










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.49 n.1 San José Mar. 2001
Alicia González Solis 1 y Daniel Torruco Gómez 1
Recibido 24-III-1999. Corregido 3-VIII-2000. Aceptado 19-IX-2000
Abstract
The fish and invertebrates community structure in the Sabancuy estuary was analyzed in two seasons and 14 sampling stations (13 along the estuary and one in the marine adjacent coast). No significant differences were found between seasons. The environmental frame defines two zones within the estuary, the first extends from the access highway to Sabancuy town until the Pujo mouth in the west; the second from the bridge to the estuary head in the east. The most abundant invertebrates were mollusks (51.8% of the total), in biomass the crustaceans dominated. The fish included 21 families and 33 species; the most abundant were Gerridae, Scianidae, Sparidae, Lutjanidae and Ciprinodontidae. The highest diversities of both communities correspond to the central part of the estuary. These communities include three sections with notable differences in faunal distribution: one is influenced by the exit to Terminos lagoon, the secondary in the estuary head and a third is in a transition zone defined by the proximity of the town access bridge. The ecological organization suggests a strong division caused by the bridge, both sides are scarce in habitats and nutrient resources and this is reflected in the low species counts.
Keywords: Crustaceans, ictiofauna, mollusks, Sabancuy Estuary, Campeche, Mexico.
Las lagunas costeras y estuarios son una zona de interfase con límites o gradientes de dimensiones variables en escala, espacio y tiempo (Margalef 1995). En el Sureste de México existe un interés creciente en estos ambientes debido a su gran potencial de utilidad (Villalobos y Zamora 1975). Estos ecosistemas presentan alta variabilidad ambiental y cambios regulares e impredecibles (Escanero 1983). Son interdependientes de sistemas aledaños como manglares, pantanos, playas y marismas, por lo que su estudio y administración de recursos requiere de un equipo multidisciplinario (Yáñez-Arancibia y Sánchez-Gil 1988). México cuenta con extensos litorales, cerca del 15% corresponde a lagunas costeras con una área estimada de 1 225 500 ha (Peralta 1984, Contreras 1993), en donde se han realizado estudios químicos y geomorfológicos (Graham et al. 1981, Day et al. 1982). En estos litorales del 70 al 80% de las especies capturadas comercialmente, pasan alguna etapa de su vida en las lagunas costeras adyacentes (Chávez y Torruco 1988, Contreras 1993). Presentan importantes áreas de anidación y reproducción de especies de aves, lo que a dado origen a convenios internacionales de conservación. Son zonas de gran importancia turística, actual o potencial ya que constituyen áreas de alto interés recreativo.
En la península de Yucatán han sido estudiadas intensamente (Lankford 1976, Gómez-Aguirre 1974, Mancilla y Vargas 1980), sin embargo son pocas las zonas con estudios continuos. En la mayoría de estos ecosistemas se presenta un alto impacto, debido a las actividades antropogénicas propias del desarrollo costero, como son la construcción de estructuras de diversos tipos y la eliminación de desechos orgánicos (Swartz et al. 1986), alteraciones que han afectado a los componentes bióticos y en algunos casos a la dinámica hidrológica con cambios en sus características físicas y químicas. Bajo esta diversidad de actividades, es básico para el entendimiento de su dinámica y la utilización racional de sus recursos, el conocimiento de la estructura y función de sus comunidades. En consecuencia en el presente estudio se analiza la caracterización biótica de la fauna béntica: peces y macroinvertebrados del estero de Sabancuy, Campeche, México.
Materiales y métodos
El estero de Sabancuy se localiza a 18o58' de latitud y 91o12' de longitud en el estado de Campeche, México (Fig. 1). El área está comprendida fisiográficamente dentro de la península de Yucatán (Psuty 1966), algunos autores lo consideran perteneciente a la planicie fluvial del suroeste de Campeche (Carranza-Edwards et al. 1979). Está integrado por cinco áreas importantes: El Pujo, San Nicolás, Ensenada Polcai, Santa Rosalia y el área del poblado donde es característico un puente de acceso (Fig. 1). Presenta dos bocas o canales de acceso, hacia el Pujo se localiza el canal que desemboca en la Laguna de Términos, formando lagunas y pantanos con extensas planicies de inundación, la zona es muy baja y con elevaciones menores a 40 m. La segunda boca o canal se localiza hacia la ensenada Polcai, es un canal artificial que desemboca hacia el área costera adyacente o frente marino. El clima de la región según Köppen es Aw1(i)g- cálido subhúmedo con lluvias en verano, su régimen de precipitación anual es de 1426.8 mm y el de temperatura de 26.9 °C (García, 1973). La hidrología superficial presenta numerosos bajos anegadizos con aguadas en sus fondos hundidos o pequeñas lagunas permanentes y alto contenido de sales en solución. En los alrededores las elevaciones son menores a diez metros con una pequeña inclinación de los terrenos hacia la costa, lo que origina extensas planicies con pequeñas ondulaciones denominadas cuyos, que son pequeños montículos donde cambia la condición del suelo.
