










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.49 n.1 San José Mar. 2001
Recibido 16-II-2000. Corregido 3-VII-2000. Aceptado 31-VII-2000
Abstract
Growth of the octopus (Octopus maya) off Yucatan (Mexico) was estimated from a long-term study (seven years) by the length-based methods ELEFAN, PROJMAT and SLCA. Some 19 251 octopuses with a range of mantle length between 50 and 240 mm were sampled from commercial landings in 1983 - 1987, 1989 and 1992. The jackknife technique was applied to deal with uncertainty in growth estimates resulting from chance variations in sampling design. The growth index
Key words: Growth, length-based methods, jackknife, uncertainty, Octopus maya, Yucatan.
Los pulpos Octopus maya (Voss y Solís, 1966) y Octopus (cf.) vulgaris constituyen el primer recurso pesquero en términos económicos y el segundo en volúmenes de captura del Estado de Yucatán, México, situándose entre las pesquerías de octópodos con mayor potencial mundial (Solís-Ramírez 1994). En el caso particular de O. maya, se ha sugerido un estado de sobreexplotación (Solís-Ramírez 1994, Chávez 1994). A efectos de un adecuado ordenamiento pesquero, se han realizado estudios dirigidos a cuantificar el crecimiento individual (Van Heukelem, 1976, Solís-Ramírez y Chávez 1986, Arreguín-Sánchez 1992, Zambrano 1992), proceso poblacional clave en la dinámica poblacional de un recurso explotado. Las estimaciones de crecimiento son esenciales como insumos a modelos poblacionales dirigidos a evaluar estrategias de manejo.
Los estudios mencionados estuvieron basados en métodos indirectos, los cuales analizan la distribución de frecuencia de longitudes de un stock para estimar los parámetros de crecimiento. No obstante, en ninguno se realizaron estimaciones alternativas por más de un método, ni tampoco se cuantificó la incertidumbre asociada a dichas estimaciones. Esta incertidumbre cobra especial importancia en recursos tropicales, donde ciertas características del ciclo de vida (e.g., ciclo reproductivo y reclutamiento continuos) impiden una clara progresión de clases modales, lo cual resalta la importancia de determinar la influencia que ejerce cada muestra mensual (Leonce-Valencia y Defeo 1997). Es muy común además la presencia de errores de medición y sesgos en la obtención de los datos. En O. maya existe una fuente adicional de incertidumbre dada por el acceso a información de capturas comerciales solamente para los cuatro meses y medio (15 agosto a 31 de diciembre) en los cuales opera la pesquería, como resultado de una medida de manejo.
Los objetivos principales de este trabajo fueron: (1) estimar los parámetros de crecimiento de O. maya de la costa de Yucatán aplicando los métodos indirectos ELEFAN (Electronic Length Frequency Analysis: Gayanilo et al. 1994), PROJMAT (Projection Matrix Method: Shepherd 1987a) y SLCA (Shepherds Length Composition Analysis: Shepherd 1987b); (2) cuantificar la incertidumbre en los parámetros resultante de variaciones en la estructura de la base de datos; y (3) comparar nuestras estimaciones con aquellas reportadas en la literatura para O. maya, usando el índice de crecimiento ![]() (Defeo et al. 1992a).
(Defeo et al. 1992a).
Materiales y métodos
Se utilizaron muestras mensuales de O. maya provenientes de muestreos de desembarque en el puerto Celestún, Yucatán, entre agosto y diciembre de los años 1983-1987, 1989 y 1992. Los organismos fueron medidos en su longitud de manto, comprendida entre los ojos y el extremo distal del mismo, con 1 mm de precisión. Las frecuencias de tallas fueron agrupadas en un intervalo de clase de 10 mm, siguiendo criterios de Defeo et al. (1992b).
