










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.49 n.1 San José Mar. 2001
S.S.S. Sarma1,*, Paula Susana Larios Jurado1 & S. Nandini2
Received 6-XII-1999. Corrected 18-VII-2000. Accepted 31-VII-2000
Abstract
We compared the population growth of B. calyciflorus and B. patulus using the green alga Chlorella vulgaris, bakers yeast Saccharomyces cerevisiae or their mixture in equal proportions as food. Food was offered once every 24 h in two concentrations (low: 1x106 and high: 3x106 ind. ml-1) separately for each species. The experiments were terminated after 15 days. In general, at any food type or concentration, B. patulus reached a higher population density. A diet of Chlorella alone supported a higher population growth of both rotifer species than yeast alone. B. calyciflorus and B. patulus achieved highest population densities (103±8 ind. ml-1 and 296±20 ind. ml-1, respectively) on a diet of Chlorella at 3x106 ind. ml-1. When cultured using the mixture of Chlorella and yeast, the maximal population densities of B. calyciflorus were lower than those grown on Chlorella. Under similar conditions, the maximal abundance values of B. patulus were comparable in both food types. Regardless of food type and density the rate of population increase per day (r) for B. calyciflorus varied from 0.13±0.03 to 0.63±0.04. These values for B. patulus ranged from 0.19±0.01 to 0.37±0.01. The results indicated that even though Chlorella was a superior foof for the tested rotifers, yeast can be effectively used at low concentrations to supplement algal requirements in rotifer culture systems.
Key words: Population growth, alga, yeast, Rotifera.
Laboratory cultivation of brachionid rotifers has been successfully done using green algae. In order to supplement the algal quantity, Hirata and Mori (1967) introduced the use of bakers' yeast as food for the saline water species Brachionus plicatilis. Since then a number of investigators have used bakers' yeast as food for this species; rotifers grown in this way have also been nutritionally enriched (Fernandez-Reiriz and Labarta 1996, Lie et al. 1997). A vast majority of researchers used yeast only for B. plicatilis and Brachionus rotundiformis. In aquaculture, in addition to these two species, several other rotifer taxa such as B. calyciflorus, B. rubens and B. patulus are used as starter food (Rottmann et al. 1991, Mookerji and Rao 1994).
Several algal species have been used while testing the use of freshwater rotifer species. Since algal cultivation under controlled conditions is laborious, time consuming and expensive, alternative food types such as yeast, wastewater from food industry and livestock have been used for mass rotifer cultures (Klekot and Klimowicz 1981). However, controlled laboratory experiments using these food types for rotifers are necessary to compare the population growth with conventional algal diets. It is also not known whether different Brachionus species show different population growth rates when grown on yeast. At the same time, a comparative information about the growth of different rotifers grown on alga, yeast and their mixture separately has rarely been published (Guevara et al. 1996).
The aim of this study was to test the effect of different concentrations of green alga and yeast and their mixture on the population growth of two brachionid rotifer species commonly found in freshwater systems.
Materials and methods
We used clonal populations of each of the two rotifer species Brachionus calyciflorus and Brachionus patulus maintained at least for 3 months prior to testing. Both the rotifer species were mass-cultured (40 l glass aquaria) using the green algae Chlorella vulgaris as the exclusive food. Two weeks prior to experimentation, the test rotifer species were also offered baker's yeast Saccharomyces cerevisiae in addition to the algal diet. For maintaining mass rotifer cultures we used reconstituted hardwater (EPA) as medium (Anonymous 1985). This was also used as medium for rotifer growth experiments.
Chlorella was cultured using Bold-basal medium. Log phase algae were harvested, centrifuged and resuspended in EPA medium. Commercially available baker's yeast was freshly procured, resuspended in EPA medium and filtered using a 20 ![]() m mesh to remove clumps. Algal and yeast cell density was estimated using haemocytometer. For each rotifer species we offered food in the following ways: 1. only alga, 2. only yeast, and 3. alga+yeast in equal density.
m mesh to remove clumps. Algal and yeast cell density was estimated using haemocytometer. For each rotifer species we offered food in the following ways: 1. only alga, 2. only yeast, and 3. alga+yeast in equal density.