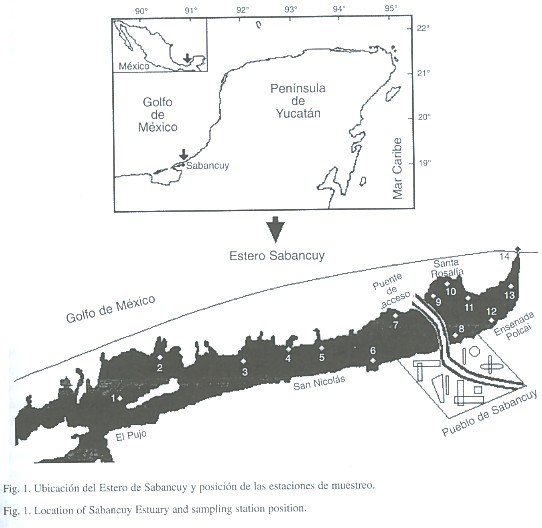
Se realizaron dos campañas de muestreo: la primera a finales de la época de sequías (mayo de 1997) y la segunda al inicio de la época de lluvias (julio de 1997). En cada muestreo se efectuaron catorce estaciones: trece a lo largo del estero y una a la salida del estuario en el área costera adyacente (Fig. 1). En cada estación se realizaron mediciones de temperatura, salinidad y oxígeno disuelto mediante un salinómetro-oxímetro YSI8I. La medición de nutrimentos se llevó acabo según el método estándar de Strickland y Parson (1972). Las muestras de sedimento se recolectaron con un nucleador de diez centímetros de diámetro a una profundidad de diez centímetros. La determinación granulométrica se hizo con la técnica de Buyoicos y la de materia orgánica mediante combustión de la fracción orgánica. En el análisis se utilizaron los promedios ambientales como un marco de referencia. La macrofauna se obtuvo mediante lances de quince minutos con una red camaronera de una luz de malla de 1.5 pulgadas (3.18 cm), a su vez se realizó en cada estación una réplica. Los ejemplares obtenidos fueron identificados, contados y pesados en el laboratorio. En ambos muestreos se realizaron pruebas univariadas de comparación estadística, para evaluar si las diferencias eran significativas entre ellos, al no encontrar diferencias (
En la caracterización de la estructura faunística se utilizaron indicadores de dominancia y diversidad que permitieron identificar las diferentes categorías y su importancia relativa en la comunidad. En la dominancia se utilizó el índice de valor de importancia (Cox 1976), que ponderó la frecuencia, abundancia y biomasa de las especies. La diversidad se evaluó con el índice de Simpson (Goulden 1960, Galluci 1973). El método seleccionado en la descripción de los patrones comunitarios fue de afinidad bajo el índice de Morisita (Orlóci 1978), que es un método numérico efectivo que facilita la comprensión de los resultados (Gnanadesikan 1977, Pielou 1984, Chang y Gauch 1986), en este procedimiento se utilizaron datos de abundancia y biomasa de ambos grupos faunísticos: peces e invertebrados.
Resultados
Marco ambiental: Con los muestreos hidrológicos fue posible definir dos zonas dentro del estero de Sabancuy. La primera fue la de mayor superficie y va del puente de acceso del pueblo hasta el Pujo, donde desemboca el estero hacia el complejo lagunar de Términos. La segunda fue la parte que va del puente de acceso hacia la ensenada Polcai. Se consideró como una tercera zona, las inmediaciones próximas al canal o boca de la ensenada Polcai (Fig. 2). La laguna es muy somera con profundidades de uno a dos y medio metros, su escasa circulación de agua origina en algunas áreas alta sedimentación, lo que contribuye a que áreas como el Pujo tengan gran cantidad de bajos que dificultan el tránsito (Fig.1). La temperatura que se registró es bastante homogénea con valores que van de 22 a 32°C, presentándose las más altas en áreas hacia el Pujo con baja profundidad y alta insolación. En el frente marino hacia la ensenada Polcai la temperatura fue más constante con fluctuaciones alrededor de los 30°C. El sistema presentó en ambas épocas del año características marinas con salinidades de 32 en la zona interna y 37 en el Pujo. En la zona interna de inundación los escurrimientos de agua dulce favorecen salinidades más bajas. En el frente marino está variable fue homogénea con alrededor de 38 . En relación con el oxígeno disuelto, la concentración dentro de la laguna fue de 3.1 a 4.8 ml/l lo que significa valores normales, el área de San Nicolás registró alta concentración con sobresaturación de este elemento (4.8 ml/l; 102-108%).