Para estimar los parámetros de crecimiento se aplicaron los métodos ELEFAN (Gayanilo et al. 1994), PROJMAT (Shepherd 1987a) y SLCA (Shepherd 1987b), los cuales suponen un crecimiento individual descrito por el modelo de von Bertalanffy (1938). En breve, cada método funciona como sigue: (1) ELEFAN identifica picos y huecos en las distribuciones de frecuencia de tallas y ajusta en forma iterativa la curva de crecimiento que pasa por el máximo número de picos, no suponiendo normalidad en dichas distribuciones. Un índice de bondad de ajuste denominado Rn es usado para evaluar la función obtenida (ver Gayanilo et al. 1994 para detalles). (2) PROJMAT está basado en la matriz de proyección de Leslie y proyecta cada muestra del tiempo t a t+1, usando valores semilla de K y L![]() (Shepherd 1987a). La muestra proyectada es evaluada estadísticamente con la observada a través de un procedimiento de minimización de suma de cuadrados de las diferencias entre ambas. Esto es repetido para las n muestras que constituyen la base de datos para un amplio rango de valores de K y L
(Shepherd 1987a). La muestra proyectada es evaluada estadísticamente con la observada a través de un procedimiento de minimización de suma de cuadrados de las diferencias entre ambas. Esto es repetido para las n muestras que constituyen la base de datos para un amplio rango de valores de K y L![]() , resultando la mejor curva aquella que minimiza la suma de cuadrados (Basson et al. 1988). (3) SLCA está basado en el ajuste de una función coseno independiente del número y posición de las potenciales modas en la muestra. El ajuste de dicha función a los datos se realiza considerando los límites superior e inferior de cada intervalo de clase, así como la fracción del año en que cada muestra fue tomada. Una función de prueba evalúa la bondad de ajuste de diferentes combinaciones de parámetros de crecimiento. Detalles de los algoritmos son aportados por Defeo et al. (1992a) y Leonce-Valencia y Defeo (1997).
, resultando la mejor curva aquella que minimiza la suma de cuadrados (Basson et al. 1988). (3) SLCA está basado en el ajuste de una función coseno independiente del número y posición de las potenciales modas en la muestra. El ajuste de dicha función a los datos se realiza considerando los límites superior e inferior de cada intervalo de clase, así como la fracción del año en que cada muestra fue tomada. Una función de prueba evalúa la bondad de ajuste de diferentes combinaciones de parámetros de crecimiento. Detalles de los algoritmos son aportados por Defeo et al. (1992a) y Leonce-Valencia y Defeo (1997).
Se aplicó la técnica de corte jackknife (Efron 1982) para evaluar la influencia de cada muestra en la estimación de los parámetros de crecimiento. Para ello se retiró una muestra (en este caso un mes) secuencialmente y con reemplazo del grupo de las n muestras disponibles en cada año, a fin de obtener n diferentes combinaciones de parámetros de crecimiento (Leonce-Valencia y Defeo 1997). El error porcentual (EP) y el coeficiente de variación porcentual (CV) fueron usados como estimadores de precisión. El primero fue obtenido como:
![]()
donde St es el estimador del parámetro de crecimiento usando la base de datos anual completa (4 meses), y Stj es el estimador promedio derivado de los n seudovalores jackknife. EP es óptimo cuando es igual a 0 (St=Stj). El CV jackknife fue obtenido como:
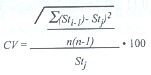
donde Sti-1 es el estimador del parámetro al omitir una muestra y n es el tamaño de la muestra anual (n = 4).
Como criterio para comparar diferentes estimaciones de crecimiento, se utilizó el índice de crecimiento estándar f = 2log10(L![]() ) + log10K (Munro y Pauly 1983, Pauly y Munro 1984), el cual provee un parámetro unificado de crecimiento y no muestra variaciones marcadas como L
) + log10K (Munro y Pauly 1983, Pauly y Munro 1984), el cual provee un parámetro unificado de crecimiento y no muestra variaciones marcadas como L![]() y K en forma individual (Defeo et al. 1992a, Leonce-Valencia y Defeo 1997). A fin de evaluar diferencias estadísticas, las estimaciones de
y K en forma individual (Defeo et al. 1992a, Leonce-Valencia y Defeo 1997). A fin de evaluar diferencias estadísticas, las estimaciones de ![]() derivadas del análisis jackknife fueron sometidas a un análisis de varianza de 2 vías, empleando los métodos y años como factores principales. En caso de detectar diferencias, se realizó un análisis de comparaciones múltiples empleando la prueba de Newman – Keuls (Zar 1996).
derivadas del análisis jackknife fueron sometidas a un análisis de varianza de 2 vías, empleando los métodos y años como factores principales. En caso de detectar diferencias, se realizó un análisis de comparaciones múltiples empleando la prueba de Newman – Keuls (Zar 1996).
