










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.49 n.3-4 San José Dec. 2001
en playa Tucacas, Falcón, Venezuela
Alberto Martín, Yusbelly J. Díaz Díaz y Claudio Paredes
Universidad Simón Bolívar. Departamento Estudios Ambientales. Apartado 89000. Caracas 1086-A. Venezuela, fax: 58-2-906-3039, correo electrónico: amartinz@usb.ve
Recibido 3-II-2000 Corregido 17-XI-2000 Aceptado 14-II-2001
Abstract
Three hundred and twelve shells of Sanguinolaria cruenta were collected in May 1999 in a sandy beach in Tucacas, Falcón, Venezuela. For each shell, antero-posterior length (Lap), overall height (Al), wide (An), weight (Pe), valves area (Arv), anterior length (La) and dorsal height (Ad) were measured. The descriptive statistic of the studied individuals, allows to appreciate that the variables that are good as parameters for the taxonomic description of S. cruenta are: Lap, Al, An, La y Ad, given their little variability. The maximum class frequencies is 41-70 mm for Lap (91% of the population). Analysis of data determined relationships of Lap to An, Lap to La were isometric (Lap = 0.98078 An + 1.50456 and Lap = 0.98052 La + 5.25840, respectively), whereas the other relationships indicate allometric growth.
Key words: Morphometrics, Sanguinolaria cruenta, bivalve, Venezuela.
Sanguinolaria cruenta (Lightfoot 1786) sinónimo de S. operculata (Gmelin 1791), es un bivalvo de concha alargada-lanceolada con el margen posterior acuminado y de color rosado intenso a rojo, su valva derecha es fuertemente convexa y la izquierda más aplanada, de superficie lisa con numerosas líneas concéntricas (Abbott 1974, Rios 1994). Esta especie es rara y poco común en el Mar Caribe y la costa continental de Suramérica, a pesar de que a sido reportada desde la Península de Yucatán hasta el sur de Brasil (Abbott 1974, Robinson y Montoya 1987, Díaz y Puyana 1994, Rios 1994). Este trabajo constituye la morfometría básica y un nuevo registro de esta especie para la costa Centro-Occidental de Venezuela, la cual ha sido reportada solo previamente al SE de Higuerote (Tello 1975) y en los Cerros de Caigüire en el Oriente del país por Pérez Nieto (1965). Por otra parte, en las costas nororientales de Venezuela se encuentra también presente S. sanguinolenta, especie poco común asociada a fondos fangosos de aguas someras (Lodeiros et al.1999).
La importancia de este trabajo reside en que contribuye al conocimiento de la morfometría y alometría de Sanguinolaria cruenta, debido a que no existen trabajos previos sobre la misma, sino reportes de su existencia en las localidades antes señaladas.
Materiales y métodos
Un total de 312 especímenes de Sanguinolaria cruenta se extrajeron a principios de mayo de 1999 en Playa Tucacas, Edo. Falcón, Venezuela (10°45' 55" N y 68°19' 24" W), inmediatamente después de una mortandad masiva de moluscos ocurrida en la zona. Esta localidad corresponde a una playa arenosa disipativa, con pendiente muy suave y una zona de oleaje bastante desarrollada (Herrera 1995).
En el laboratorio, las conchas fueron lavadas y secadas, y en ellas se midieron las siguientes variables morfométricas: longitud antero-posterior (Lap), altura total (Al) y ancho de la concha (An) (Stanley 1970) (Fig. 1), con un vernier de 0.01 mm de apreciación. También fueron pesadas (Pe) con una balanza analítica de 0.01 g de apreciación, removiendo previamente cualquier resto de tejido que se encontrará en ellas. Luego fueron desprendidas las valvas izquierdas, para determinar su área (Arv). Para estimar el Arv, primero se dibujó el contorno de las mismas en un papel de aluminio, que fue pesado y posteriormente convertido en área, al conocer el peso de un pedazo del mismo papel de área conocida. Para la determinación de la longitud anterior (La) y la altura dorsal (Ad) se fotocopiaron las valvas izquierdas, y en el papel se les dibujo una línea a lo largo del eje máximo antero-posterior y otra perpendicular a la primera, a través del umbo (Fig. 1). La longitud anterior fue definida como la distancia desde la intersección de estas líneas al margen anterior de la valva. Similarmente, la altura dorsal fue definida como la distancia desde la intersección al punto más externo del umbo (Donn 1990).
