










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.49 n.2 San José Jun. 2001
Recibido 23-V-2000. Corregido 25-X-2000. Aceptado 31-X-2000.
Abstract
Interannual ichthyoplankton variation, was analyzed in Bahía de la Ascensión, Mexico, during December of four consecutive years (1994-1997). A total of 32 families, 35 genera and 21 species of fish larvae were identified. The most abundant fish larvae were the Gobiidae followed by the Callionymidae, Clupeidae and Tetraodontidae. Larval diversity was low when compared with other periods (dry and rainy). Three spatial associations (internal, medium and external) were found in December 1994 and 1995. In 1996-1997, stations of the inner and outer parts of the bay were mixed. The dominant families characterized most of the faunal associations. Egg density was highest in the external zone of the bay, whereas larvae were most abundant in the inner area. Major factors affecting the fish larval assemblages during December (1994-1997) in Bahía de la Ascensión seem to be related to the nursery areas location and to the tropical fishes reproductive period.
Key words
Ichthyoplankton, distribution, spatial associations, Bahía de la Ascensión, Caribbean Sea.
El estudio sobre la biología de los peces no puede estar completo si se desconoce la historia natural de los estadios tempranos de vida. Existe un gran vacío en el conocimiento de las larvas de peces costeras tropicales; una de las principales causas de ello, es su complejidad taxonómica. El problema anterior se intensifica en los ambientes costeros con alta diversidad (Leis y Thrnski 1989, Leis 1991, Neira et al. 1998). La importancia de los estuarios como áreas de crianza para peces está bien documentada (Cain y Dean 1976, Weinstein 1979, Potter et al. 1986) sin embargo, la mayoría de esos estudios se refieren a juveniles sin atender a las etapas larvarias y abarcan periodos relativamente cortos (un año). Sólo algunos estudios analizan las fluctuaciones estacionales de las asociaciones de larvas de peces para periodos de tiempos mayores (Able 1978, Potter et al. 1986, Allen y Barker 1990, Drake y Arias 1991). Para el área costera del Caribe mexicano se han realizado muestreos de plancton desde 1990 que han generado información sobre el zooplancton (Suárez-Morales y Gasca 1994, Suárez-Morales y Gasca 1996) y el ictioplancton (Vásquez-Yeomans 1990, Vásquez-Yeomans et al. 1993, Vásquez-Yeomans et al. 1998) de esta zona. Se enfatiza la parte central del litoral de Quintana Roo, donde se localiza la Bahía de la Ascensión (Fig. 1). Esta bahía forma parte de la Reserva de la Biosfera de Sian Kaan y ha sido descrita como área de crianza para peces costeros tropicales (Espejel-Montes 1983, Vásquez-Yeomans y Richards 1999, Vásquez-Yeomans 2000).
Se pretende determinar la composición y estructura de las asociaciones de larvas de peces en la bahía, para los meses de diciembre de 1994 a 1997, meses que caracterizan a la época de vientos del Norte.
Materiales y Métodos
El material proviene de 55 muestras totales recolectadas en 15 estaciones de muestreo en la Bahía de la Ascensión, visitadas en los meses de diciembre de cuatro años consecutivos (1994 a 1997). Los arrastres fueron superficiales (0-1 m de profundidad), diurnos, circulares y con una duración de 10 minutos. Se utilizó una red estándar de plancton con boca circular (0.5 m de diámetro) con luz de malla de 0.33 mm. En la boca de la red se montó un flujómetro General Oceanics 2030R. Las muestras se fijaron con formalina al 4%, y posteriormente fueron separados y cuantificados la totalidad de los huevos y larvas de peces. Las larvas fueron identificadas al menor nivel taxonómico posible.