En el frente marino se registraron altos valores de saturación (4.9 ml/l; 112%). La concentración de nitratos tuvo variaciones que van de 0.9-2.1 ug-at N/l, con los valores más bajos entre las estaciones tres y seis del sistema (0.9-1.7 ug-at N/l). En el caso de los nitritos las concentraciones van desde 0.3 a 0.6 ug-at N/l, en el frente marino fue muy homogénea (0.41 ug-at N/l). Sin embargo el amonio se encontró en altas concentraciones con un gradiente que va desde el Pujo (40.0 ug-at N/l) a San Nicolás (27.4 ug-at N/l), disminuyendo la concentración hacia el puente de acceso (14.9 ug-at N/l) y la ensenada Polcai (20.2 ug-at N/l). Los fosfatos también presentaron un gradiente con las concentraciones más altas en la ensenada Polcai (5.2 ug-at P/l) y las más bajas hacia el Pujo (12 ug-at P/l). En los silicatos el gradiente fue inverso, las mayores concentraciones se presentaron en el Pujo (48.6-54.7 ug-at Si/l) y las menores hacia la ensenada Polcai (22 ug-at Si/l).
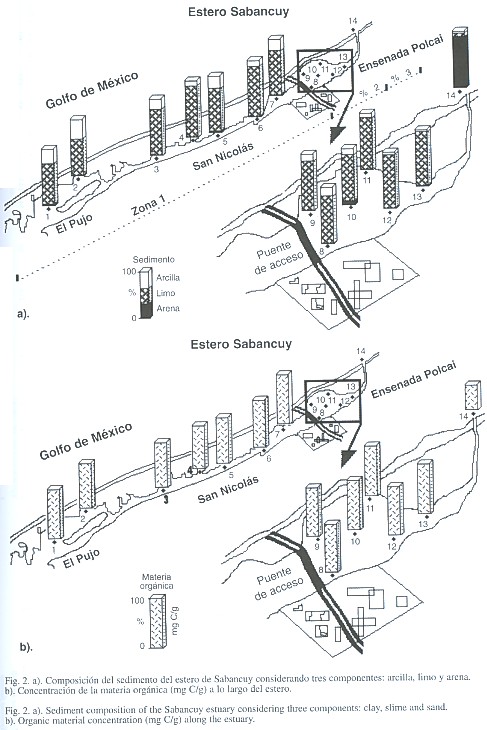
En el análisis granulométrico se observó que los sedimentos del estero están compuestos de arenas, arenas finas, limos arenosos y arcillosos, frecuentemente con restos de moluscos. El sedimento arenoso se encontró principalmente en la zona marina con una marcada tendencia al aumento de diámetro en las áreas cercanas a la costa (Fig. 2 a). El sedimento más común en el interior del estero es el limo con varios grados de mezcla, más arenoso de la ensenada Polcai hacia el frente marino (estación catorce). El sedimento limo-arcilloso se presentó entre San Nicolás y el Pujo con un diámetro más pequeño. La clasificación granulométrica marcó la tendencia a concentrar el sedimento limo-arcilloso hacia las orillas, mientras que en el canal de navegación fue limoso o limo-arenoso. El substrato del sistema presentó una capa muy delgada de sedimento no consolidado, aparentemente la placa de roca calcárea subyacente es superficial. En el análisis de materia orgánica los registros de 7 mg C/g fueron semejantes en todo el cuerpo lagunar. En el Pujo se registraron los valores más altos, en concordancia con el sedimento limo-arcilloso. Sin embargo las estaciones cercanas a la ensenada Polcai también registraron altos valores a pesar de que su sedimento fue más arenoso. Los canales de navegación y San Nicolás presentaron en promedio 6 mg C/g, mientras que la estación catorce del frente marino registró el valor más bajo 3.5 mg C/g (Fig. 2 b)
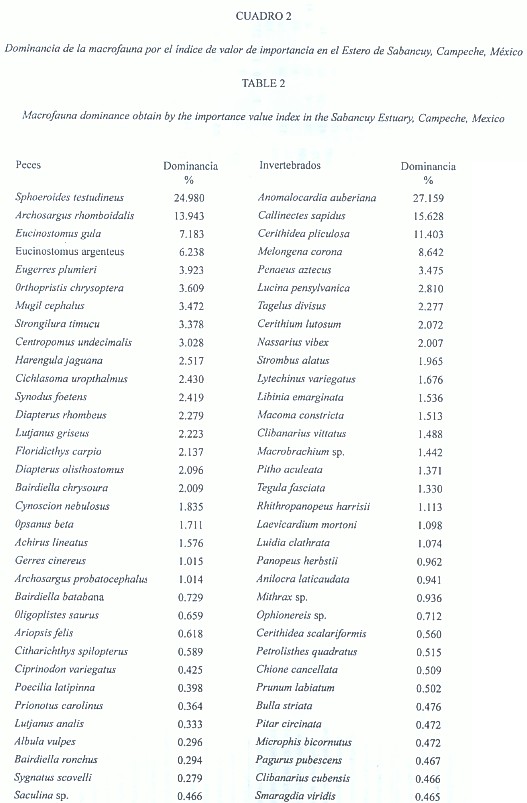
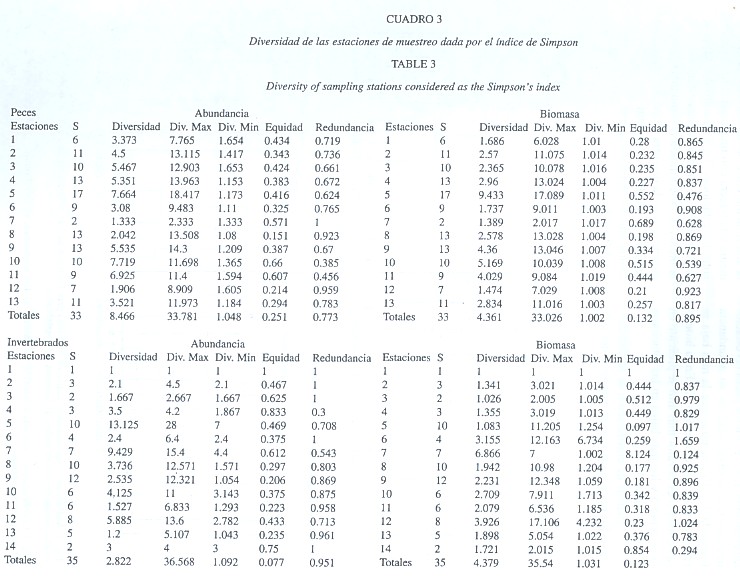
