










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.49 n.1 San José Mar. 2001
Mario Ortaz 1
Recibido 7-XII-1999. Corregido 21-VI-2000. Aceptado 31-VII-2000
Abstract
The diets of four diurnal fish species (Creagrutus bolivari, Knodus deuterodonoides, Knodus sp. and Poecilia reticulata) were examined during a year in the Orituco stream at northern Venezuela. The fishes were sampled monthly from February 1991 to March 1992 (except October 1991 and February 1992) in the stream main channel with a beach seine and a cast net. Diet is reported as frequency of ocurrence and numeric proportion because variation in prey sizes was small. Non-parametric statistical tests were applied. A total of 18 distinct prey items were found in stomachs. The diet of these fishes consisted of aquatic insects (Coleoptera, Diptera, Hemiptera, Odonata, Plecoptera and Trichoptera), allochthonous plant matter (fragments of leaves and seeds), microalgae (Chlorophyta and Bacillariophyceae) and terrestrial arthropods (Coleoptera, Diptera, Hymenoptera and Arachnida). The low percentage of empty guts and the high fullness percentage of guts suggest that food was always abundant. Aquatic insects were more important in the dry season (November - April) while plant matter and terrestrial arthropods increased in the wet season (May - October). The Proportional Similarity Measure (PS) was high between characid species and low between characids and P. reticulata in the dry season. PS decreased during the wet season because of a reduction in aquatic insect consumption. The seasonal diet shift indicated the greater importance of allochthonous food only in the wet season.
Key words: Tropical fishes; Characidae; Poecilidae; Creagrutus bolivari; Poecilia reticulata; Knodus deuterodonoides; Stomach content analysis; Orituco stream; Venezuela.
It is generally believed that fishes in shaded headwater tropical streams rarely use their feeding specializations and that they depend primarily on allochthonous food such as terrestrial arthropods, terrestrial plant debris and other food that drops into the water (Lowe-McConnell 1987). However, some studies have indicated that fishes inhabiting these environment may depend on autochthonous items such as periphyton, aquatic insects and fishes (Zaret and Rand 1971, Moyle and Senanayake 1984, Olurin et al 1991). Angermeier and Karr (1983) studied the relation between fish community structure and food availability in Panamanian streams; they classified fishes in seven feeding guilds (algivores, aquatic insectivores, general insectivores, piscivores, scale-eaters, terrestrial herbivores and omnivores). Furthermore, they found that the species richness of feeding guilds increased with stream size and canopy openness.
According to Welcomme (1985), fishes appear to be highly facultative in their feeding in tropical rivers and stream. Evidence from a variety of systems (Knöppel 1970, Mago-Leccia 1970, Bishop 1973, Lowe-McConnell 1975, 1987, Saul 1975, Silva et al. 1980, Goulding 1980, Prejs and Prejs 1987, Olurin et al. 1991) indicate that the same food resource may be shared by numerous species and that each specie may successively exploit several different resources during the year. However, other reports from small streams indicate the opposite (Zaret and Rand 1971, Silva and Kortmulder 1977, Moyle and Senanayake 1984, Winemiller 1989). There are few studies about the diet of fishes in small headwater streams in Venezuela, and the present study aims at providing information on the diet seasonality and overlap among some fishes inhabiting a shaded headwater stream in northern Venezuela.
Materials and methods
Study area: the Orituco stream (66º 15 - 66º 45 W; 10º 00 - 10º 15 N), has a drainage area of 8 907 km2 and a total length of 348 km. The origin of Orituco stream is in Guatopo National Park at 1 200 masl (Rangel 1973). In this zone, the stream is 4th order (Whitton 1975) and it is bordered by a mostly evergreen gallery forest. The mean temperature and conductivity are 23.8 oC and 158 ![]() s/cm, respectively. Discharge range was from 0.39 m3/s during the dry season (November to April) to 1.56 m3/s during the wet season (May to October). Stream substrate consisted of boulders, pebbles and cobbles in riffles, and fine substrate (sand and mud) in pool habitats. The mean width and depth were 3 m and 0.5 m, respectively.
s/cm, respectively. Discharge range was from 0.39 m3/s during the dry season (November to April) to 1.56 m3/s during the wet season (May to October). Stream substrate consisted of boulders, pebbles and cobbles in riffles, and fine substrate (sand and mud) in pool habitats. The mean width and depth were 3 m and 0.5 m, respectively.