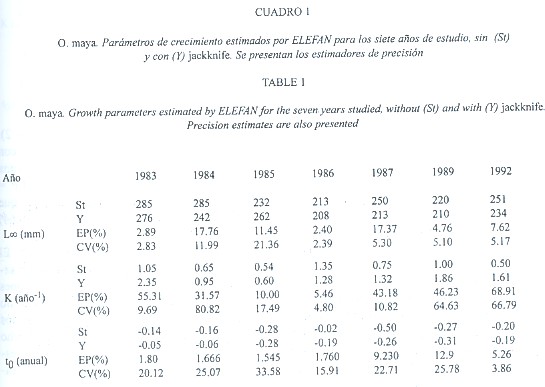
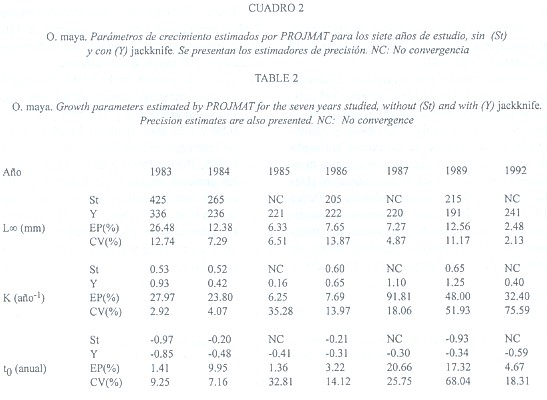
1) Toda la información: ELEFAN convergió en una estimación óptima de parámetros. L![]() varió entre 213 y 285 mm y K entre 0.50 y 1.35 año-1 (Cuadro 1). PROJMAT no convergió en 3 de los 7 años analizados (1985, 1987 y 1992) y en los años restantes presentó amplia variación en L
varió entre 213 y 285 mm y K entre 0.50 y 1.35 año-1 (Cuadro 1). PROJMAT no convergió en 3 de los 7 años analizados (1985, 1987 y 1992) y en los años restantes presentó amplia variación en L![]() (205 a 425 mm), mientras que K fluctuó de 0.52 a 0.65 año-1 (Cuadro 2). SLCA aportó estimaciones poco variables de L
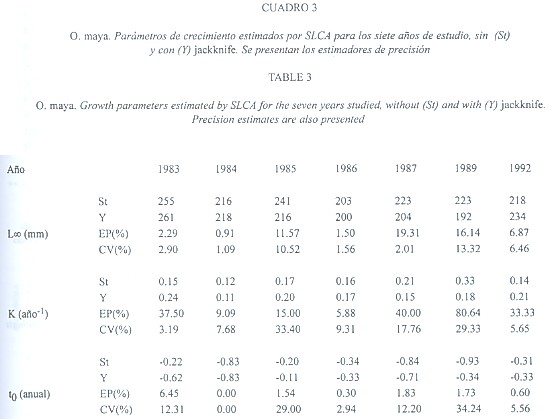
(205 a 425 mm), mientras que K fluctuó de 0.52 a 0.65 año-1 (Cuadro 2). SLCA aportó estimaciones poco variables de L![]() (203 a 255 mm) y K (0.12 a 0.33 año–1) (Cuadro 3).
(203 a 255 mm) y K (0.12 a 0.33 año–1) (Cuadro 3).
2) Análisis jackknife: La aplicación del jackknife conjuntamente con ELEFAN, PROJMAT y SLCA mostró una fuerte influencia interanual en la estimaciones. La mayor variabilidad fue observada en L![]() (191 a 336 mm) y K (0.16 a 1.25 año-1) estimados por PROJMAT (Cuadro 2), seguidas en orden decreciente por ELEFAN (L
(191 a 336 mm) y K (0.16 a 1.25 año-1) estimados por PROJMAT (Cuadro 2), seguidas en orden decreciente por ELEFAN (L![]() de 208 a 276 mm y K de 0.60 a 2.35 año-1: Cuadro 1) y SLCA (L
de 208 a 276 mm y K de 0.60 a 2.35 año-1: Cuadro 1) y SLCA (L![]() de 192 a 261 mm y K de 0.11 a 0.24 año-1: Cuadro 3).
de 192 a 261 mm y K de 0.11 a 0.24 año-1: Cuadro 3).
