) was used slightly less than semi-deciduous forest (8.2 %), and more than BP (4.6 %) and SF (2.5 %). The study troop spent more time in PC than other habitats because of the abundant E. guineensis present. This palm is found less commonly in other habitat types (P, SF, RF and LFR). RF provided the greatest diversity of native food species (Table 2). Chapman and Fedigan (1990) studied C. capucinus feeding patterns for three field seasons (January-July) in a national park in northwest Costa Rica.
They found feeding times on plants between three troops varied between 53-81 % of total feeding time, with the remainder dedicated to insects. Only Sloanea terniflora and Ficus sp. were utilized in both studies; S. terniflora was consumed extensively in Chapman and Fedigans (1990) study and only marginally in ours. Agricultural plants (mangos, African oil palm, and bananas) in our study probably replaced many plant species found by Chapman and Fedigan (1990).
RF was the habitat most used during the hottest hours (0930-1330 hr), probably because it provided a shady, moist protected resting site along the Curu river. Early morning movement, morning and late afternoon feeding, and midday resting were also observed for C. capucinus in western Costa Rica during the late dry season (Moscow and Vaughan 1987). Preliminary observations indicate that a similar pattern of troop use of LFR, PC, and RF exists at other periods of the year.
Although marginally used for feeding (6.0 %) and overall observations (12.2 %), LFR was used more than its availability (Table 1). The troop usually traveled along a LFR to reach feeding trees in PC, RF, MG or P. LFR consisted primarily of S. mombin, Bombacopsis quinatum, and G. ulmifolia. Only G. ulmifolia was used as a food source during the study. The most common sleeping site (n = 7 nights) was found in a LFR, between MG, P and AF. In addition, LFR and PC were used as travel routes to individual trees of C. nucifera, E. guineensis and M. indica, all found in P. On one occasion, a Cebus individual traveled 60 m to feed at an isolated C. nucifera. Pastures accounted for 66.4 % of the home range, but only 3.7 % of observations. Isolated E. guineensis and M. indica trees in pastures accounted for 30 % and 16 % of pasture feeding observations respectively.
The 1 290 m average daily distance (ADT) traveled by the study troop was less than the 4 500 m ADT observed for C. capucinus in Palo Verde (Moscow and Vaughan 1987), 1 746-3 469 m ADT for C. apella in French Guyana by Zhang (1995), and 2 000-4 000 m ADT for C. olivaceus by Ruiter (1986). We agree with Zhang (1995) that troop feeding and ranging patterns were responses to changes in fruit availability and distribution. In CWR, though the troop used LFR, PC, and RF corridors extensively more than their availability, limited daily movement indicates concentrated agricultural food sources in a small area provided all needed food resources. Eight of the nocturnal sleeping sites were within 200 m of each other, and most were bordered by pasture.
LFR, PC, and RF are important habitats for the CWR study troop during the latter part of the dry season, providing travel routes between habitats, protection, and food sources. The troop preferred these habitats to the other six habitats in the study area. Considering reluctance of C. capucinus to travel long distances through open pasture, travel between habitats without these conduits seems unlikely. Our first management recommendation is plant more conduits with sufficient density, width, tree species and food resources to provide a safe passage. This will probably increase C. capucinus populations on agricultural landscapes.
CWR maintains wildlife species because it has a non-traditional approach to agricultural production and a fervent interest in wildlife. Management policies which favor C. capucinus at CWR includes: a) enforcing a strict non-hunting policy, b) maintaining LFR and PC as food sources and travel routes for wildlife (other farms use metal or treated wooden fence posts), c) maintaining E. guineensis, E. oleifera and banana varieties in PC, P, RF and BP for wildlife and cattle food, d) permitting C. capucinus and other wildlife species to consume coconut, mango and banana production, and e) maintaining natural vegetation adjacent to agricultural lands. Our short study indicates that these guidelines are beneficial. Because over 50 % of CWRs present income is generated from ecotourism and non-human primates are major attractions (Schutt and Vaughan 1995), this wildlife management plan (protection, living fence rows, and artificial food) should be continued. Agricultural loss is compensated for by ecotourism. In addition, studies in CWR have shown that capuchins foraging in the mangrove orchards eat insects that damage mango crops (A. Schutt pers. comm.). Intelligent management of LFR will ensure free movement of primates (Lindenmayer and Nix 1993). We believe that many wildlife species could benefit from a positive attitude from their landowners and by following some habitat and water management principles.