The sampling site was a 4 km long stretch of the Orituco stream within Guatopo National Park. This area has a relative seasonality with a dry season between November to April, and a wet season between May and October.
Methods: the fishes were sampled monthly between February 1991 and March 1992 except for October 1991 and February 1992. Fishes were collected using a beach seine (5 m length, 10 mm mesh) and a cast net (3.3 m diameter, 10 mm mesh) from the main channel. Sampling was done between 9: 00 and 15: 00 hours. The fishes were inmediately placed on ice after capture and they were subsequently put in 10% formalin. Fishes were measured to the nearest 0.01 mm of standard length (SL) and weighed (wet weight) to the nearest 0.01g.
The stomach (in characid species) or guts (in P. reticulata) were dissected and the contents were examined using a binocular dissecting microscope. Stomach or gut fullness percentage was determined using the following arbitrary classification: 1 = 0-24 %, 2 = 25-49 %, 3 = 50-74 %, and 4 = 75-100 %.
Food items were sorted, identified to the lowest taxonomic level posible using information provied by Merrit and Cummins (1984) and counted. Results of diet analysis were reported as numeric proportion (% n) and ocurrence frequency (% f) because variation in prey sizes was small (Hyslop 1980). The ocurrence frequency was estimated for discrete material (arthropods) and unidentificable material (vegetal debris). Estimations of the Proportional Similarity Measure (PS) were used to evaluate seasonal interspecific diet overlap (Zaret and Smith 1984). Intraspecific seasonal changes in diet were evaluated using the non-parametric Spearman rank correlation coefficient (rs) and a Kolmogorov-Smirnov two sample test (D). The diets among species were compared by a chi-square analysis.
Results
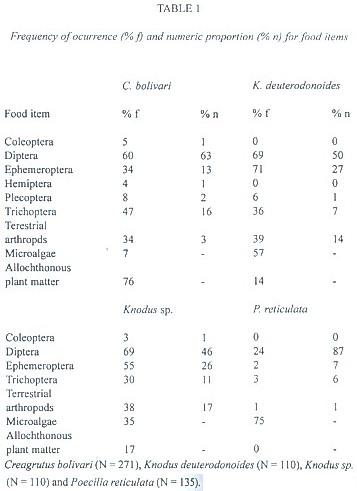
A total of 271 specimens of Creagrutus bolivari, 110 specimens of Knodus deuterodonoides, 110 specimens of Knodus sp. (a specie under taxonomic investigation) and 135 specimens of Poecilia reticulata were analyzed. Size (SL) of captured specimens was 21 - 69 mm for C. bolivari, 23 - 53 mm for K. deuterodonoides, 22 - 47 mm for Knodus sp. and 7 - 32 mm for P. reticulata.
A total of 18 distinct prey items were found, 18 for Knodus sp. 17 for C. bolivari, 14 for K. deuterodonoides and 7 for P. reticulata. In general, the diet of the four species consisted of aquatic insects, allochthonous plant matter, microalgae and terrestrial arthropods. The three Characidae were principally invertivorous (C. bolivari, K. deuterodonoides and Knodus sp.), preying mainly on Diptera, Ephemeroptera, Trichoptera and terrestrial arthropods while P. reticulata consumed mainly microalgae.
Creagrutus bolivari: the percentage of stomach fullness was high along sampling period. The diet was diversified. General values of ocurrence and numeric proportion showed a similar trend. By numeric proportion, Diptera comprised 63 %, with Trichoptera and Ephemeroptera as other important items in stomach contents (Table 1). Although ocurrence frequency of terrestrial arthropods was high, numeric proportion was not. Ocurrence values were influenced by animal fragments (heads, wings, legs and other), in which case numeric proportion could not be determined accurately. Creagrutus bolivari has the lowest microalgae frequency in relation to others species, but the highest frequency of allochthonous plant matter. Monthly numeric proportions showed that Diptera, Ephemeroptera and Trichoptera were the more important aquatic insects in C. bolivari diet. The numeric proportion of Diptera was different between the following months: March-April (D = 222.64; p <.01); April-May (D = 112.2; p <.05) and September-November (D = 61.95; p <.05). The values obtained in August and December were not representative due to low ocurrence frequency of these prey types. The numeric proportion of Trichoptera were different between April-May (D = 143.8; p <.05), and November-December (D = 36.1; p <.05). Statistical differences between September-November were not determined due to low ocurrence frequency of Trichoptera in September. The numeric proportion of Ephemeroptera was different in the following periods: April-May (D = 28.14; p <.05), November-December (D = 16.1; p <.05) and December-January (D = 36.1; p <.01). Statistical differences between August-September and September-November were not determined due to low ocurrence frequency in September. In C. bolivari, the mean number of prey was lower during the wet season (X = 7.1 in drt season versus X = 4.6 in wet season) (z = 6; p <.01). In contrast, in wet season the ocurrence frequency of allochthonous plant matter was high.
