










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.49 n.3-4 San José Dec. 2001
Monobia schrottkyi (Hymenoptera, Vespidae, Sphecidae)
Evandro Camillo
Departamento de Biologia, Faculdade de Filosofia Ciências e Letras de Ribeirão Preto-USP, 14040-901, Ribeirão Preto, SP, Brasil. Fax: 55 016 602 3666. E-mail: ecamillo@ffclrp.usp.br
Received 12-VII-2000. Corrected 11-XII-2000. Accepted 08-XI-2001.
Abstract
Key words: Eumeninae, Sphecidae, inquilinism, Monobia, Montezumia, Podium, Trypoxylon.
Several species of solitary wasps utilize previously existing cavities for nest construction, including the abandoned mud nests constructed by sphecids, which provide a large number of cavities (open and empty cells). Among them, several species belonging to the genera Monobia and Montezumia (Vespidae:Eumeninae), Trypoxylon and Podium (Sphecidae) have been found occupying such abandoned nests (Rau 1940, Krombein 1967, Freeman and Jayasingh 1975, Fritz and Genise 1980, Willink 1982, Genaro 1994).
Monobia and Montezumia are Neotropical, most species being present in the moist forests of South America, yet some species occur in the Nearctic region (Willink 1982). Monobia schrottkyi Bertoni, 1918 occurs in northern Argentina, Paraguay and Brazil; Monobia curvata Fox, 1899 occurs in Bolivia and Brazil, and Montezumia petiolata Saussure, 1855 is distributed from Venezuela and Colombia to northern Argentina (Willink 1982). According to Bohart and Menke (1976), Podium is primarily a Neotropical genus, occurring in
the eastern half of North America where one species ranges as far north as New York; P. denticulatum Smith, 1856 is found from Mexico to Brazil. Trypoxylon is a cosmo-politan genus, but poorly represented in Australia and the various Pacific island groups; T. rogenhoferi Kohl, 1884 occurs in Argentina and Brazil (Bohart and Menke 1976).
The purpose of this paper is to present biological data on M. schrottkyi, M. curvata, M. petiolata, T. rogenhoferi and P. denticulatum as inquilines of Brachymenes dyscherus Saussure, 1852 nests.
Materials and methods
Study site: The study was carried out in four locations at Fazenda Santa Carlota, Cajuru, São Paulo, Brazil (21° 17 – 18 S, 47° 12 – 18 W). The study area covers 6 300 ha, with altitudes ranging from 540 to 944 m, and it is drained by several streams. Of the total area, approximately 2 200 ha is covered by native vegetation preserved in isolated patches or gallery forests (semideciduous forests, riparian forests, cerrados and cerradões) surrounded by cultivated land and/or pastures. There are some areas at various stages of secondary growth of the main vegetation types. The predominant soils are Rhodic Ferralsol and Orthic Ferralsol (Oliveira et al. 1983). The study area contained various abandoned rural buildings, surrounded by pastures, plantations (mainly sugar cane), and abandoned orchards.
Two seasons, distinguished by tempera-ture and precipitation, are recognized in this region: 1- Cold and dry season (May to August) with monthly average temperatures ranging from 18 to 22 °C and precipitation from 0 to 81.7 mm; 2- Hot and wet season (September to April) with monthly average temperatures ranging from 19.9 to 26.1 °C and precipitation from 35.3 to 422.6 mm.