Los estimadores de precisión EP y CV reflejaron una importante variación intra e interanual, pero no variaron significativamente entre métodos (ANOVA 0.08<F2,18<2.61: P>0.10 en todos los casos). ELEFAN proveyó valores de L![]() con un CV entre 2.39 (año 1986) y 21.36 (1985), y un EP entre 2.89 y 17.76 en 1983 y 1984 respectivamente, mientras que K presentó un CV de 4.80 a 66.79 y EP entre 5.46 y 68.91 (Cuadro 1). PROJMAT presentó el mayor valor de EP para L
con un CV entre 2.39 (año 1986) y 21.36 (1985), y un EP entre 2.89 y 17.76 en 1983 y 1984 respectivamente, mientras que K presentó un CV de 4.80 a 66.79 y EP entre 5.46 y 68.91 (Cuadro 1). PROJMAT presentó el mayor valor de EP para L![]() en 1983 (26.48), mientras que el CV fue máximo en 1986 (13.87); los valores de K variaron hasta 91.81 (EP) en el año 1987 y 75.59 (CV) en 1992 (Cuadro 2). SLCA presentó la menor variación con respecto a los otros 2 métodos: la máxima variación de L
en 1983 (26.48), mientras que el CV fue máximo en 1986 (13.87); los valores de K variaron hasta 91.81 (EP) en el año 1987 y 75.59 (CV) en 1992 (Cuadro 2). SLCA presentó la menor variación con respecto a los otros 2 métodos: la máxima variación de L![]() se dio en 1987 (EP = 19.31) y 1989 (CV = 13.32), mientras que para K ocurrió en 1989 (EP = 80.64) (Cuadro 3).
se dio en 1987 (EP = 19.31) y 1989 (CV = 13.32), mientras que para K ocurrió en 1989 (EP = 80.64) (Cuadro 3).
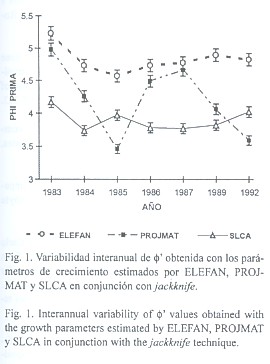
3) Análisis comparativo mediante ![]() : El análisis de
: El análisis de ![]() derivado de la omisión secuencial de muestras mostró patrones claramente definidos (Fig. 1): a)
derivado de la omisión secuencial de muestras mostró patrones claramente definidos (Fig. 1): a) ![]() no mostró la variabilidad individual de los parámetros individuales K y L
no mostró la variabilidad individual de los parámetros individuales K y L![]() , hecho reflejado en una reducida desviación estándar para cada año y método considerados. b) ELEFAN aportó valores significativamente mayores de
, hecho reflejado en una reducida desviación estándar para cada año y método considerados. b) ELEFAN aportó valores significativamente mayores de ![]() (4.57 a 5.23), seguido en orden decreciente por PROJMAT (3.46 a 4.97) y SLCA (3.74 a 4.66). c)
(4.57 a 5.23), seguido en orden decreciente por PROJMAT (3.46 a 4.97) y SLCA (3.74 a 4.66). c) ![]() varió significativamente entre métodos (F2,60 = 25.87; P<<0.0001) y entre años (ANOVA F6,60 = 4.52; P<0.001). Las comparaciones múltiples a través de la prueba de Newman-Keuls para rangos críticos identificó diferencias individuales significativas entre métodos (Cuadro 4a) y del año 1983 con respecto a los demás, presentando mayor
varió significativamente entre métodos (F2,60 = 25.87; P<<0.0001) y entre años (ANOVA F6,60 = 4.52; P<0.001). Las comparaciones múltiples a través de la prueba de Newman-Keuls para rangos críticos identificó diferencias individuales significativas entre métodos (Cuadro 4a) y del año 1983 con respecto a los demás, presentando mayor ![]() en los tres métodos empleados (Fig. 1, Cuadro 4b).
en los tres métodos empleados (Fig. 1, Cuadro 4b).