Marco faunístico: Se registró una abundancia total de 2 179 ejemplares de macrofauna con una biomasa de 42 232.05 g, repartidos en 1 385 ejemplares (39 992.8 g) de peces y 794 (2 239.25 g) de invertebrados. Se obtuvieron 68 especies, 33 de peces y 35 de invertebrados: 17 especies de moluscos, 15 de crustáceos y 3 de equinodermos (Cuadro 1). En los invertebrados las familias Cerithiidae y Chionidae fueron las de mayor abundancia, en cuanto a la biomasa las jaibas (Portunidae) aportaron los valores más altos, a pesar de que los Melongenidae y Chionidae presentaron valores importantes. En los peces los Tetrodontidae y Sparidae fueron más frecuentes y proporcionaron la mayor biomasa. Las especies restantes de la macrofauna presentaron abundancias y biomasas medias o esporádicas. Las especies de peces más importantes fueron Sphoeroides testudineus y Archosargus rhomboidalis con el 38.92 % del total. En los invertebrados correspondió a las especies Anomalocardia auberiana (molusco), Callinectes sapidus (crustáceo) y Cerithidea pliculosa (molusco) con el 54.19 % de la dominancia (Cuadro 2). La mayor diversidad en la abundancia de los invertebrados se registró en las estaciones 5 y 7; los valores más bajos se registraron en las estaciones 1 y 13. Con los datos de biomasa, el valor más alto se presentó en la estación 7 y el más bajo en las estaciones 1, 3 y 5. En los peces las estaciones con mayor diversidad en su abundancia fueron la 5 y 10, mientras que la más baja se registró en las estaciones 7 y 12. En la biomasa de los peces el valor más alto se presentó en la estación 5 y los más bajos en las estaciones 7 y 12 (Cuadro 3).
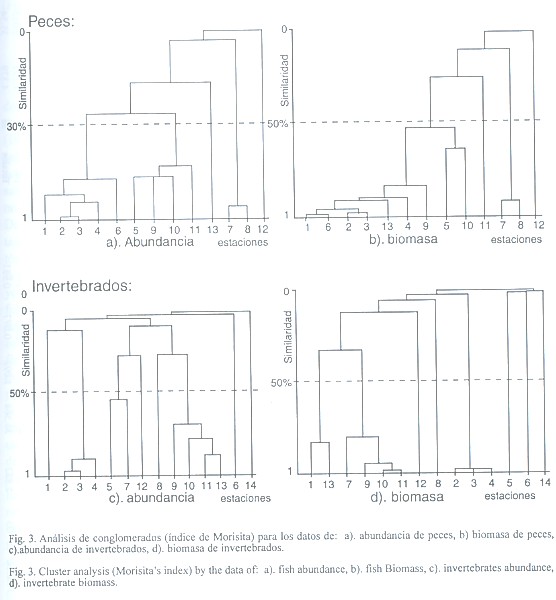
El análisis de clasificación de la abundancia de los peces a un nivel de similitud del 50% presentó tres agrupaciones. La primera con estaciones de la parte oeste, la segunda evidenció la alta relación entre las estaciones de la parte media y la tercera con una asociación paritaria entre las estaciones 7 y 8 que están próximas al puente de acceso del poblado (Fig. 3 a). En la biomasa, la ictiofauna presentó dos grupos; el primero con la mayoría de las estaciones y el segundo al igual que en la abundancia presentó una asociación paritaria entre sus estaciones 7 y 8 (Fig. 3 b).
La abundancia de los invertebrados permitió reconocer tres grupos. El primero reúne a las estaciones 2, 3 y 4 ubicadas al oeste del estero, el segundo a las estaciones 5 y 7 de la parte media y el tercer grupo une a las estaciones (9, 10, 11, 13) localizadas al este (Fig. 3 c). Con los registros de biomasa también se presentaron tres conglomerados, el primero que une a estaciones de las bocas del estero, el segundo fue más numeroso e incluye a estaciones muy cercanas al puente y el tercero une consistentemente a estaciones de la parte oeste (Fig. 3 d).