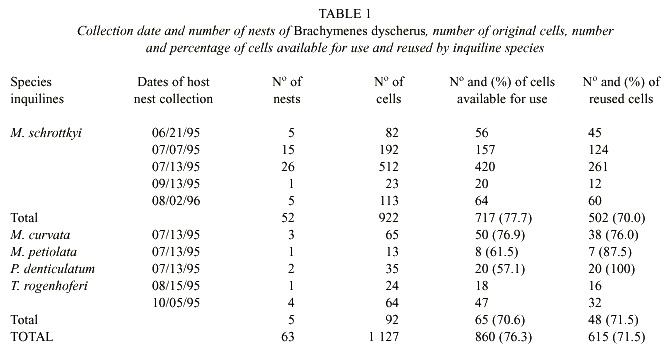
Nests: Sixty-three inactive nests of B. dyscherus with reused cells (5 nests on 6/21/95, 15 on 7/7/95, 31 on 7/13/95, 2 on 8/15/95, 1 on 9/13/95, 4 on 10/5/95 and 5 on 8/2/96) were collected from the walls inside abandoned houses, using a pallet knife. The closure plugs, generally constructed with mud of different colors or presenting a little depression, helped identify the reused cells. In the P. denticulatum cells the closure plugs were polished and shiny. When, as happened in some nests, the immatures from the first layer in contact with the substrate were exposed, they were collected and placed in labeled individual vials and identified. In the laboratory all nests were placed in plastic boxes and together with the vials were kept at room temperature until the adults emerged. After emergence the nests were opened to analyze their cells. All the wasps are deposited in the Entomology Collection of Biology Department-FFCLRP-USP.Nests of Brachymenes dyscherus: The nests were constructed with mud on the interior walls of abandoned houses located at least 1 m from the floor, in dry and shaded places. They were composed of 1 to 4 layers, with cells constructed in two parallel series. Once completed, the nests were wholly covered with mud, resulting in a distinctive shape. The surface was rough and in some cases it was impossible to distinguish individual cells. The number of cells per nest ranged from 3 to 62, and they were provisioned with larvae of Lepidoptera (from 4 to 29 preys). In nests with 4 layers, the maximum number of cells counted was 23 for the first layer, 20 for the second, 16 for the third, and 5 for the fourth. The cocoons, milkish white and more or less transparent, were affixed to the cell walls; remains of prey together with larval feces were found adhered to the external surface of the cocoons. Brachymenes dyscherus is a univoltine species whose immatures undergo prepupal diapause. The development period ranges from 158 to 309 days for males, and from 167 to 320 days for females. Most larvae develop into adults during the same year of nest construction, with the greatest number of emergences occurring from September to December, although some individuals had much longer development periods (from 343 to 595 days). The sex ratio was 1.5 males to 1 female (Camillo 1999).
Results
In nests (n = 63) of B. dyscherus, of 1 127 original cells there were 860 (76.3 %) available for use, of which 615 (71.5 %) were reused by inquiline species. Of these nests 52 were occupied exclusively by M. schrottkyi, 3 by M. curvata, 1 by M. petiolata, 2 by P. denticulatum and 5 by T. rogenhoferi. Cells not reused (sealed) contained old material from the first generation. The percentage of available cells, in relation to total number of cells in the nests of each species and those reused, in relation to available cells were statistically similar (X2 = 4.39 and X2 = 7.27, gl = 4, p < 0.05, respectively). Correlation among these percentages was negative and significant (r = 0.91, p < 0.05) (Table 1). Cells of B. dyscherus available for use and reused by inquilines can be observed in the Fig. 1.

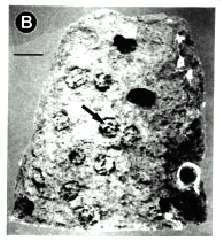
Fig. 1. A - Nest of Brachymenes dyscherus with several cells available for use; B - Reused
cells showing the closure plugs. The lines correspond to 1.0 cm.
Only 3 nests of B. dyscherus were used by two different inquiline species. One of these, collected on 7/13/95 with 28 original cells and 21 available, was used by P. denticulatum and M. schrottkyi. The first species reused 10 cells in which 2 dead females and 8 dead immatures were found; the second one reused only 2 cells, from which 1 male (11/8/95) and 1 female (12/23/95) emerged. Parancistrocerus areatus Fox (two individuals) and T. rogenhoferi emerged from the other two nests. The data from the first case and from P. areatus is not reported in the tables and figs.
Monobia schrottkyi
This species used 52 nests of B. dyscherus, which had 922 cells; among the 717 available for use 502 were reused (Table 1). The number of reused cells per nest varied from 3 to 24, with most nests having from 6 to 11 cells (Fig. 2). The number of individuals emerged per nest ranged from 1 to 16 with most nests having from 1 to 6 individuals (Fig. 3).