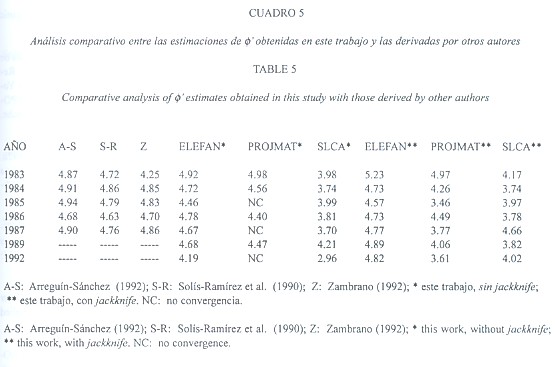
Los valores de ![]() obtenidos con ELEFAN estuvieron comprendidos en el ámbito de variación de aquellos derivados de las combinaciones de K y L
obtenidos con ELEFAN estuvieron comprendidos en el ámbito de variación de aquellos derivados de las combinaciones de K y L![]() reportadas en la literatura, mientras que los de PROJMAT y SLCA fueron menores (Cuadro 5). Esta tendencia fue válida para todos los años donde se registraron estimaciones por varias fuentes (i.e., 1983 a 1987), lo cual sugiere una tendencia sistemática de PROJMAT y SLCA a subestimar los parámetros de crecimiento (Fig. 1).
reportadas en la literatura, mientras que los de PROJMAT y SLCA fueron menores (Cuadro 5). Esta tendencia fue válida para todos los años donde se registraron estimaciones por varias fuentes (i.e., 1983 a 1987), lo cual sugiere una tendencia sistemática de PROJMAT y SLCA a subestimar los parámetros de crecimiento (Fig. 1).

Discusión
El jackknife permitió determinar una fuerte variabilidad en los parámetros individuales de crecimiento para los 3 métodos, en especial PROJMAT. SLCA presentó la menor variabilidad en las estimaciones, aunque fuera del rango observado de valores previamente reportados en la literatura. El análisis comparativo de f en conjunción con jackknife corroboró la suma utilidad de este parámetro a efectos comparativos, ya que la variabilidad dentro de años disminuyó sensiblemente (ver Fig. 1). La pequeña variación intermensual mostraría una fuerte coherencia en las estimaciones de crecimiento a la omisión secuencial de muestras. Asimismo, permitió cuantificar diferencias estadísticamente significativas entre años y especialmente entre métodos, donde se ratificó la subestimación de la tasa de crecimiento de O. maya por parte de PROJMAT y SLCA.
Las estimaciones de crecimiento de O. maya condicen además con aspectos claves de su biología, tales como su corta a mediana longevidad, estimada en cautiverio (hasta 12 meses: Van Heukelem 1976, 1977) o con base en muestreos biológicos o de la flota comercial (de 1.5 a 5 años según Solís-Ramírez y Chávez 1986, ver también Van Heukelem 1983 y Solís-Ramírez 1994). La longevidad del pulpo, estimada en este trabajo como el cociente 3/K y usando valores promedio de K derivados del análisis jackknife en el largo plazo, sería cercana a 2.5 años según estimaciones del ELEFAN, mientras que a partir de PROJMAT y SLCA sería 6.5 y 17.4 años. Estos valores están totalmente fuera del contexto biológico de la especie y reafirman su poca utilidad a efectos de estimar el crecimiento en el pulpo.
En suma, teniendo en cuenta las características biológicas del recurso, el desempeño de los 3 métodos con base en un análisis robusto de largo plazo y la coincidencia de las estimaciones de ![]() con aquellas reportadas en la literatura, se sugiere estimar el crecimiento de pulpo empleando el jackknife conjuntamente con ELEFAN. Las estimaciones de PROJMAT y SLCA generaron valores de
con aquellas reportadas en la literatura, se sugiere estimar el crecimiento de pulpo empleando el jackknife conjuntamente con ELEFAN. Las estimaciones de PROJMAT y SLCA generaron valores de ![]() y estimaciones de longevidad no acordes con la biología del recurso y por tanto no serían recomendables para estimar el crecimiento de O. maya.
y estimaciones de longevidad no acordes con la biología del recurso y por tanto no serían recomendables para estimar el crecimiento de O. maya.