Acknowledgments
We thank the Schutt family, owners of CWR for their hospitality and assistance, Judy Magnan for computer instruction, Michael McCoy for statistical analysis, and Karl DeLong and Adrian Treves for helpful comments on this manuscript.
Resumen
Se estudió el uso de hábitat por una tropa de C. capucinus en una zona de agricultura durante la estación seca tardía (Marzo-Abril 1994) en el noroeste de Costa Rica. Los bosques riparios, canales de palmas y cercas de árboles vivos contaron con el 82 % de las observaciones, significantemente más que los otros seis hábitats presentes. La tropa de estudio consumió 24 especies de plntas y cinco animales. La alimentación se concentró en la palma de aceite Africana introducida (Elaeis guineensis) (33.6 %) y en mango (Mangifera indica) (27.2 %), encontrados principalmente en los canales de palmas y huertos de mango respectivamente. La tropa descansó entre las 0930-1330 hr y se alimentó y movió entre las 0530-0930 hr y 1330-730 hr. Las cercas de árboles vivos fueron usadas como rutas de paso o corredores y menos intensamente para otras actividades.
References
Altmann, J., 1974. Observational study of behavior: Sampling methods. Behavior 49: 227-265. [ Links ]
Anonymous. 1985. Catastro de las series de precipitaciones medidas en Costa Rica. Instituto Meteorológico Nacional, San José, Costa Rica. 445 p. [ Links ]
Carrillo, E. & C. Vaughan. 1994. La vida silvestre de Mesoamerica: Diagnóstico y estrategia para su conservacion. EUNA, Heredia, Costa Rica. [ Links ]
Chapman, C. 1988. Flexibility in diets of three species of Costa Rican primates. Folia Primatol. 49: 90-105. [ Links ]
Chapman, C. & L. Fedigan. 1990. Dietary differences between neighboring Cebus capucinus groups: Local traditions, food availability or responses to food profitability? Folia Primatol. 54: 177-186. [ Links ]
Cornelius, S. 1991. Wildlife conservation in Central America: Will it survive the 90s? Trans. 56 th N. A Wildl. Natur. Resour. Conf. 56: 40-49. [ Links ]
Hayne, D.W. 1949. Calculations of size of home range. J. Mammal. 30: 1-18. [ Links ]
Jacobs, J. 1974. Quantitative measurement of food selection. Oecologia 14: 413-417. [ Links ]
Lindenmayer D.B. & H.A. Nix. 1993. Ecological principles for the design of wildlife corridors. Cons. Biol. 7: 627-630. [ Links ]
Mohr, C.O. 1947. Table of equivalent populations of North American small animals. Amer. Midland Natur. 37: 223-249. [ Links ]
Moscow, D. & C. Vaughan. 1987. Troop movement and food habits of white-faced monkeys in a tropical-dry forest. Rev. Biol. Trop. 35: 287-297. [ Links ]
Ruiter, J.R. de. 1986. The influence of group size on predator scanning and foraging behavior of wedge-capped capuchin monkeys (Cebus olivaceus). Behaviour 77: 240-258. [ Links ]
Schutt, A. & C. Vaughan. 1995. Incorporating wildlife into development: The case of the Curu Wildlife Refuge and Farm, Costa Rica, p. 250-254. In J. Bissonette & P. Krausmann (eds.). Integrating people and wildlife for a sustainable future. The Wildlife Soc., Bethesda, Maryland. [ Links ]
Vaughan, C. 1993. Human population and wildlife: A Central American focus. Trans. 58th N.A. Wildl. Natur. Resour. Conf. 55: 129-136 [ Links ]
Vaughan, A., J. Cabrera, D. Moore & C. Vaughan. 1997. Water loss at twenty natural waterholes in Guanacaste National Park, Costa Rica. Rev. Biol. Trop. 45: 1681-1684. [ Links ]
Zhang, S. 1995. Activity and ranging patterns in relation to fruit utilization by brown capuchins (Cebus apella) in French Guyana. Int. J. Primatol. 16: 489-507. [ Links ]