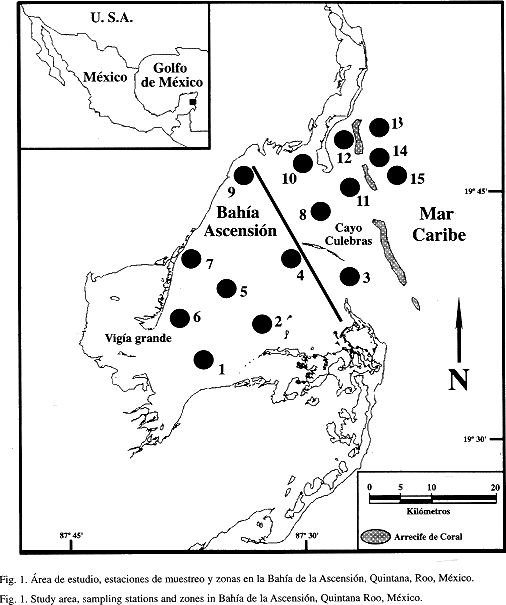
Las abundancias de huevos y larvas fueron estandarizadas como org./100 m³, y para cada estación y mes de muestreo se expresaron como el valor promedio. Debido a la dificultad que representa la identificación de larvas de peces en zonas costeras tropicales (Richards 1985, Kendall y Matarese 1994) todos los análisis ecológicos se trabajaron a nivel familia (Leis 1993). Para determinar a las familias características se utilizó el índice biológico de Sanders (1960); la diversidad larval por año fue calculada con el índice de Shannon-Wiener. Para describir las asociaciones larvales se aplicó el índice de Jaccard y el método de unión flexible (Lance y Williams 1967). Todos los análisis ecológicos se realizaron con el programa Anacom (De la Cruz-Agüero 1993). Se aplicaron análisis de varianza multifactorial (Sokal y Rohlf 1985) sobre las abundancias de huevos, larvas de peces y la diversidad larval. Los valores de abundancia fueron previamente transformados a log (x+1). Los factores de variabilidad considerados fueron el tiempo (años 1994-1997) y el espacio (zonas de la bahía: interna y externa). La zona interna fue representada por ocho estaciones (1, 2, 4, 5, 6, 7, 9 y 10), localidades someras y cercanas a las zonas de manglar, y la zona externa por siete (3, 8, 11, 12, 13, 14 y 15), estaciones con mayor influencia marina y cercanas a los arrecifes. Las larvas identificadas fueron depositadas en el Museo de Zoología de Ecosur-Chetumal (Eco-Ch-Lp).
Resultados
Se cuantificaron 766 larvas y 56 633 huevos de peces.La mayor abundancia promedio de larvas (14.2 larvas/100 m3 ± 6.9 e.e.) correspondió a 1996, mientras que la mínima (6.7 larvas/100 m3 ± 2.0 e.e.) fue registrada en 1994. Estas diferencias interanuales no resultaron significativas (Fs=0.432, p=0.7313). La distribución espacial mostró mayores abundancias larvales hacia la zona interna de la bahía (15.0 larvas/100 m3 ± 3.6 e.e.), específicamente en las estaciones cercanas a la línea de costa (1, 6, 7 y 10). La menor (5.2 larvas/100 m3 ± 1.2 e.e.) correspondió a la zona externa . El ANOVA reveló diferencias significativas en la abundancia entre zonas (Fs=6.059, p=0.0176). La interacción entre factores (años y zonas) resultó no significativa (Fs=0.481, p=0.6973).