Agradecimientos
Este trabajo forma parte de la tesis de Maestría de R.N.V. en CINVESTAV-IPN Unidad Mérida, la cual fue realizada con el apoyo de CONACYT. Agradecemos las sugerencias de dos revisores anónimos, así como el apoyo editorial por parte del Dr. Monge-Nájera.
Resumen
Los parámetros de crecimiento del pulpo (Octopus maya) de las costas de Yucatán (México) fueron estimados en siete años comprendidos entre 1983-1987, 1989 y 1992. A tales efectos se realizó un análisis de frecuencia de tallas de 19 251 pulpos muestreados de la captura comercial, con un intervalo de longitud de manto comprendido entre 50 y 240 mm. Se usaron los métodos indirectos ELEFAN, PROJMAT y SLCA. La incertidumbre de las estimaciones inherente a cada método y derivada del diseño de muestreo fue analizada con la técnica jackknife. Los parámetros de crecimiento L![]() y K se unificaron a efectos comparativos mediante el índice phi prima (
y K se unificaron a efectos comparativos mediante el índice phi prima (![]() ). El método ELEFAN resultó ser el más adecuado para estimar los parámetros de crecimiento de O. maya, brindando valores en el rango de aquellos previamente reportados en la literatura, mientras que PROJMAT y SLCA mostraron problemas en la convergencia de los parámetros, y tendieron a subestimarlos. Las estimaciones mediante jackknife detectaron poca variabilidad intermensual en
). El método ELEFAN resultó ser el más adecuado para estimar los parámetros de crecimiento de O. maya, brindando valores en el rango de aquellos previamente reportados en la literatura, mientras que PROJMAT y SLCA mostraron problemas en la convergencia de los parámetros, y tendieron a subestimarlos. Las estimaciones mediante jackknife detectaron poca variabilidad intermensual en ![]() pero gran variabilidad interanual. Los estimadores de precisión de la técnica jackknife, error porcentual y coeficiente de variación porcentual, no difirieron significativamente entre métodos. Las estimaciones de
pero gran variabilidad interanual. Los estimadores de precisión de la técnica jackknife, error porcentual y coeficiente de variación porcentual, no difirieron significativamente entre métodos. Las estimaciones de ![]() derivadas del ELEFAN variaron entre 4.19 y 5.23, coincidiendo con aquellas reportadas en la literatura (entre 4.25 y 4.91), mientras que PROJMAT y SLCA aportaron valores significativamente menores. Los resultados de dicho métodos condicen además con aspectos de la biología de la especie (e.g., corta longevidad). Sugerimos utilizar el ELEFAN conjuntamente con jackknife para estimar los parámetros de crecimiento de Octopus maya.
derivadas del ELEFAN variaron entre 4.19 y 5.23, coincidiendo con aquellas reportadas en la literatura (entre 4.25 y 4.91), mientras que PROJMAT y SLCA aportaron valores significativamente menores. Los resultados de dicho métodos condicen además con aspectos de la biología de la especie (e.g., corta longevidad). Sugerimos utilizar el ELEFAN conjuntamente con jackknife para estimar los parámetros de crecimiento de Octopus maya.