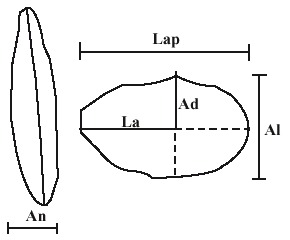
longitud anterior (La); altura dorsal (Ad) y ancho (An).
Fig. 1. Five of the morphometric variables measured. An-teroposterior length (Lap); total heigth (Al);
anterior length (La); dorsal heigth (Ad) and width (An).
Para describir la morfometría de los especímenes recolectados, se utilizaron variables descriptivas, análisis de correlaciones, regresión lineal y la distribución de tallas, siguiendo la metodología sugerida en Sokal y Rohlf (1979).
Resultados
La distribución de frecuencias para la longitud antero-posterior de la población analizada (Fig. 2), indica que las mayores frecuencias se encuentran entre los 56 mm y 60 mm con un 21.23% de individuos, un 16.09% menores de 45 mm, un 30.13% entre 45 mm y 55 mm y un 32.53% mayores de 60 mm.
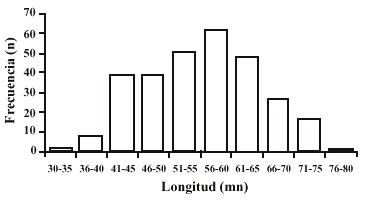
Fig. 2. Size of S. cruenta using the antero-posterior longitude (Lap).
La estadística descriptiva de los individuos estudiados (Cuadro 1), permite apreciar que las variables que sirven como parámetros para la descripción taxonómica de S. cruenta son: longitud antero-posterior (Lap), altura total (Al), ancho de la concha (An), longitud anterior (La) y altura dorsal (Ad), dada su relativamente baja variabilidad (coeficientes de variación entre 16.27 y 18.39). Por otra parte, el peso (Pe) y área de la valva (Arv) fueron las variables que presentan las mayores desviaciones estándar (Cuadro 1).
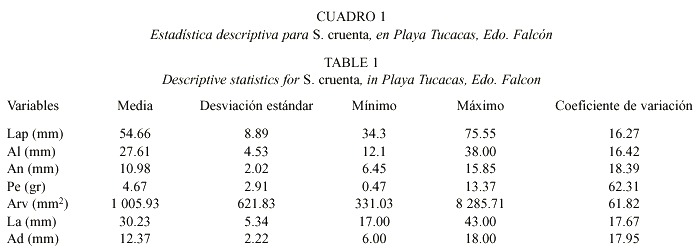
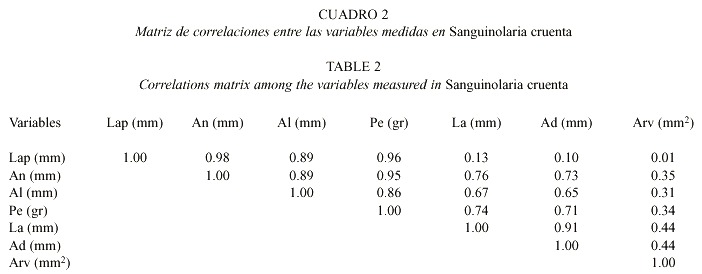
La matriz de correlaciones entre las variables medidas en los organismos de S. cruenta recolectados en Playa Tucacas, se observa en el Cuadro 2, en donde se aprecian valores elevados fluctuando desde 0.86 hasta 0.98 (Pearson, p < 0.01) entre Lap, An, Al y Pe. Así mismo, las correlaciones entre las variables anteriores y las referentes a la valva izquierda (La, Ad, Arv) de S. cruenta fueron menores. Finalmente, existe también una alta correlación entre La y Ad (0.91).