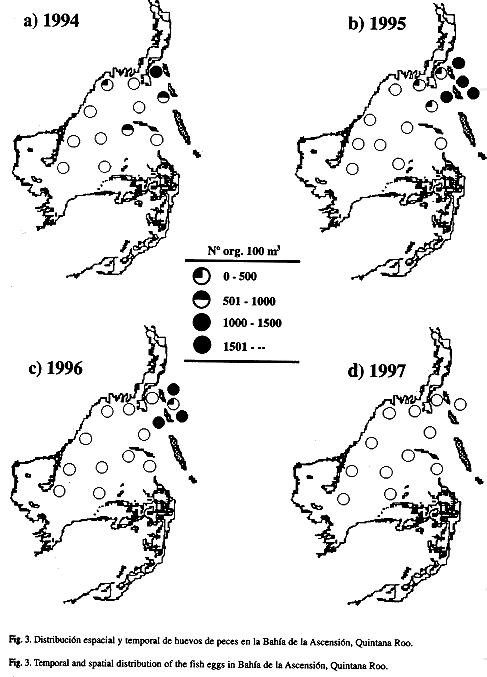
La mayor abundancia de huevos fue observada en 1995 (1 126.3 huevos/100 m3 ± 399.5 e.e.) y la menor en 1997 (90 huevos/100 m3 ± 26.2 e.e.); las variaciones interanuales en la abundancia de huevos fueron significativas (Fs=3.648, p=0.0191). Se registraron marcadas variaciones en la distribución espacial de huevos (Fs=17.126, p=0.0001).. El patrón general mostró mayores abundancias hacia la zona externa de la bahía (Fig. 3); la interacción entre factores (años y zonas de la bahía) resultó no significativa (Fs=0.339, p=0.7971).
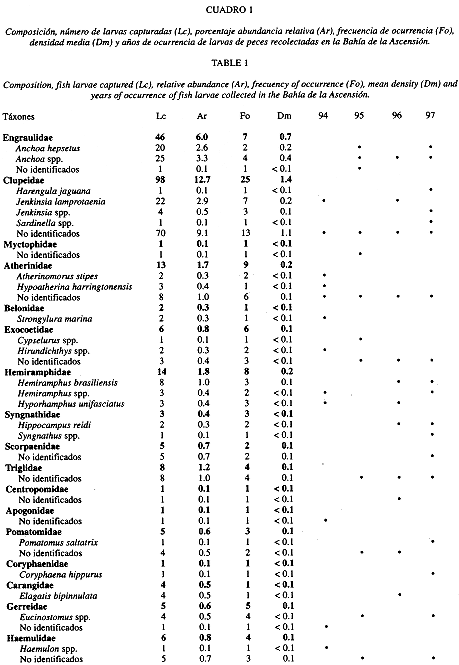
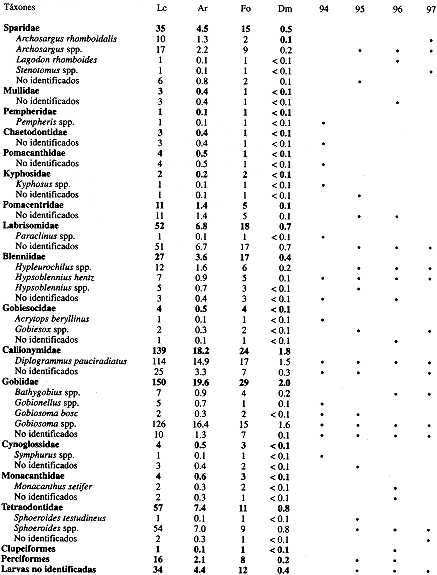
Se identificó el 93.4% de las larvas de peces recolectadas; se lograron reconocer 32 familias, 35 géneros y 21 especies (Cuadro 1). Las muestras estuvieron dominadas numéricamente por larvas de góbidos (19.6%), calionímidos (18.2%), sardinas y arenques (12.7%), tetraodóntidos (7.4%), labrisómidos (6.8%) y anchovetas (6.0%). Los táxones más abundantes fueron: Gobiosoma spp. (16.4%), Diplogrammus pauciradiatus (14.9%), Sphoeroides spp. (7.0%), labrisómidos no identificados (6.7%), Anchoa spp. (3.3%) y Jenkinsia lamprotaenia (2.9%). Un total de ocho familias (Gobiidae, Callionymidae, Clupeidae, Labrisomidae, Blenniidae, Atherinidae, Exocoetidae y Gobiesocidae) estuvieron consistentemente presentes en los cuatro años (Cuadro 1). Según el índice de dominancia de Sanders, las familias de peces que caracterizaron a los meses de diciembre del periodo 1994-1997 fueron: Callionymidae (15.5%), Clupeidae (15.0%), Gobiidae (14.5%), Engraulidae (9.1%), Tetraodontidae y Blenniidae (8.2% cada uno). La riqueza varió de 18 familias en 1994 a 20 en 1996. La diversidad larval (H) varió de 2.5 a 3.3 bits/ind correspondiendo la mínima a 1994 y la máxima a 1995. En general las diversidades observadas en la zona interna de la bahía tuvieron un comportamiento más constante en el tiempo en comparación con las registradas en la zona externa.