Discusión
Marco ambiental: Los valores registrados de oxígeno se encuentran dentro de los intervalos obtenidos por Villalobos y Zamora (1975) para la Sonda de Campeche. La sobresaturación registrada es un indicador de la alta actividad fotosintética de la zona. La más baja saturación observada fue del 70% por lo cual no hay déficit de este elemento en el sistema. De las principales fuentes de nitrógeno reportadas, las concentraciones no son limitantes ni tóxicas para el desarrollo planctónico y sólo en la parte media del estero los valores disminuyen, debido a la homogeneidad de la actividad fotosintética por el fitobentos. El amonio presentó un gradiente que es producto de la remineralización de la materia orgánica, ya que es fácilmente asimilado por los productores primarios, la menor concentración en el área del puente puede ser indicio de un incremento en la tasa de producción. Los valores reportados en la zona marina adyacente dan evidencia de un aporte de la laguna hacia el mar por acción de la marea fenómeno ya señalado por Bravo y Yáñez-Arancibia (1979). Los fosfatos no son limitantes; sin embargo, las altas concentraciones cerca del puente de acceso y en la boca del estero podrían ser consecuencia de la gran cantidad de detergentes que se vierten al sistema por los drenajes urbanos provenientes del pueblo. Asimismo, es probable que los fertilizantes químicos que se utilizan en las actividades agrícolas de los campos adyacentes sean una fuente importante de fosfatos. Los silicatos, al tener un ciclo más lento que los anteriores elementos, podría acumularse en El Pujo a causa de la baja circulación; en la zona urbanizada, por el contrario este elemento es exportado hacia la zona marina adyacente por las corrientes de marea (Suárez y Gómez 1965).
El sedimento arenoso cercano al frente marino se consideró está asociado a la turbulencia de las masas de agua, donde los gránulos son transportados por las corrientes y el oleaje sobre la costa, su extensión dentro de la laguna se debe también al transporte eólico y a las corrientes de marea, asociados a los canales de entrada y salida al estero. Los sedimentos limosos y los mixtos, mal clasificados (diámetros entre 1 y 2 micras) están ampliamente distribuidos en la laguna, particularmente en los extremos paralelos a la orilla continental, es probable que su deposito haya sido el resultado del drenaje superficial y la erosión costera. Todo el estero se encuentra rodeado de mangle y es evidente que la cantidad de materia orgánica está íntimamente relacionada con el aporte que proporciona esta vegetación. Al oeste del estero no existe un gran aporte de materia orgánica debido principalmente al escaso flujo de marea. El área marina al ser un fondo arenoso presentó bajos valores, a pesar de la presencia de manchones de pastos marinos que podrían elevar los valores de materia orgánica.
Marco biológico: La alta dominancia que presentaron los moluscos: Anomalocardia auberiana y Cerithidea pliculosa y el crustáceo Callinectes sapidus, se consideró que está íntimamente relacionada con las condiciones del substrato, tanto en su granulometría y contenido de materia orgánica. En los peces, además de la influencia de estos factores, es probable que otros componentes de influencia marina originen la dominancia de Sphoeroides testudineus y Archosargus rhomboidalis, ya que la corriente de marea caracteriza casi en su totalidad al estero. La dominancia que presentan especies de estos grupos ayuda a ejemplificar las diferentes regiones y puede dar evidencia de la amplitud del prisma de mareas en esa zona (Woodin 1981). El patrón de la diversidad de los moluscos, crustáceos y peces, puede integrarse de la siguiente manera: la diversidad fue baja cerca del Pujo y aumentó hasta llegar a San Nicolás, posteriormente se tuvo un decremento con los valores más bajos en las proximidades del puente (estación 7), después del puente la diversidad se incrementó con los valores más altos en la ensenada Polcai considerada la zona más amplia del estero, asimismo conforme se estrecha el estero hacia el frente marino la diversidad baja considerablemente. Esta última reducción se consideró natural, debido a la relación directa de la superficie que involucra (Swartz et al. 1986), sin embargo parte de la reducción registrada cerca del puente se debe a las estructuras de concreto del mismo, que probablemente actúan como una barrera física al movimiento de aguas en el complejo estuarino y constituye un elemento importante de inestabilidad ambiental. Los valores de equidad siguen una progresión, se consideraron un indicador de la disponibilidad y utilización de recursos. Es posible que el puente actué de manera que evita la dominancia de algunas especies.