Each food type was offered in two densities viz. low (1x106) and high (3x106 ind. ml-1). For each food type and density, we maintained four replicates. Thus for population growth of B. calyciflorus, we maintained a total of 24 plastic jars (3 food types x 2 food concentrations x 4 replicates), each containing 20 ml of EPA medium with appropriate food density. The initial density of rotifers in each test jar was 5 ind. ml-1. For the population growth of B. patulus also the above design was used. Experiments were conducted at 23±2°C.
Following inoculation of B. calyciflorus or B. patulus, at every 24 h interval, we counted the number of female rotifers alive under a stereomicroscope. For this we either counted the whole volume of the test vessel or two aliquot samples each of 1-5 ml depending the density of rotifers per container. After estimating the density, the individuals were transferred to fresh EPA medium containing appropriate food type. The transfer of rotifers to fresh medium was done either individually, when the densities were low, or using a 50 ![]() m mesh during later stages of the study. Experiments were terminated after 15 days when most populations began to decline. Thoughout this study males were not encountered.
m mesh during later stages of the study. Experiments were terminated after 15 days when most populations began to decline. Thoughout this study males were not encountered.
The rotifer population growth was obtained from a mean of 4-5 values during the exponential phase using the equation r = (ln Nt - ln No)/t, where, No = initial population density, Nt = density of population after time t (days) (Krebs, 1985).
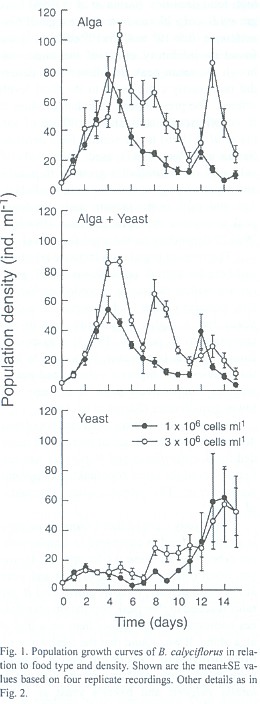
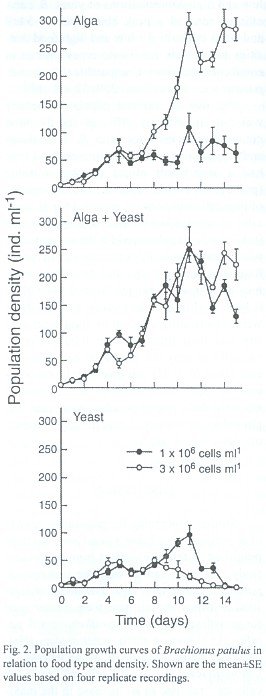
Results
The population growth curves of B. calyciflorus and B. patulus reared under three food types and two densities are shown in Figs. 1 and 2. The maximal population density of B. patulus influenced significantly (p<0.01) by food type, its concentration as well as their interaction but not for B. calyciflorus (p>0.05, ANOVA). In general, at any food type or concentration, B. patulus reached higher population density when compared to B. calyciflorus. In 1x106 ind. ml-1, B. calyciflorus reached 77±12 ind. ml-1; at the same food level B. patulus attained 109±26 ind. ml-1. At 3x106 ind. ml-1 density, B. calyciflorus reached a peak density of 103±8 ind. ml-1. Under comparable conditions, B. patulus reached much higher peak abundance 296±20 ind. ml-1. When yeast was used as exclusive food, the maximal abundance values reached by B. calyciflorus were 62±19 and 57±25 ind. ml-1 under low (1x106 ind. ml-1) low and high (3x106 ind. ml-1) food concentrations, respectively. On the other hand, B. patulus showed peak population abundances of 97±17 and 50±6 ind. ml-1 in low and high concentrations of yeast. B. calyciflorus reached a peak abundance of 54±9 and 86±3 ind. ml-1 for low and high food densities when both these food types offered in equal concentrations. Comparable values of B. patulus were 251±12 and 259±32 ind. ml-1.