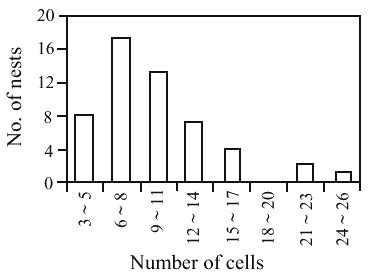
schrottkyi in nests of Brachymenes duscherus
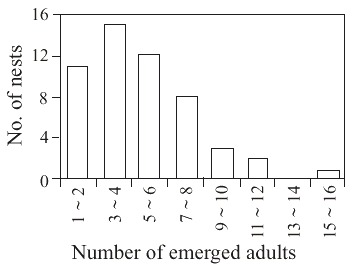
schrottkyi emerged from nests of Brachymenes duscherus
In total, 258 individuals of M. schrottkyi emerged from September (9/4/95) to April (4/2/96) (hot and wet season), with the greatest frequency occurring in January (Fig. 4). Monthly numbers of these emergences, according to the date of collection of the B. dyscherus nests, are presented in Table 2. It is worth emphasizing that eigth individuals were found dead inside the cells. All immatures were in prepupal diapause when the nests were collected.
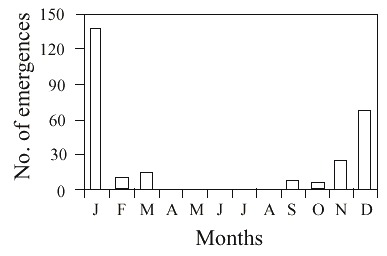

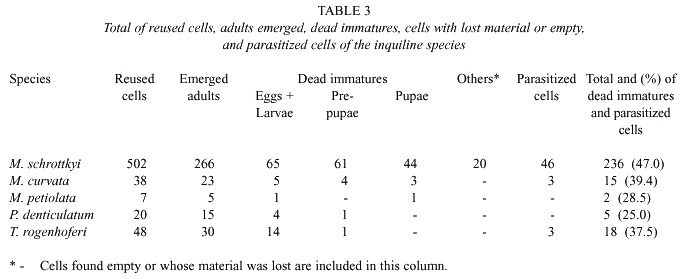
The sex ratio, determined for the total population produced, independently of the season of nests collection, was 154 males to 112 females, which was significantly different from 1:1 (X2 = 6.32, gl = 1, p < 0.05). Mortality in M. schrottkyi and in the other four inquilines species was statistically similar (X2 = 4.3, gl = 4, p > 0.05). Among the reused cells, 44 were parasitized by Melittobia sp. (Eulophidae) and 2 by Ichneumonidae (Table 3).
Monobia curvata
This species used 3 nests of B. dyscherus, which had 65 cells. Among the 50 available for use, 38 were reused (Table 1). The number of reused cells per nest was 8, 15 and 15, of which emerged 5, 7 and 11 individuals, respectively. The emergences started in the final part of the cold and dry season, in August (8/28/95) (n = 2) extending to September (n = 10), October (n = 6) and November (until 11/19/95) (n = 3). In addition, one individual was dead inside the cell and for another, the emergence date was not determined. The sex ratio was 15 males to 8 females, which was not statistically different from 1:1 (X2 = 1.56, gl = 1, p > 0.05). Among the reused cells, three were parasitized by Ichneumonidae (Table 3). When the nests were collected, all immatures were in prepupal diapause.
Montezumia petiolata
This species used 1 nest of B. dyscherus with 13 cells, of which 8 were available for use. Of these, 7 were reused (Table 1), and 5 individuals (2 males and 3 females) emerged in September (from 9/2 to 9/11/95). When the nests were collected all immatures were in prepupal diapause.
Podium denticulatum
This species used 2 nests of B. dyscherus with 35 cells, which had 20 cells available for use. Of these, 5 in one nest and 15 in another were reused (Table 1). In August (from 8/14 to 8/27/95) 1 and 12 individuals emerged, respectively. In each nest 1 adult was dead inside the cell. The sex ratio was 11 males to 4 females, which is not significantly different from 1:1 (X2 = 2.40, gl = 1, p > 0.05).
Trypoxylon rogenhoferi
This species used 5 nests of B. dyscherus that had 92 cells. Of these 48 were reused among the 65 available for use (Table 1). In the nests 3, 8, 10, 11 and 16 cells were reused, from which 2, 2, 5, 9 and 12 individuals emerged, respectively. The emergences from nests collected on 8/15/95 occurred in September (from 9/8 to 9/16/95) (n = 13), and for those collected on 05/10/95, the emergences occurred in October (10/31/95) (n = 2) and November (until 11/19/95) (n = 15). The sex ratio was 19 males to 11 females, which was not statistically different from 1:1 (X2 = 1.62, gl = 1, p > 0.05). Three of the reused cells were parasitized, producing individuals from the families Ichneumonidae (n=2) and Chrysididae (n = 1).