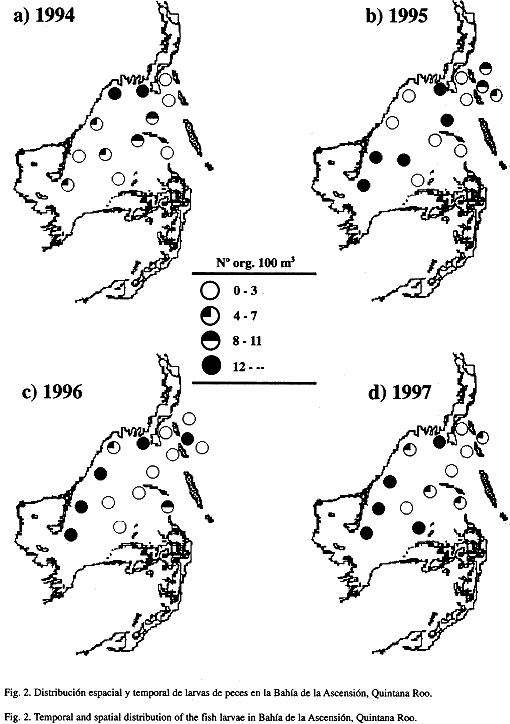
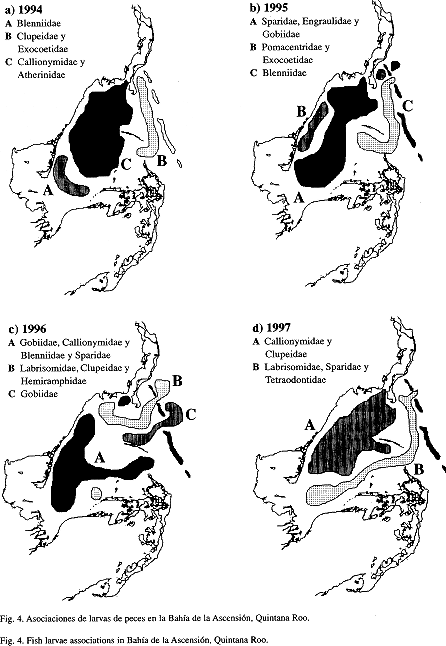
A partir de los análisis de clasificación interanuales fueron detectadas algunas variaciones. En diciembre de 1994 y 1995 se presentaron tres asociaciones de estaciones bien definidas, una que incluyó estaciones de la zona interna, otra con estaciones de la zona media y la última formada por estaciones de la zona externa. Para 1996 aunque persiste la presencia de tres asociaciones, estas se conformaron por la mezcla de estaciones de la zona interna con estaciones de la zona externa. En 1997 se observaron básicamente dos asociaciones larvales y al igual que en 1996 hubo una mezcla de estaciones (Fig. 4).