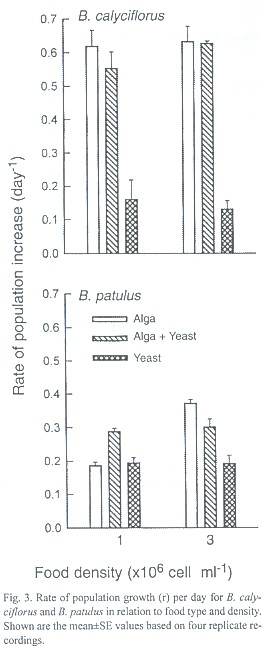
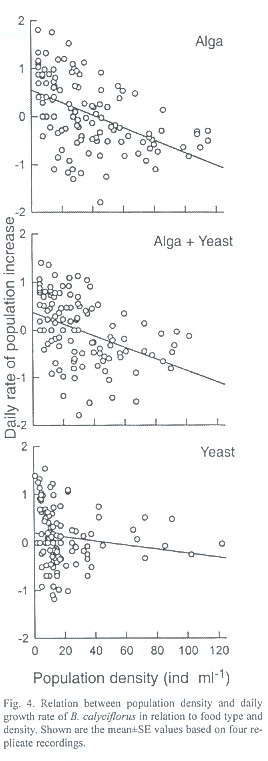
The day of maximal population density was not significantly different for the food concentrations used for both B. calyciflorus and B. patulus (p>0.05). However, food type had a significant effect on this variable (p<0.01) for both the species. The highest rate of population growth (r) recorded for B. calyciflorus was 0.63±0.04 and the lowest r value (0.13±0.03) was observed for the same species when grown in high concentration of yeast. Regardless of food type and density the r values of B. patulus ranged from 0.19±0.01 to 0.37±0.01 (Fig. 3). The r values for B. patulus were significantly affected by food type, density and their interaction (p<0.001). For B. calyciflorus only the food type had a significant effect (p<0.01). An inverse relation occurred for both B. calyciflorus and B. patulus when the daily rate of population increase was plotted againt the population density of the same day (Figs. 4 and 5).
| Species | Experiment | Food type | Food level | r value | Reference |
| Anuraeopsis fissa | Population growth | Scenedesmus obliquus | 0.5 - 8 x 106 cells ml-1 | 0.45 - 0.86 | Dumont et al. , 1995 |
| Population growth | Scenedesmus acutus | 0.5 - 40.5 x 106 cells ml-1 | 0.44 - 0.88 | Sarma et al. , 1996 | |
| Life table | - | - | 0.10 | Ooms-Wilms, 1997 | |
| Brachionus angularis | Population growth | Stichococcus bacillaris | - | 0.58 | Walz, 1993 |
| B. calyciflorus | Life table | - | - | 2.20 | Wang and Li, 1997 |
| Population growth | Scenedesmus acutus | 0.5 - 40.5 x 106 cells ml-1 | 0.79 -1.49 | Sarma et al. , 1996. | |
| Population growth | Various types of algae | - | 0.80 | Rothhaupt, 1990 | |
| B. patulus | Life table | Chlorella | 1-4 x 106 cells ml-1 | 0.14-0.61 | Sarma and Rao, 1991 |
| Population growth | Chlorella | 1-3 x 106 cells ml-1 | 0.12-0.24 | Sarma and Rao, 1990 | |
| B. plicatilis | Population growth | Tetrathelmis tetrathele | 0.05 x 106 cells ml-1 | 0.24 - 0.49 | Okauchi and Fukusho, 1984 |
| Population growth | Chlorella | 1.5 x 106 cells ml-1 | 0.16-0.47 | Okauchi and Fukusho, 1984 | |
| B. rubens | Population growth | Various types of algae | - | 0.80 | Rothhaupt, 1990 |
| Population growth | Chlorella | 3 x 106 cells ml-1 | 0.79 | Iyer and Rao, 1993 | |
| B. urceolaris | Life table | - | - | 1.32 | Wang and Li, 1997 |
| Keratella cochlearis | Population growth | Cryptomonas erosa | 0.005-0.01 x 106 cells ml-1 | 0.28-0.40 | Smith and Gilbert, 1995 |