Discussion
Iwata (1976) classified the nesting habits of solitary Vespidae into three categories: 1-borrowers that excavate nests in the soil; 2-builders that construct their entire nests from materials such as mud or masticated plant material and 3- renters that occupy and modify preexisting cavities. These are opportunistic and occupy a wide variety of cavities such as the pith of twigs, cavities in dead plants, or cells of abandoned mud dauber nests (Cowan 1991).
Species classified as inquilines can occupy and manipulate cells of old mud nests made by other solitary wasps. The mud used in these nests is quite durable and does not wash away in rain, perhaps because of the addition of saliva to the water and soil which would strengthen the nest (Isely 1913), as has been suggested for a sphecid (Qureshi and Ahmad 1978 in Cowan 1991). According to Genaro (1994), the nests of Sceliphron spp., are highly durable and can remain intact for 7 years (Freeman 1974). Nests of Sceliphron spp. are similar to those of B. dyscherus studied in this work, and are reused by several Vespidae and Sphecidae (see for example Krombein 1967, Freeman 1973, Willink 1982, Genaro 1994).
The inquilines studied in this work used abandoned nests of B. dyscherus because their open cells serve as suitable cavities. Thus, these species presented similar behavior to those that nest in preexisting cavities. For some species the inquilinism here observed seems to be facultative, because in the vicinity, and during the same period, nests of T. rogenhoferi (Camillo et al. 1994), P. denticulatum (Camillo et al. 1996) and Monobia angulosa Saussure, 1852 (Camillo et al. 1997) were obtained from bamboo cane trap-nests.
According to Evans (1973) the genus Montezumia could have species that nest in the soil (M. infernalis (Spinola, 1851), M. nigriceps (Spinola, 1841)) and others which construct free mud nests (M. ferruginea, M. dimidiata Saussure, 1852, M. cortesioides Willink, 1982). Nevertheless and according to Willink (1982) this classification should be reconsidered, because the only data on species that construct nests in soil correspond to those two previously mentioned and this is impossible to confirm. Previously, Strand (1912 in Willink 1982) observed M. infernalis constructing free nests or nests in wall cavities, but not in the soil.
Recent observations have been made in relation to species that could construct free nests: M. ferruginea, M. dimidiata, M. cortesioides, M. vechti Willink, 1982, M. pelagica Saussure, 1852, M. brethesi Bertoni, 1918 and M. platinia Saussure, 1852 (Willink 1982). The possibility that they were inquilines, as observed by Fritz and Genise (1980) in M. ferruginea, was not mentioned in any of these cases; nevertheless the majority of the mentioned cases of mud nests could correspond actually to species that use abandoned nests of other wasps (in Willink 1982). As observed in this study, M. petiolata is an inquiline of B. dyscherus nests.
Acknowledgments
I am especially grateful to the owners of Santa Carlota farm, Cajuru, Brazil for allowing us free access to their land. I thank James M. Carpenter (American Museum of Natural History) for identifying the Monobia and Montezumia species, Carlos Alberto Garófalo for their comments and suggestions, and José Carlos Serrano for technical assistance.Resumen
Se recolectaron 64 nidos inactivos de la avispa unta-dora de barro solitaria Brachymenes dyscherus reusados por 5 especies inquilinas en Fazenda Santa Carlota, Caju-ra, São Paulo, Brasil en 1995 y 1996. Monobia schrottkyi usó 52 nidos; entre las 717 celdas disponibles, 502 fueron reusados. El número de celdas por nido varió de 3 a 24; de 1 a 16 individuos emergieron desde setiembre hasta abril (154 machos y 112 hembras). Cuarenta y seis celdas fue-ron parasitadas por Melittobia sp. (n = 44) e Ichneumoni-dae (n = 2). Monobia curvata usó 3 nidos; entre las 50 cel-das disponibles, 38 fueron reusadas y 15 machos y 8 hembras emergieron desde agosto hasta noviembre. Tres celdas fueron parasitadas por Ichneumonidae. Montezumia petiolata ocupó un nido; entre las 8 celdas disponibles, 7 fueron reusadas y 2 machos y 3 hembras emergieron en setiembre. Podium denticulatum usó 2 nidos; las 20 celdas disponibles fueron reusadas y 11 machos y 4 hembras emergieron en agosto. Trypoxylon rogenhoferi usó 5 nidos que tuvieron 65 celdas disponibles; 48 fueron reusadas, de las cuales emergieron 19 machos y 11 hembras desde se-tiembre hasta noviembre. Tres celdas fueron parasitadas por Ichneumonidae (n = 2) y chrysididae (n = 1).