Knodus deuterodonoides: as in C. bolivari, global values of ocurrence frequency and numeric proportion showed similar trends. The percentage of full stomach was high along the sampling period. The order of numeric proportions obtained was: Diptera > Ephemeroptera > terrestrial arthropods > Trichoptera (Table 1). In K. deuterodonoides, the ocurrence frequency of terrestrial arthropods was the highest. The numeric proportion of aquatic prey types was different between seasons (rs = 0.7; p <.05). In the wet season, numeric proportion of Diptera was low (D = 6.1; p <.05) and Ephemeroptera consumption was high (D = 7.1; p <.05). The increase observed in numeric proportion of terrestrial arthropods in the wet season was not statistically significant. In the wet season, the mean number of prey types was low (X = 7.1 in dry season versus X = 3.3 in wet season) (z = 3.33; p <.01). In contrast, in the wet season the ocurrence frequency of allochthonous plant matter was high.
Knodus sp.: the percentage of full stomachs was high in both seasons. The order of numeric proportion of prey types was: Diptera > Ephemeroptera > terrestrial arthropods > Trichoptera (Table 1). As in K. deuterodonoides, numeric proportion of terrestrial arthropods was high. The diet of Knodus sp. was different between seasons (rs = 0.8; p <.05). In this case, the proportion of Diptera was lower in the wet season (D = 11; p <.05), whereas that of terrestrial arthropods was high (D = 7; p <.05). In addition, in the wet season ocurrence frequency of aquatic insects decreased while allochthonous plant matter increased. In the wet season the mean number of prey types was low (X = 6 in dry season versus X = 3.2 in wet season) (z = 4.46; p <.01).
Poecilia reticulata: the percentage of gut fullness was high in both seasons. The ocurrence frequency of algae was high (75 %) whereas aquatic insects (principally Diptera preys) made up 24 %. The numeric proportion of aquatic insects consumed were: Diptera >> Ephemeroptera > Trichoptera (Table 1). In P. reticulata, predation on Diptera was high. No interseasonal differences in numeric proportion of aquatic insects was detected. In P. reticulata, the mean number of prey items was similar in both season (X = 3.2 in dry season and X = 2 in wet season).
Species Comparisons: the chi-square analysis detected significant differences in diet between species (X2 exp = 132.41; X2 cri (df12,p<.05) = 21.03). In the dry season PS values were high. The greatest diet overlap ocurred between K. deuterodonoides and Knodus sp. while the lowest was between Knodus sp. and P. reticulata. In the dry season, half of PS values were lower than 0.67. In the wet season, PS values decreased in all comparisons and all but one value were lower 0.67. As in the dry season, the highest diet overlap was between K. deuterodonoides and Knodus sp. whereas the lowest was between K. deuterodonoides and P. reticulata. The diet overlap was moderate among characid species and low between some of them and P. reticulata (Table 2). According to the PS index, the decrease of diet overlap between characid fishes in wet season fluctuated from 7 % (C. bolivari - Knodus sp.) to 34 % (C. bolivari - K. deuterodonoides).

Discussion
The low percentage of empty stomachs (or guts in P. reticulata) and the high percentage of stomach fullness (or guts in P. reticulata), suggest that food was abundant. Similar results have been reported in other tropical streams where climatic fluctuations are smaller (Moyle and Senanayake 1984, Ortaz 1992, Segnini and Bastardo 1995). However, in tropical fishes inhabiting streams or rivers with considerable climatic fluctuations, the percentage of full stomachs are different during the year because of seasonal variation in food abundance (Zaret and Rand 1971, Winemiller 1989, Machado-Allison 1990).
Poecilia reticulata has been reported in other environment as an important insectivorous fish (Machado-Allison 1987, Ortaz 1992). However, it showed preference for other food such as microalgae in the Orituco stream, a common food of other poeciliid species (e.g Poecilia sphenops, Zaret and Rand 1971).