| Population growth | - | - | 0.13 | Ooms-Wilms, 1997 | |
| K. crassa | Population growth | Cryptomonas erosa | 0.005-0.01 x 106 cells ml-1 | 0.32 | Smith and Gilbert, 1995 |
| K. testudo | Life table | - | - | 0.15 -0.39 | Stemberger, 1988 |
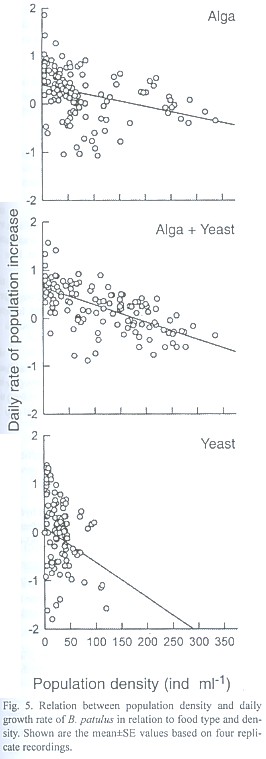
Studies concerning the population growth of B. patulus using baker's yeast have not been published so far. It is evident from the present study that yeast can be used for culturing B. patulus together with alga. The range of algal food densities chosen here were earlier used for growth studies of B. calyciflorus and B. patulus. As shown in many other studies (Halbach and Halbach-Keup 1974), an increase in Chlorella level from 1x106 ind. ml-1 to 3x106 ind. ml-1 resulted in an increase in the maximum population abundance of both B. calyciflorus and B. patulus (Figs. 1 and 2). However, when different food types were combined, the maximum peak density of B. calyciflorus was not statistically significant due to similar growth curves of rotifers fed yeast in low and high food densities. Sarma et al. (1996) have grown B. calyciflorus in a wide range of Scenedesmus (0.5x106 to 40.5x106 ind. ml-1) and found no inhibitory effect of the algae, although the mean peak population abundances did not exactly correspond to the food levels offered. The present peak abundance values of B. calciflorus are comparable to those of Sarma et al. (1996) under similar food densties. Sarma and Rao (1987) used 1x106 - 4x106 ind. ml-1 of Chlorella for growing B. patulus and reported peak abundance values of 110 - 325 ind. ml-1. In the present study, we found peak abundance values ranged from 109±26 to 296±20 depending on the algal food level.
The rates of population increase (r) observed here for both B. calyciflorus and B. patulus are within the range recorded earlier for Brachionidae (Table 1). In general, B. calyciflorus has a higher population growth rate compared to B. patulus. This is also evident from Fig. 3. It is however, important to note that species with higher r values need not always be competitively superior to those with lower growth rates (Sarma et al. 1999).
The inverse relation between population density and per capita rate of increase as recorded for B. calyciflurus and B. patulus was earlier observed for other zooplankton (Daphnia: Kerfoot et al. 1985, Anuraeopsis: Dumont et al. 1995).