Discusión
Los resultados del presente estudio muestran características similares a las observadas en otros ecosistemas marinos y estuarinos someros (Houde y Lovdal 1984, Ditty 1986, Powell et al. 1989, Vásquez-Yeomans et al. 1998, Vásquez-Yeomans y Richards 1999, Vásquez-Yeomans 2000): densidades larvales relativamente bajas, una predominancia de larvas pequeñas, y una baja diversidad. El ictioplancton dominante encontrado fue similar a los reportados en estudios previos en los Everglades (Collins y Finucane 1984), en la bahía de Viscaino y áreas adyacentes de la Florida (Houde y Lovdal 1984, Powell et al. 1989), en el área costera de Mahahual (Vásquez-Yeomans et al. 1998) y en la misma Bahía de la Ascensión (Vásquez-Yeomans 2000). En general, el ictioplancton del presente estudio fue dominado por larvas de cinco familias de peces: góbidos, callionímidos, clupeidos, tetraodóntidos y labrisómidos que juntas alcanzaron el 64.7% del total de larvas recolectadas. En diciembre de 1990 en la misma bahía predominaron en el ictioplancton las familias Labrisomidae y Engraulidae (78.3% del total capturado)(Vásquez-Yeomans 2000), mientras que en Mahahual la comunidad ictioplánctica estuvo caracterizada por labrisómidos, góbidos y clupeidos (78.5% del total capturado)(Vásquez-Yeomans et al. 1998). En general, aunque hubo variaciones interanuales en el orden de abundancia de las familias de peces dominantes, se mantuvo una gran correspondencia entre las familias más abundantes y las familias determinadas como más importantes, según el Indice de Sanders.
En general, los valores de diversidad del presente estudio coinciden con los informados para diciembre de 1990 (Vásquez-Yeomans 2000). Contrario a lo esperado, la zona interna registró la mayor diversidad larval. Probablemente esto se debió a que no siempre fue posible muestrear todas las estaciones de la zona externa, específicamente en 1994. Comparando la diversidad obtenida en el presente estudio (época de Nortes) con la registrada en otras épocas del año (secas o lluvias) para la misma bahía, se observó que ésta fue consistentemente baja. Las mayores diversidades (53 familias y 73 especies) en la Bahía de la Ascensión se han registrado para la época de lluvias (julio y octubre) alcanzando una diversidad de hasta 3.97 bits/ind (Vásquez-Yeomans y Richards 1999).
Todas las asociaciones espaciales estuvieron caracterizadas por la presencia de las familias dominantes, lo cual es típico de ambientes costeros. El patrón de distribución larval puede relacionarse con la estrategia reproductiva de los peces costeros en áreas tropicales (Johannes 1978). Los blénidos, góbidos, tetraodóntidos y labrisómidos tienen huevos demersales por lo que sus larvas son retenidas en el interior de la bahía. Por otro lado los clupeidos, engraulidos, calionímidos y esparídos tienen huevos pelágicos y sus larvas son retenidas en las aguas someras. Johannes (1978) no explicó la estrategia seguida por las larvas de esas familias, para hacer frente a las condiciones de alta depredación de aguas someras.
Las mayores abundancias larvales registradas en la zona interna confirma lo encontrado en estudios previos en esta bahía (Vásquez-Yeomans y Richards 1999, Vásquez-Yeomans 2000), y coincide con los altos valores de biomasa zooplanctónica reportados para la zona interna a lo largo de un ciclo anual (Gasca et al. 1994). Lo anterior sugiere que en esta zona las larvas de peces encuentran una mayor disponibilidad de alimento. Además, en esta zona de manglares y con parches de pastos marinos, las larvas encuentran un mayor refugio. Para copépodos planctónicos se registró un patron de distribución espacial más homogéneo en esta bahía durante la época de vientos del Norte de 1990 a 1991 de acuerdo con Suárez-Morales y Gasca (1996). Estos autores atribuyeron que en Nortes los zooplanctontes se ven afectados por los vientos. Es probable que este efecto no sea igualmente notable en las larvas como lo es en otros organismos (Suárez-Morales y Gasca 1996). A diferencia de la abundancia larval, las mayores abundancias de huevos se registraron en estaciones de la zona externa. Esto probablemente está asociado con la proximidad de los arrecifes, ya que muchos peces arrecifales producen gran cantidad de huevos pelágicos (Johannes 1978, Sale 1980, Leis 1991) que pudieran ser transportados pasivamente hacia la boca de la bahía.