If criteria of Moyle and Senanayake (1984) are applied (PS > 0.67 indicate a high diet overlap), half of the PS values calculated for the dry season are high. In this season, the high PS values were associated with an increase in predation on aquatic insects. In other reports of dry season increase in diet overlap the situation is different due to a decrease in feeding rates or a switch to detritivory (e.g Lowe-McConnell 1964, Prejs and Prejs 1987). In Orituco stream there are frequent low intensity rains in some months of the dry season (December to February), caused by cold currents from intermediate latitudes (PDVSA 1993). This climatic condition and the forest phenology may promote an adequate availability of aquatic insects in this season as had been reported for other similar environments (Moyle and Senanayake 1984). The reduction of diet overlap (PS values) in the wet season may be a consequence of a general reduction in predation on aquatic insects. Among the factors responsible are: 1) a reduction of abundance of aquatic insects because of an increase in hydraulic discharge (Flecker and Feifarek 1994). 2) a decrease in prey detection because of water turbidity (Lazzaro 1987). 3) an increase in catastrophic drift caused by high discharge (Bhatnagar and Karamchandani 1970).
My observations suggest that the low predation on aquatic insects in the wet season is compensated by an increase in consumption of allochthonous matter. This is in agreement with arguments of Welcomme (1985) about the temporal diet plasticity of tropical fishes. However, in contrast to other reports that indicate the importance of allochthonous food throughout the year, the results showed that allochthonous foods are important only in the wet season. As an implication, alteration of terrestrial environment can have a negative effect on food resource for fishes.
Acknowledgments
Thanks to Cesar Diaz for field assistance. The logistical assistance of Francisco Perez and his wife are much appreciated. Thanks to the Instituto Nacional de Parques (Inparques) for logistical assistance; and Diego Rodriguez, Olga Oliveros and Maria E. Rocha for criticism on the manuscript and valuable suggestions. This work was supported by a Consejo de Desarrollo Científico y Humanístico (Universidad Central de Venezuela) grant Nº 03-10-2712/92.
Resumen
Se analizó la dieta de cuatro especies de peces (Creagrutus bolivari, Knodus deuterodonoides, Knodus sp. y Poecilia reticulata) que habitan el río Orituco al norte de Venezuela. Los muestreos se realizaron mensualmente entre febrero de 1991 y marzo de 1992 (excepto octubre/91 y febrero/92). Los peces se recolectaron con chinchorro y atarraya en el canal principal del río. La dieta se expresó como frecuencia numérica y de ocurrencia y se analizó con pruebas estadísticas no paramétricas. Se encontraron 18 items en la dieta y esta consistió de insectos acuáticos (Coleoptera, Diptera, Ephemeroptera, Hemiptera, Odonata, Plecoptera y Trichoptera), material vegetal alóctono (fragmentos de hojas y semillas), microalgas (Chlorophyta y Bacillariophyceae) y artrópodos terrestres (Coleoptera, Diptera, Hymenoptera y Arachnida). El bajo porcentaje de estómagos vacíos y el alto grado de llenura estomacal sugirieron que el alimento siempre fue abundante. En la época de sequía (noviembre - abril), los insectos acuáticos fueron los más consumidos mientras que en lluvias incrementó el consumo de artrópodos terrestres y de material vegetal. En la época de sequía el Indice de Similaridad Proporcional (PS) fue alto entre los carácidos y bajo entre ellos y P. reticulata. En lluvias, PS disminuyó debido a la reducción en el consumo de insectos acuáticos. El cambio estacional en la dieta de las especies indicó la mayor importancia del material alóctono sólo en lluvias.
References
Angermeier, P.L. & J.R. Karr. 1983. Fish communities along environmental gradients in a system on tropical streams. Env. Biol. Fish. 9: 117-135. [ Links ]
Bhatnagar, G.K. & B.J. Karamchandani. 1970. Food and feeding habits of Labeo fimbriatus (Bloch) in river Narbada near Hoshangabad. J. Insland. Fish. Soc. India. 2: 30-40.
Bishop, J.E. 1973. Limnology of a small Malayan river, Sumgae Gombak. Monographiae Biol. 22: 1-435. [ Links ]
Flecker, A.S. & B. Feifarek. 1994. Disturbance and the temporal variability of invertebrate assemblages in two Andean streams. Freshwater Biol. 31: 131-142. [ Links ]
Goulding, M. 1980. The fishes and the forest. Exploration Amazonian Natural History. University of California, Los Angeles, California. 280 p.