Considerando que el criterio de análisis utilizado para obtener la asociación de localidades pondera en cada caso los valores de densidad (número o biomasa), la riqueza específica y la presencia o ausencia de los elementos registrados para cada localidad, en el estero de Sabancuy se presentó una gran heterogeneidad en sus características biológicas. Lo heterogéneo del sistema fue más evidente al oriente, ya que al occidente fue ligeramente más homogéneo. De hecho, se identifican cinco estaciones al oriente que si bien algunas de ellas se asocian a otras por la presencia y densidad de las especies que las componen, se consideran áreas de crianza de juveniles. Dos estaciones identificadas con estas características son la 8 y 9 en la ensenada Polcai, cercanas a la comunicación con el mar. Ambas presentan hábitats adecuados para el crecimiento de Orthopristis chrysoptera y la 8 para el mojarrón Archosargus romboidalis. En esta parte de la laguna y hacia el final del estero (estación 13) Harengula jaguana encuentra condiciones excelentes para su crecimiento. Al occidente, Eucinostomus gula encontró condiciones favorables de la estación 5. La especie Sphoeroides testudineus mantuvo cierta preferencia por el área de la estación 6, aunque su registro fue consistente en todo el estero, otro aspecto interesante es que presentó al occidente del estero tallas más grandes, por lo que probablemente utiliza las dos porciones del sistema estuárico de manera diferencial para su crecimiento, como lo han postulado Yáñez-Arancibia y Nugent (1978) para otras especies de la laguna de Términos.
Con este esquema, es consistente la asociación de la porción oriental del estero con el frente marino. Las estaciones de la porción occidental (El Pujo) y la zona marina adyacente a la desembocadura se separan de otros agrupamientos quedando aisladas, lo que permite inferir que existen diferencias en el ambiente que influyen fuertemente en estas áreas. Sin embargo en la asociación por estaciones, la influencia directriz de estos cambios ambientales fue menos evidente y de ahí su alejamiento del grupo principal que se presentó. Es probable que las especies de estas estaciones respondan de otra manera a los cambios que imponen los controladores ecológicos de esa zona. Del análisis de los factores físicos y químicos del sistema estuarino, se muestra un gradiente originado por la salinidad y el fósforo o por la interacción de ambos factores, esto lleva a proponer la posibilidad que estos elementos sean los responsables de los patrones de abundancia obtenidos con la ictiofauna del estero. En los invertebrados se presentó una relación más estrecha con el patrón de materia orgánica registrado.
Consideraciones faunísticas generales: La macrofauna del estero de Sabancuy presentó ensamblajes significativos que invalidan una hipótesis de distribución faunística independiente entre sus localidades, ya que los agrupamientos entre localidades están determinados por requerimientos semejantes o antagonismos definidos en la utilización de recursos (espacio, hábitat, alimento), lo que implica que la presencia de una especie determine la presencia o ausencia de otra. En el estero, predominan especies eurihalinas del componente marino y especies temporales del componente estuarino, mientras que el componente dulceacuícola esta representado solo por dos especies de peces (Cichlasoma uropthalmus y Poecilia latipina). Este sistema presentó un comportamiento ligeramente hiperhalino. Uno de los aspectos que más destacó en el análisis, fueron las altas interacciones que existen entre las estaciones del área estuárica. En relación con las condiciones de baja profundidad en el estero, es probable que las tasas de recambio de oxígeno y fósforo entre la columna de agua con el sedimento sea muy fuerte en la parte norte donde existe comunicación con el mar, producto del flujo de marea. La biomasa de los invertebrados está determinada por un complejo de variables, en donde la mayor variación explicada se consideró que está dada por la composición del sedimento más que por la salinidad, ya que el flujo de nutrimentos en el sedimento juega un papel fundamental en el reciclaje de materia en estos sistemas someros, donde además las fragmentaciones de invertebrados debidas al oleaje contribuyen al enriquecimiento del detritus, que es parte importante de la cadena alimenticia del estero (Yáñez-Arancibia et al. 1982). De los escasos datos obtenidos de vegetación, se podría pensar que la producción de carbono orgánico por el fitobentos es alta, siendo éste una trampa de sedimentos y nutrimentos que logra máximas concentraciones en la materia orgánica.
Mann (1980) menciona que los sedimentos representan el almacén de los materiales formados en la columna de agua y que eventualmente una fracción se recicla vía el macro y microbentos. En Sabancuy, la remoción efectiva del sistema estuárico se debe fundamentalmente a la poca profundidad y a las fuerzas de la corriente que imperan en el estero (Yáñez-Correa, 1963). El esquema de organización ecológica que se registró en el complejo estuarino, implica la división del estero en dos porciones separadas por el puente de acceso. Ambos extremos de la laguna presentan pobreza de especies, al occidente (El Pujo) es el resultado de la baja profundidad y escasa comunicación con la Laguna de Términos. Hacía la ensenada Polcai en el oriente podría deberse a la topografía del estero, ya que al disminuir su amplitud se restringe la circulación de agua, a pesar de la influencia del prisma de mareas. Estos factores dan un resultado común: escasez de hábitats y de recursos alimenticios.