Resumen
Se analizaron las variaciones interanuales del ictioplancton, en la Bahía de la Ascensión, Quintana Roo durante diciembre de 1994 a 1997. Fueron identificadas 32 familias, 35 géneros y 21 especies de peces. Las familias dominantes fueron la Gobiidae, Callionymidae, Clupeidae y Tetraodontidae. La diversidad larval fue baja comparada con la registrada en otras épocas del año (secas y lluvias). En diciembre de 1994 y 1995 se observaron tres asociaciones espaciales (interna, media y externa). En diciembre de 1996 y 1997 las asociaciones se conformaron por la mezcla de estaciones de la parte interna y externa de la bahía. Las familias dominantes caracterizaron a la mayoría de las asociaciones faunisticas. La densidad de huevos de peces fue más alta en la zona externa de la bahía, mientras que las larvas fueron más abundantes en la zona interna. Los principales factores que afectan a las asociaciones de larvas de peces durante diciembre (1994-1997) en la bahía de la Ascensión parecen estar relacionados a la localización de áreas de crianza, zonas de desove y al periodo y estrategia reproductiva de peces estuarinos y arrecifales.
Referencias
Able, K. W. 1978. Ichthyoplankton of the St. Lawrence Estuar: composition, distribution and abundance. J. Fish. Res. Bd. Can. 35: 1518-1531. [ Links ]
Allen D. M. & D. L. Barker. 1990. Interannual variations in larval fish recruitment to estuarine epibenthic habitats. Mar. Ecol. Prog. Ser. 63: 113-125 [ Links ]
Cain, R. L. & J. M. Dean. 1976. Annual occurrence, abundance, and diversity of fish in a South Carolina intertidal creek. Mar. Biol. 36: 369-379. [ Links ]
Collins, L. A. & J. H. Finucane. 1984. Ichthyoplankton survey of the estuarine and inshore waters of the Florida Everglades, May 1971 to February 1972. U.S. Dep. Commer., NOAA Technical Report. NMFS-6, U.S. 75 p. [ Links ]
De la Cruz-Agüero, G. 1993. Anacom. Sistema para el análisis de comunidades. Versión 3.0. Manual de usuario. Depto. de Recursos del Mar, Cinvestav-Ipn. Unidad-Mérida. Yucatan, México. 99 p. [ Links ]
Ditty, J. G. 1986. Ichthyoplankton in neritic waters of the northern Gulf of Mexico off Louisiana: composition, relative abundance, and seasonality. Fish. Bull. U.S. 84: 935-946. [ Links ]
Drake, P. & A. M. Arias. 1991. Composition and seasonal fluctuations of the ichthyoplankton community in a shallow tidal channel of Cadiz Bay. J. Fish. Biol. 39: 245-263. [ Links ]
Espejel-Montes, J., L. 1983. Biología acuática, descripción general de los recursos bióticos y económicos. p. 195-215. In Centro de investigaciones de Quintana Roo (ed.). Sian Kaan, Estudios preliminares de una zona de Quintana Roo propuesta como reserva de la Biosfera. Centro de Inv. de Q. Roo. Puerto Morelos, México. [ Links ]
Gasca, R., E. Suárez-Morales & L. Vásquez-Yeomans. 1994. Estudio comparativo del zooplancton (biomasa y composición) en dos bahías del Mar Caribe mexicano. Rev. Biol. Trop. 42 (3): 595-604. [ Links ]
Houde, E. D. & J. D. A. Lovdal. 1984. Seasonality of ocurrence foods and food preferences of ichthyoplankton in Biscayne Bay, Florida. Estuar. Coast. Shelf Sci. 18: 403-419 [ Links ]
Johannes, R. E. 1978. Reproductive strategies of coastal marine fishes in the tropics. Env. Biol. Fish. 3 (1): 65-84. [ Links ]
Kendall, A. W. Jr. & A. C. Matarese. 1994. Status of early life history descriptions of marine teleosts. Fish. Bull. 92: 725-736. [ Links ]