This study showed that Chlorella vulgaris is a superior food as compared to Saccharomyces cerevisiae for these rotifer species. However, when offered a mixture of alga and yeast at low food density, B. calyciflorus and B. patulus reached higher peak population abundances comparable to or higher than on a diet of alga alone. It was found that the freshwater rotifers Brachionus calyciflorus and B. patulus were able to grow well on a mixed diet of Chlorella vulgaris and baker's yeast at 1x106 ind. ml-1 density. Only yeast was not suited for both rotifer species under 1x106 and 3x106 ind. ml-1 density. Thus, although a diet of yeast alone was not comparable to that of Chlorella, it can be effectively used at low concentrations to supplement algal requirements in rotifer culture systems.
Acknowledgements
SSSS and SN thank the National System of Investigators, Mexico (SNI-18723 and 20520, respectively) for support. Jose Luis G. Flores helped with the Spanish summary.
Resumen
Se comparó el crecimiento poblacional de dos especies planctónicas (B. calyciflorus y B. patulus) desarrolladas con el alga verde Chlorella vulgaris, la levadura de cerveza Saccharomyces cerevisiae y la mezcla de ambas dietas en proporciones iguales. B. patulus alcanzó las mayores densidades con cualquier tipo de alimento utilizado en comparación con B. calyciflorus. La dieta a base de Chlorella vulgaris sola promovió el mayor crecimiento poblacional en relación con la dieta de levadura sola. B. calyciflorus y B. patulus alcanzaron las mayores densidades de 103±8 ind. ml-1 y 296±20 ind. ml-1, respectivamente, con la dieta de Chlorella en 3x106 células ml-1. En condiciones similares, los valores máximos de abundancia de B. patulus fueron semejantes para ambos tipos de alimento. La tasa de incremento poblacional por día (r) para B. calyciflorus vario de 0.13±0.03 a 0.63±0.04, sin importar el tipo y densidad de alimento. Los resultados indican que la dieta a base de Chlorella fue mejor para los rotíferos considerados, y que la levadura puede usarse de manera efectiva a concentraciones bajas para complementar los requerimientos algales del sistema de cultivo de rotíferos.
References
Anonymous. 1985. Methods of measuring the acute toxicity of effluents to freshwater and marine organisms. US Environment Protection Agency EPA/600/4-85/013. [ Links ]
Dumont, H. J., S. S. S. Sarma & A. J. Ali. 1995. Laboratory studies on the population dynamics of Anuraeopsis fissa (Rotifera) in relation to food density. Freshwater Biol. 33: 39-46. [ Links ]
Fernandez-Reiriz, M. J. & U. Labarta. 1996. Lipid classes and fatty acid composition of rotifers (Brachionus plicatilis) fed two algal diets. Hydrobiologia 330: 73-79. [ Links ]
Guevara, M., A. G. Gaspar & N. Marin. 1996. The use of microalgae and baker's yeast in the culture of Brachionus plicatilis from the Araya's saline. Acta Cientifica Venezolana 47: 255-261. [ Links ]
Halbach, U. & G. Halbach-Keup. 1974. Quantitative beziehungen zwischen phytoplankton und der populationsdynamik des rotators Brachionus calyciflorus Pallas. Befunde aus laboratoriumsexperimenten und freilanduntersuchungen. Arch. Hydrobiol. 73: 273-309. [ Links ]
Hirata, H. & Y. Mori. 1967. Cultivation of the rotifer Brachionus plicatilis fed on a mixed diet of marine Chlorella and baker's yeast. Saibai Gyigyo 5: 36-40. [ Links ]
Iyer, N. & T. R. Rao. 1993. Effect of the epizoic rotifer Brachionus rubens on the population growth of three cladoceran species. Hydrobiologia 255/256: 325-332. [ Links ]