Referencias
Arreguín-Sánchez, F. 1992. Growth and seasonal recruitment of the octopus (Octopus maya) fishery from the Campeche Bank, caught by the artisanal fleet. NAGA, Filipinas 15: 31-34 [ Links ]
Basson, M., A.A. Rosenberg & J.R. Beddington. 1988. The accuracy and reliability of two new methods for estimating growth parameters from length frequency data. J. Cons. Int. Explor. Mer. 44: 277-285. [ Links ]
Bertalanffy, L. von. 1938. A quantitative theory of organic growth. Human Biol. 10: 181-213. [ Links ]
Chávez, E. 1994. Los recursos marinos de la Península de Yucatán, p. 1-12. In A. Yañez-Arancibia (ed.). Recursos faunísticos del litoral de la Península de Yucatán. Universidad Autónoma de Campeche, EPOMEX Serie Científica 2, Campeche, Campeche, México. [ Links ]
Defeo, O., F. Arreguín-Sánchez & J. Sánchez. 1992a. Growth study for the yellow clam Mesodesma mactroides: a comparative analysis of three length-based methods. Sci. Mar. 56: 53-59. [ Links ]
Defeo, O., E. Ortiz & J.C. Castilla. 1992b. Growth, mortality and recruitment of the yellow clam Mesodesma mactroides in Uruguayan beaches. Mar. Biol. 114: 429-437. [ Links ]
Efron, B. 1982. The Jackknife, the Bootstrap and Other Resampling Plans. Society for Industrial and Applied Mathematics, Filadelfia, Pensilvania. 92 p. [ Links ]
Gayanilo, F.C. Jr., P. Sparre & D. Pauly. 1994. The FAO-ICLARM Stock Assessment Tools (FISAT) Users Guide. FAO Comp. Inf. Ser. Fisheries 7, FAO, Roma, 126 p. [ Links ]
Leonce-Valencia, C. & O. Defeo. 1997. Evaluation of three length-based methods for estimaitng growth in tropical fishes: the red snapper Lutjanus campechanus of the Campeche Bank (Mexico). Sci. Mar. 61: 297-303. [ Links ]
Munro, J.L. & D. Pauly. 1983. A simple method for comparing the growth of fishes and invertebrates. Fishbyte 1: 5-6. [ Links ]
Pauly, D. & J.L. Munro. 1984. Once more on the comparison of growth in fish and invertebrates. Fishbyte 2: 21. [ Links ]
Rosenberg, A.A. & J.R. Beddington. 1987. Monte-Carlo testing of methods for estimating growth from length-frequency data with general conditions for their applicability, p 283-298. In D. Pauly & G.R. Morgan (ed.). Length-based methods in fisheries research. ICLARM Conf. Proc. 13, Manila. [ Links ]
Sheperd, J. G. 1987a. Towards a method for short-term forecasting of catch rates based on length compositions, p. 167-176. In D. Pauly & G.R. Morgan (ed.). Length-based methods in fisheries research. ICLARM Conf. Proc. 13, Manila. [ Links ]
Shepherd, J.G. 1987b. A weakly parametric method for estimating growth parameters from length composition data, p. 113-119. In D. Pauly & G.R. Morgan (ed.). Length-based methods in fisheries research. ICLARM Conf. Proc. 13, Manila. [ Links ]
Solís-Ramírez, M. J. 1994. Mollusca de la Península de Yucatán, México, p. 13-32. In A. Yañez-Arancibia (ed.). Recursos faunísticos del litoral de la Península de Yucatán. Universidad Autónoma de Campeche. EPOMEX Serie Científica 2, Campeche, Campeche, México. [ Links ]
Solís-Ramírez, M. J. & E.A. Chávez. 1986. Evaluación y régimen óptimo de pesca del pulpo en la Península de Yucatán. An. Inst. Cien. Mar Limnol. UNAM 13: 1-18. [ Links ]
Solís-Ramírez, M.J., M.E. Gónzalez de la Rosa& J. Sánchez-Chávez. 1990. Coeficientes de la relación peso-longitud y parámetros de crecimiento del pulpo Octopus maya (Mollusca: Cephalopoda) de la costa de Yucatán. Informe Técnico, Centro Regional de Investigación Pesquera de Yucaltepén, Yucatán. 10 p. [ Links ]
Van Heukelem, W.F. 1976. Growth, bioenergetics and life span of Octopus cyanea and Octopus maya. Tesis de Doctorado, Universidad de Hawaii. [ Links ]
Van Heukelem, W.F. 1977. Laboratory maintenance, breeding, rearing and biomedical research potential of the Yucatan octopus (Octopus maya). Lab. Anim. Sci. 27: 852-859. [ Links ]
Van Heukelem, W.F. 1983. Octopus maya, p. 311-323. In P.R. Boyle (ed.). Cephalopod life cycles, Vol. 2. Academic, Nueva York. [ Links ]
Zambrano, G.L. 1992. Contribución al conocimiento de la mortalidad natural de Octopus maya en la plataforma continental de la Península de Yucatán, México. Tesis de Licenciatura, Facultad de Ciencias UNAM, México D.F., México. [ Links ]
Zar, J.H., 1996. Biostatistical Analysis. 3a edición, Prentice-Hall, Nueva Jersey. 662 p. [ Links ]
1 Laboratorio de Biología Pesquera. CINVESTAV IPN Unidad Mérida, A. P. 73 Cordemex, 97310 Mérida, Yucatán, México; fax 5299-812334; odefeo@mda.cinvestav.mx