Lance, G. N. & N. T. Williams. 1967. A general theory of classificatory sorting strategies. 1. Hiererchical systems. Computer J. 9: 373-380. [ Links ]
Leis, J. M. 1991. The pelagic stage of reef fishes: the larval biology of coral reef fishes. pp. 183-230. In P. F. Sale (ed.). The Ecology of Fishes on Coral Reefs. University of Hawaii. Honolulu [ Links ]
Leis, J. M. 1993. Larval fish assemblages near Indo-Pacific coral reefs. Bull. Mar. Sci. 53 : 362-392. [ Links ]
Leis, J. M. & T. Trnski. 1989. The larvae of Indo-Pacific shorefishes. University of Hawaii. 371 p. [ Links ]
Neira, F. J., A. G. Miskiewicz & T. Trnski. 1998. Larvae of temperate Australian fishes. Laboratory guide for larval fish identification. University of Western Australia. 474 p. [ Links ]
Potter, I. C., P. N. Claridge & R. M. Warwick. 1986. Consistency of seasonal changes in an estuarine fish assemblage. Mar. Ecol. Prog. Ser. 32: 43-58. [ Links ]
Powell, A. B., D. E. Hoss, W.F. Hettler, D. S. Peters & S. Wagner. 1989. Abundance and distribution of ichthyoplankton in Florida Bay and adjacent waters. Bull. Mar. Sci. 44: 35-48. [ Links ]
Richards, W. J. 1985. Status of the identification of the early life stages of fishes. Bull. Mar. Sci. 37: 756-760. [ Links ]
Sale, P. F. 1980. The ecology of fishes on coral reefs. Oceanogr. Mar. Biol. 18: 367-421. [ Links ]
Sanders, H. L. 1960. Benthic studies in Buzzards Bay III. The structure of the soft-bottom community. Limnol. Oceanogr. 5: 138-153. [ Links ]
Sokal, R. R. & F. G. Rohlf. 1995. Biometry. W. H. Freeman, Nueva York, 859 p. [ Links ]
Suárez-Morales, E. & R. Gasca. 1994. Zooplankton biomass fluctuations in a Mexican Caribbean bay (Bahía de la Ascensión) during a year cycle. Car. J. Sci. 30: 116-123. [ Links ]
Suárez-Morales, E & R. Gasca. 1996.Planktonic copepods of Bahía de la Ascensión, Caribbean coast of Mexico: A estacional survey. Crustacea. 69: 2 [ Links ]
Vásquez, L. 1990. Larvas de peces de Bahía de la Ascensión, Quintana Roo, p. 321-330. In D. Navarro & J.G. Robinson (eds.). Diversidad biológica en la Reserva de la Biosfera de Sian Kaan, Quintana Roo, México. Centro de Inv. de Q. Roo-PSTC/Univ. of Florida. Chetumal, México [ Links ]
Vásquez-Yeomans, L. & W. J. Richards. 1999. Variación estacional del ictioplancton de la Bahía de la Ascensión, Reserva de la Biosfera de Sian Kaan (1990-1994). Rev. Biol. Trop. 47: 201-212. [ Links ]
Vásquez, L., M. A. González, R. A. Gasca; E. Suárez, R. M. Hernández e I. Castellanos. 1993. El ictioplancton costero del Caribe mexicano: Análisis comparativo de dos bahías. Centro de Inv. de Q. Roo/Conacyt,Chetumal, México. 189 p. [ Links ]
Vásquez-Yeomans, L., U. Ordóñez-López & E. Sosa-Cordero. 1998. Fish larvae adjacent to a coral reef in the western Caribbean Sea off Mahahual, Mexico. Bull. Mar. Sci. 62: 229-245. [ Links ]
Vásquez-Yeomans, L. 2000. Seasonal variation of ichthyoplankton in a western Caribbean bay system. Env. Biol. Fishes. 58: 349-392. [ Links ]
Weinsten, M. P. 1979. Shallow marsh habitats as primary nurseries for fishes and shellfish, Cape Fear River, North Carolina. Fish. Bull., U. S. 77: 339-357. [ Links ]
1Laboratorio de Plancton, Departamento de Ecología y Sistemática Acuática.
El Colegio de la Frontera Sur. A.P. 424, C.P. 77000, Chetumal, Quintana Roo, México. Tel. (983) 216-66. FAX: (983) 204-47; cesar@ecosur-qroo.mx.