Kerfoot, W.C., W. R. Demott & C. Levitan. 1985. Non-linearities in competitive interactions: component variables or system response? Ecology 66: 959-965. [ Links ]
Klekot, L. & H. Klimowicz. 1981. Rotifer communities of ponds supplied with post-waste water. Holarctic Ecol. 4: 1981. 208-214. [ Links ]
Krebs, C. J. 1985. Ecology. The experimental analysis of distribution and abundance. 3rd edn. Harper and Row, New York. 789 pp. [ Links ]
Lie, O., H. Haaland, G. I. Hemre, A. Maage, E. Lied, G. Rosenlund, K. Sandnes & Y. Olsen. 1997. Nutritional composition of rotifers following a change in diet from yeast and emulsified oil to microalgae. Aquacult. Int. 5: 427-438. [ Links ]
Mookerji, N. & T. R. Rao. 1994. Influence of ontogenetic changes in prey selection on the survival and growth of rohu, Labeo rohita and singhi, Heteropneustes fossilis larvae. J. Fish Biol. 44: 479-490. [ Links ]
Okauchi, M. & K. Fukusho. 1984. Food value of minute alga, Tetraselmis tetrathele, for the rotifer Brachionus plicatilis culture: 1. Population growth with batch culture. Bull. Nat. Res. Inst. Aquacult. 5: 13-18. [ Links ]
Ooms-Wilms, A.L. 1997. Are bacteria an important food source for rotifers in eutrophic lakes? J. Plankton Res. 19: 1125-1141. [ Links ]
Rothhaupt, K.O. 1990. Population growth rates of two closely related rotifer species effects of food quantity particle size and nutritional quality. Freshwater Biol. 23: 561-570. [ Links ]
Rottmann, R. W., J. V. Shireman & E. P. Lincoln. 1991. Comparison of three live foods and two dry diets for intensive culture of grass carp and bighead carp larvae. Aquaculture 96: 269-280. [ Links ]
Sarma, S. S. S. & T. R. Rao. 1987. Effect of food level on body size and egg size in a growing population of the rotifer Brachionus patulus Müller. Arch. Hydrobiol. 111: 245-253. [ Links ]
Sarma, S.S.S. & T. R. Rao. 1990. Population dynamics of Brachionus patulus Müller (Rotifera) in relation to food and temperature. Proc. Indian Acad. Sci. (Anim. Sci.) 99: 335-343. [ Links ]
Sarma, S.S.S. & T. R. Rao. 1991. The combined effects of food and temperature on the life history parameters of Brachionus patulus Müller (Rotifera). Int. Revue ges. Hydrobiologia 76: 225-239. [ Links ]
Sarma, S. S. S., N. Iyer & H. J. Dumont. 1996. Competitive interactions between herbivorous rotifers: Importance of food concentration and initial population density. Hydrobiologia 331: 1-7. [ Links ]
Sarma, S.S.S., M. A. Fernández-Araiza & S. Nandini. 1999. Competition between Brachionus calyciflorus Pallas and Brachionus patulus (Müller) (Rotifera) in relation to algal food concentration and initial population density. Aquatic Ecol. 33: 339-345. [ Links ]
Smith, A. D. & J. J. Gilbert. 1995. Relative susceptibilities of rotifers and cladocerans to Microcystis aeruginosa. Arch. Hydrobiol. 132: 309-336. [ Links ]
Stemberger, R. S. 1988. Reproductive costs and hydrodynamic benefits of chemically induced defenses in Keratella testudo. Limnol. Oceanogr. 33: 593-606. [ Links ]
Walz, N. 1993. Carbon metabolism and population dynamics of Brachionus angularis and Keratella chochlearis. In: N. Walz (ed). Plankton regulation dynamics. Experiments and models in rotifer continuous cultures. Springer-Verlag, Berlin. p. 89-105. [ Links ]
Wang, J. & D. Li. 1997. Comparative studies on principal parameters of population growth of five freshwater rotifers. Acta Hydrobiol. Sin. 21: 131-136. [ Links ]
1 Carrera de Biología, UNAM Campus Iztacala, AP 314, CP 54090, Los Reyes, Tlalnepantla, Edo. de México, México. Fax: +52 5 623 1155, e-mail: sarma@servidor.unam.mx
2 CyMA Project.
*Corresponding author