Hyslop, H.B.N. 1980. Stomach contents analysis, a review of methods and their application. J. Fish. Biol. 17: 411-429. [ Links ]
Knöppel, H.A. 1970. Food of Central Amazonian fishes. Amazoniana 2: 257-352. [ Links ]
Lazzaro, X. 1987. A review of planktivorous fishes: Their evolution, feeding behaviours, selectivities, and impacts. Hydrobiologia 146: 97-167. [ Links ]
Lowe-McConnell, R.H. 1964. The fishes of the Rupununi savanna district of British Guaina, South America. Part 1. Ecological grouping of fish species and effects of the seasonal cycle on the fish. J. Limn. Soc. 45: 103-144.
Lowe-McConnell, R.H. 1975. Fish communities in tropical freshwaters, Logman, London. 337 p. [ Links ]
Lowe-McConnell, R.H. 1987. Ecological studies in tropical fish communities. Cambridge University, London. 382 p.
Machado-Allison, A. 1987. Los peces de los llanos de Venezuela: un ensayo sobre su historia natural. Universidad Central de Venezuela, Caracas. 143 p.
Machado-Allison, A. 1990. Ecología de los peces de las áreas inundables de los llanos de Venezuela. Interciencia 15: 411-423. [ Links ]
Mago-Leccia, F. 1970. Estudios preliminares sobre la ecología de los peces de los llanos de Venezuela. Acta. Biol. Venez. 7: 71-102. [ Links ]
Merrit, R.W. & K.W. Cummins. 1984. An introduction to the aquatic insects of North America. Kendall/Hunt, Duduque, Iowa. 722 p.
Moyle, P.B. & F.R. Senanayake. 1984. Resource partitioning among fishes of rainforest stream in Sri Lanka. J. Zool. 202: 195-224. [ Links ]
Olurin, K.B. O.O. Awolesi. & Ago-Iwoye. 1991. Food of some fishes of Owa stream, south-western Nigeria. Arch. Hydrobiol. 122: 95-103. [ Links ]
Ortaz, M. 1992. Hábitos alimenticios de los peces de un río de montaña neotropical. Biotropica 24: 550-559. [ Links ]
PDVSA. 1993. Imagen atlas de Venezuela. Arte, Caracas. 271 p. [ Links ]
Prejs, A. & K. Prejs. 1987. Feeding of tropical freshwater fishes: seasonality in resource availability and resource use. Oecologia 71: 397-404. [ Links ]
Rangel, O. 1973. Determinación de la planicie de inundación, caso de la zona baja del río Orituco. Tesis de Licenciatura, Universidad Central de Venezuela, Caracas.
Saul, W.G. 1975. An ecological study of fishes at a site in upper Amazonian Ecuador. Proc. Acad. Nat. Sci. Phila. 127: 93-134. [ Links ]
Segnini, S. & H. Bastardo. 1995. Cambios ontogenéticos en la dieta de la trucha arcoiris (Oncorhynchus mykiss) en un río andino Neotropical. Biotropica 27: 495-508. [ Links ]
Silva, S.S. de. & K. Kortmulder. 1977. Some aspects of the biology of three species of Puntius (Barbus) (Pisces: Cyprinidae), endemic to Sri Lanka. Neth. J. Zool. 27: 182-194.
Silva, S.S. de. P.R.T. Cumaranatunga. & C.D. de Silva. 1980. Food, feeding ecology and morphological features associated with feeding of four co-ocurring cyprinids (Pisces: Cyprinidae). Neth. J. Zool. 30: 54-73. [ Links ]
Welcomme, R.L. 1985. River fisheries. FAO Fish. Tech. Pap, Rome. 330 p. [ Links ]
Whitton, B.A. 1975. River ecology. Blackwell, Berkeley, California. 725 p. [ Links ]
Winemiller, K.O. 1989. Ontogenetic diet shifts and resource partitioning among piscivorous fishes in the Venezuelan llanos. Env. Biol. Fish 26: 177-199. [ Links ]
Zaret, T.M. & A.J. Rand. 1971. Competition in tropical stream fish communities. Ecology 59: 507-515. [ Links ]
Zaret, T.M. & E.P. Smith. 1984. On measuring niches and not measuring them, p. 127-137. In T.M. Zaret (ed.). Evolutionary ecology of Neotropical freshwater fishes. Dr. Junk, Boston, Massachusetts.
1 Instituto de Biología Experimental, Universidad Central de Venezuela. Apartado postal 47114 Caracas 1041. Fax 58.2.7535897; capricorniomvos@starmedia.com