En relación con las interacciones biológicas, se infiere que existe depredación y competencia entre la macrofauna registrada y aunque fue un factor difícil de observar de manera directa (Orians y Colliers 1963), la dominancia es un indicador de que las especies se traslapan en la utilización de los recursos y da una idea de la magnitud del mismo (Mac Arthur y Levins 1967). En el trópico la supervivencia y persistencia de una especie depende de diversos recursos, tales como heterogeneidad de microhábitats, alta productividad, heterogeneidad espacial, competencia y depredación entre otros, es por ello las complejas relaciones que presentan los organismos que habitan estas regiones. Siempre que los límites son definidos por interacciones biológicas, el problema de estructurar estas comunidades consiste en identificar sus interacciones, así que una línea de inicio es el estudio de las características de los grupos de especies que se encuentren en el centro del intervalo y en fronteras coterminales, asimismo la macrofauna y sobre todo las especies dominantes son elementos susceptibles a ser seleccionados como indicadores de eventos o disturbios que se presenten de manera natural o artificial.
Según la literatura la hipótesis más factible es que la rapidez de los procesos es mayor en los trópicos que en regiones análogas de zonas templadas. Sin embargo, en ocasiones los datos resultan contradictorios y necesariamente las hipótesis generadas necesitan someterse a diversas pruebas y evaluaciones en su relación directa con la tasa metabólica de los sistemas contribuyentes, así como con las tasas de productividad, descomposición y recirculación, las cuales deben conocerse para valorar el rendimiento orgánico potencial, la capacidad natural de procesamiento de desechos y las alternativas de los procesos de degradación (Wilbur y Tiller 1970). El análisis de este conjunto de variables es vital a fin de idear y desarrollar las prácticas de manejo relativas a la recuperación del ecosistema, la sedimentación del estero y a la optimización o diseño de nuevos sistemas de explotación. El presente estudio será la base de esta nueva etapa.
Resumen
La estructura comunitaria de peces e invertebrados en el estero de Sabancuy fue analizada en dos temporadas y 14 estaciones de muestreo (13 a lo largo del estero y una en la costa marina adyacente). No se encontraron diferencias significativas entre las temporadas. El marco ambiental define dos zonas dentro del estero, la primera va de la carretera de acceso al pueblo de Sabancuy hasta la boca del Pujo en el oeste; la segunda del puente a la cabeza del estero en el este. Los invertebrados más abundantes fueron los moluscos (51.8 % del total) y en la biomasa los crustáceos. Los peces registraron 21 familias y 33 especies. Las familias más abundantes fueron Gerridae, Scianidae, Sparidae, Lutjanidae y Cipinodontidae. Las diversidades más altas de ambas comunidades corresponden a la parte central del estero. Estas comunidades presentaron tres agrupaciones con diferencias notables en la distribución de la fauna: una influenciada por la salida hacia la laguna de Términos, la segunda en la cabecera del estuario y la tercera es una zona de transición definida por la cercanía al puente de acceso del poblado. La organización ecológica implica una fuerte división debida al puente; ambos extremos presentan escasez de hábitats y recursos alimenticios, lo que se refleja en la pobreza de especies en esas áreas.
Referencias
Bravo, N. E. & A.Yáñez?Arancibia. 1979. Ecología de la Boca de Puerto Real Laguna de Términos, Campeche, México. An. Centro Cienc. del Mar y Limnol. U.N.A.M. 7: 69-118. [ Links ]
Carranza, Edwards, A., M. Gutiérrez & R. Rodríguez. 1979. Unidades morfotectónicas continentales de las costas mexicanas, México. An. Centro Cienc. del Mar y Limnol. U.N.A.M. 2: 81? 88. [ Links ]
Chang, D. H. & H. G. Gauch. 1986. Multivariate analysis of plant communities and environmental factors in Ngari, Tibet. Ecol. 67: 1568-1575. [ Links ]
Chávez, E. A. & D. Torruco. 1988. Actualización de los conocimientos y formulación de estrategias de manejo de lagunas costeras para pesca y acuicultura. Caso II: Sistema Carmen-Pajonal- Machona, Tabasco. Workshop proyecto Aquila/FAO, Venezuela.47 p. [ Links ]
Contreras, F. 1993. Ecosistemas Costeros Mexicanos. U.N.A.M., México. 217 p. [ Links ]
Cox, C. W. 1976. Laboratory manual of general ecology. W.C.Brown Pub. Dubuque Iowa. 215 p. [ Links ]
Day, J. W., R. H. Day, M. Barreiro, F. Le-Lou & C. Madden. 1982. Primary production in the Laguna de Términos a tropical estuary in the southern Gulf of Mexico. Proc. Inter. Sym. Coastal Lagoons. ISCOL-UNESCO. Acta 0ceanol. 269-276 p. [ Links ]