References
Bohart, R.M. & A.S. Menke. 1976. Sphecid wasps of the world. University of California, Berkeley. 695 p. [ Links ]
Camillo, E. 1999. Asolitary mud-daubing wasp Brachymenes dyscherus (Hymenoptera, Vespidae, Eumeninae), from southeastern Brazil with evidence for a life-cycle poliphenism. Rev. Biol. Trop. 47: 949-958. [ Links ]
Camillo, E., C.A. Garófalo & J.C. Serrano. 1994. Observações sobre a biologia de Trypoxylon (Trypargilum) rogenhoferi Kohl (Hymenoptera: Sphecidae). Ann. Soc. Entomol. Brasil 23: 299-310. [ Links ]
Camillo, E., C.A. Garófalo, J.M.F. Assis & J.C. Serrano. 1996. Biologia de Podium denticulatum Smith em ninhos armadilhas (Hymenoptera: Sphecidae: Sphecinae). Ann. Soc. Entomol. Brasil 25: 439-450. [ Links ]
Camillo, E., C.A. Garófalo & J.C. Serrano. 1997. Biologia de Monobia angulosa Saussure em ninhos armadilhas (Hymenoptera: Vespidae: Eumeninae). Ann. Soc. Entomol. Brasil 26: 169-175. [ Links ]
Cowan, D.P. 1991. The solitary and presocial Vespidae, p. 33-73. In K.G. Ross & R.W. Matthews (eds.). The social biology of wasps. Cornell Univ., Ithaca, New York. [ Links ]
Evans, H.E. 1973. Notes on the nests of Montezumia (Hymenoptera, Eumenidae). Entomol. News 84: 285-290. [ Links ]
Freeman, B.E. 1973. Preliminary studies on the population dynamics of Sceliphron assimile (Dahlbom) (Hymenoptera: Sphecidae) in Jamaica. J. Anim. Ecol. 42: 173-182. [ Links ]
Freeman, B.E. 1974. The distribution in Jamaica of the mud wasp Sceliphron assimile (Dahlbom) (Sphecidae) and its associates. Caribbean J. Sci. 14: 115-124. [ Links ]
Freeman, B.E. & D.B. Jayasingh. 1975. Population dynamics of Pachodynerus nasidens (Latr.) in Jamaica (Hymenoptera: Eumenidae). Oikos 26: 86-91. [ Links ]
Fritz, M.A. & J.F. Genise. 1980. Notas sobre nidos de barro de Sphecidae (Hymenoptera). Constructores, inquilinos, parasitoides, cleptoparásitos y detívoros. Rev. Soc. Entomol. Arg. 39: 67-81. [ Links ]
Genaro, J.A. 1994. Inquilinos de Sceliphron assimile, com ênfasis en Podium fulvipes (Hymenoptera: Vespidae, Sphecidae, Megachilidae). Caribbean J. Sci. 30: 268-270. [ Links ]
Isely, D. 1913. The biology of some Kansas Eumenidae. Univ. Kansas Sci. Bull. 8: 235-309. [ Links ]
Iwata, K. 1976. Evolution of instinct: Comparative ethology of Hymenoptera. Amerind, New Delhi. 235 p. [ Links ]
Krombein, K.V. 1967. Trap-nesting wasps and bees. Life histories, nests and associates. Smithsonian Inst., Washington, D.C. 570 p. [ Links ]
Oliveira, V., A.M. Costa & W.P. Azevedo. 1983. Pedologia, folhas SF. 23/24, Rio de Janeiro. Projeto RADAMBRASIL, Levantamento de Recursos Naturais 32: 485-552. [ Links ]Rau, P. 1940. Some mud-daubing wasp of Mexico and their parasites. Ann. Entomol. Soc. Amer. 33: 590-595. [ Links ]
Willink, A. 1982. Revisión de los géneros Montezumia Saussure y Monobia Saussure. Bol. Acad. Nac. Cien. 55. Córdoba, Argentina. 321 p. [ Links ]