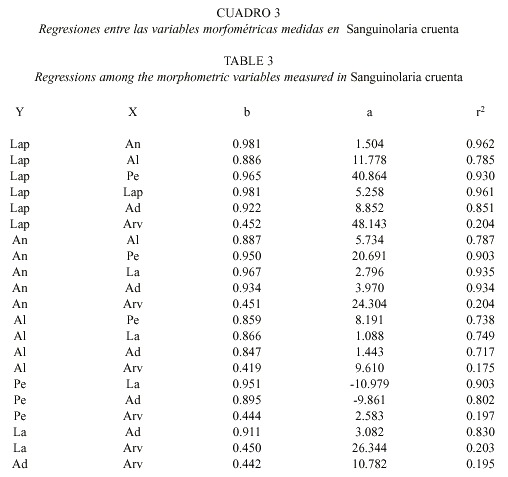
Las ecuaciones de regresión obtenidas para las relaciones entre las variables morfométricas medidas se muestran en el Cuadro 3, e indican que existe un crecimiento de tipo alométrico positivo entre las variables (t-student, p< 0.05).
Discusión
La talla máxima evaluada para S. cruenta fue de 75.55 mm, menor que la máxima reportada en el Caribe (88.90 mm) por Abbott (1974), siendo atribuible posiblemente esta diferencia al sistema de colecta, dado que se recolectaron las conchas arrastradas hacia la orilla de la playa posterior a su mortandad, con lo cual puede ser que los especímenes de mayor tamaño estuviesen enterrados profundamente en el sedimento y que no hallan muerto durante el evento. La escasa bibliografía disponible sobre dicha especie no permite hacer mayores comparaciones con relación a las tallas.
La mayor frecuencia por tallas para la población de S. cruenta en playa Tucacas, está comprendida entre 51 mm y 65 mm, estando dentro del intervalo reportado para el Caribe por Abbott (1974), Robinson y Montoya (1987), Díaz y Puyana (1994) y Rios (1994). La distribución de la población estudiada se comporta como una curva normal sin marcadas variaciones, no encontrándose organismos inferiores a los 30 mm durante la colecta. La ausencia de individuos menores de 30 mm, se puede atribuir a factores como la depredación y mayormente a la fragilidad de las conchas pequeñas, las cuales pudieron romperse fácilmente durante su arrastre hasta el supralitoral de la playa.
Debido a que esta especie posee un mayor crecimiento a lo largo del eje antero-posterior y al bajo coeficiente de variación que presenta, dicha medida constituye la mejor variable para la descripción taxonómica de S. cruenta, pudiendo ser utilizada como estimador del crecimiento, puesto que generalmente se han utilizado variables morfométricas de las conchas de los moluscos como estimadores del mismo (Salaya y Penchaszadeh 1978, Farache 1980, Donn 1990, Na Sins 1993, Cabrera et al. 1995, Solano et al. 1995, 1997).
El crecimiento de esta especie es de tipo isométrico entre la longitud anteroposterior vs el ancho y la longitud anteroposterior vs la longitud anterior, mientras que es alométrico positivo para el resto de las variables establecidas, lo cual ha sido reportado para otros bivalvos como: Mytilus edulis (Friis 1968), Mytella guyanensis (Sibaja y Villalobos 1986), Modiolus capax (Garza y Bückle 1989, Cabrera et al. 1995) y Pinctada mazatlanica (Solano et al. 1995, 1997).
Resumen
Se colectaron 312 conchas de S. cruenta con su ligamento en Playa Tucacas, Edo. Falcón, en mayo de 1999. A cada ejemplar se le midió: longitud antero-posterior, altura total, ancho, longitud anterior, altura dorsal, área de la valva y su peso. Las variables biométricas más apropiadas para la caracterización de la especie fueron la longitud antero-posterior, la altura total, el ancho, la altura dorsal y la longitud anterior, dada su poca variabilidad. La clase de tallas más frecuente se encontró entre 41-70 mm (91% de la población). Las relaciones longitud anteroposterior-ancho y longitud anteroposterior-longitud anterior son tipo isométrico (Lap = 0.98078 An + 1.50456 y Lap = 0.98052 La + 5.25840, respectivamente), mientras las restantes son del tipo alométrico positivo.
Referencias [ Links ]
Díaz, J.M. & M. Puyana. 1994. Moluscos del Caribe colombiano. Colciencia y Fundación Natura Colombia, Colombia. 291 + LXXIV p. [ Links ]
Donn, T.E. Jr. 1990. Morphometrics of Donax serra Röding (Bivalvia: Donacidae) populations with contrasting zonation patterns. J. Coast. Res. 6: 893-901. [ Links ]Cabrera, J., R.A. Cruz, Y. Solano & M. Protti. 1995. Biometría de Modiolus capax (Bivalvia: Mytilidae) en Playa Ocotal, Guanacaste, Costa Rica. Rev. Biol. Trop. 43: 173-176. [ Links ]
Farache, V. 1980. Factores ambientales que se relacionan con la densidad relativa, la variación morfométrica y los patrones de coloración de dos especies de chipichipi, Donax denticulatus y D. striatus (Bivalvia: Donacidae). Tesis de Licenciatura, Universidad Simón Bolívar, Caracas, Venezuela. [ Links ]
Friis, B. 1968. Growth and mortality of culture mussels in the Danish Wadden Sea. Mzddelelser fra Doumarks Fiskeriog Havundersogelser 6: 47-78. [ Links ]Garza, M.C. & L.F. Bückle. 1989. Estructura de tallas, reclutamiento y crecimiento del mejillón Modiolus capax (Conrad) (Bivalvia-Mytilidae) en La Bahía de Los Angeles, Baja California, México. An. Inst. Cien. Mar Limnol. Univ. Nac. Autón. México 16: 245-254. [ Links ]
Herrera, A.J. 1995. Variación espacio-temporal de la macroinfauna bentónica asociada a playa Tucacas, Estado Falcón. Tesis de Licenciatura, Universidad Simón Bolívar, Caracas, Venezuela. [ Links ]Lodeiros, C., B. Marin & A. Prieto. 1999. Catálogo de moluscos marinos de las costas nororientales de Venezuela: Clase Bivalvia. APUDONS, Venezuela. 109 p + 9 láminas. [ Links ]
Pérez Nieto, H. 1965. Lista preliminar de los moluscos marinos del Plioceno de las capas de Cumaná, cerros de Caigüire Cumaná, Venezuela. Lagena 7: 11-21. [ Links ]Na Sins. 1993. Age and growth of the black-lip pearl oyster, Pinctada margaritifera (L) (Bivalvia, Pteriidae). J. Shellfish Res. 12: 223-228. [ Links ]
Rios, E. 1994. Seashells of Brazil. R.S. da Furg, Rio Grande, Brazil. 492 p. [ Links ]
Robinson, D.G. & M. Montoya. 1987. Los moluscos marinos de la costa Atlántica de Costa Rica. Rev. Biol. Trop. 35: 375-400. [ Links ]
Salaya, J.J. & P. Penchaszadeh. 1978. Pesqueria de la vieira, Pecten papyraceus (Mollusca-Bivalvia), en Venezuela. Proc. Gulf Carib. Fish. Inst. 31: 105- 126. [ Links ]
Sibaja, W. & C. Villalobos. 1986. Crecimiento del mejillón Mytella strigata Hanley (Bivalvia: Mytilidae), de la playa Lepanto, Puntarenas, Costa Rica. Rev. Biol. Trop. 34: 231-236. [ Links ]
Sokal, R. & J. Rohlf. 1979. Biometría. Blume, Barcelona, España. 922 p. [ Links ]
Solano, Y., J. Cabrera, M. Protti & R. Cruz. 1995. Relaciones morfométricas de Pinctada mazatlanica (Bivalvia: Pteriidae) en Puntarenas, Costa Rica. Rev. Biol. Trop. 43: 177-180. [ Links ]
Solano, Y., J. Cabrera, R. Cruz & J. Palacios. 1997. Estructura de la población y crecimiento de Pinctada mazatlanica (Pterioida: Pteriidae), Golfo de Nicoya, Costa Rica. Rev. Biol. Trop. 45: 1055-1060. [ Links ]Stanley, S.M. 1970. Relation of shell form to life habits of the Bivalvia (Mollusca). Geol. Soc. Amer., Mem. 125: 1-296. [ Links ]
Tello, J. 1975. Catálogo de la fauna venezolana. VIII Mollusca. Arte, Caracas. 599 p. [ Links ]