Escanero, F. G. 1983. Ciclo hidrográfico anual (marzo, 1981- abril 1982) de la Laguna de Términos, Campeche. Tesis Maestría, CICESE, México. [ Links ]
Galluci, U. F. 1973. 0n the principles of thermodynamics in ecology. Ann. Rev. Ecol. Sys. 4: 329?357. [ Links ]
García, E. 1973. Clasificación climática de Köppen. U.N.A.M., México. [ Links ]
Gnanadesikan, R. 1977. Methods for statistical data analysis of multivariate observations. Wiley Nueva York. 311 p. [ Links ]
Gómez-Aguirre, S. 1974. Reconocimientos estacionales de hidrología y plancton en la Laguna de Términos, Campeche, México (1964-1965). An. Centro Cienc. del Mar y Limnol. U.N.A.M. 1: 61-82. [ Links ]
Goulden, C. E. 1960. Temporal changes in diversity. 96-102. In: Diversity and stability in ecological systems. Brokhaven symposia in Biology National Technical Information Service. Springfield. [ Links ]
Graham, D. S., J. P. Daniels, J. M. Hill & J. W. Day. 1981. A preliminary model of the circulation of Laguna de Términos, Campeche, Mexico. An. Centro Cienc. del Mar y Limnol. U.N.A.M. 8: 51-62. [ Links ]
Lankford, R. R. 1976. Coastal Lagoons of Mexico, their origin and classification. p.182-215. In: Wiley, M. (ed.). Estuarine processes: Circulation, sediments and transfer of material in the Estuary. Estuarine Research federation conference, Galveston, Texas. Academic Press.NuevaYork. [ Links ]
MacArthur, R. H. & R. Levins. 1967. The limiting similarity convergence and divergence of coexisting species. Amer. Nat. 101: 377-385. [ Links ]
Mancilla, M. & M. Vargas. 1980. Los primeros estudios sobre la circulación y el flujo neto de aguas a través de la Laguna de Términos, Campeche, México. An. Centro Cienc. del Mar y Limnol. U.N.A.M. 7: 1-12. [ Links ]
Mann, K. H. 1980. Ecology of coastal waters: Studies in Ecology. Blackwell Scientific Publications. London.325 p. [ Links ]
Margalef, R. 1995. Teoría de los ecosistemas ecológicos. Universidad de Barcelona Pub. 289 p. [ Links ]
Orians, G. H.& G. Colliers. 1963. Competition and Blackbird social systems. Evolution. 17: 449-459. [ Links ]
Orlóci, L. 1978. Multivariate analysis in vegetation research. W. J. Junk . Netherlands. 189 p. [ Links ]
Peralta, M. D. 1984. Informe de actividades ostrícolas realizadas en las lagunas de Mechoacán, Carmen-Pajonal-Machona durante el bimestre marzo-abril de 1984. Secretaría de Pesca. Centro de producción acuícola de Puerto Ceiba, subprograma de producción ostrícola. Tabasco, México. 42p. [ Links ]
Pielou, E. C. 1984. The interpretation of ecological data: a premier on classification and ordination. Wiley, M. Nueva York. 396 p. [ Links ]
Psuty, N. P. 1966. The geomorphology of beach ridges in Tabasco, Mexico. Louisiana State Univ. Coastal Studies. Inst. Tech. Rept. 38 p. [ Links ]
Strickland, J. D. H. & T. R. Parsons. 1972. A practical handbook of sea water analysis. Fish. Res. Board Can. Bull. Ottawa. 167 p. [ Links ]
Suarez, C. J. & S. Gómez. 1965. 0bservaciones sobre el plancton de la Laguna de Términos, Campeche, México. Bull. Mar. Sci. 15: 1072-1120.
Swartz, R. C., F. A. Cole, D. W. Schultz & W. A. DeBen. 1986. Ecological changes in the Southern California Bight near a large sewage outfall: benthic conditions in 1980-1983. Mar. Ecol. Prog. Ser. 31: 1-13. [ Links ]
Villalobos, A. F. & M. E. Zamora. 1975. Importancia biológica de la Bahía de Campeche de la Península de Yucatán. Mem. II Symp. Latino. 0ceano. Biol. Venezuela. 47-56 p. [ Links ]
Wilbur, L. & J. Tiller. 1970. Man and environment change in South America. BioSci. 32: 1040-1042. [ Links ]
Woodin, S. A. 1981. Disturbance and community structure in a shallow water sand flat. Ecol. 62: 1052-1066. [ Links ]
Yáñez-Arancibia, A. & R. S. Nugent. 1978. El papel ecológico de los peces en estuarios y lagunas costeras, México. An. Centro Cienc. del Mar y Limnol. U.N.A.M. 4: 107-114. [ Links ]
Yáñez-Arancibia, A., A. Lara, P. Sánchez, I. Vargas, P. Chavance, F. Amezcua, A. Aguirre & S. Díaz. 1982. Ecosystem dynamics and nichthemeral and seasonal programing of fish community structure in a tropical estuarine inlet, Mexico. Proc. Inter. Symp. on Coastal Lagoons SCOR/IABO UNESCO. Italy. 417-429 p. [ Links ]
Yáñez-Arancibia, A. & P. Sánchez-Gil. 1988. Ecología de los recursos demersales marinos. AGT Editor, México.137 p. [ Links ]
Yáñez-Correa, A. 1963. Batimetría, salinidad, temperatura y distribución de los sedimentos recientes de la Laguna de Términos, Campeche, México. Bol. Inst. Geol. U.N.A.M. 67: 1-47. [ Links ]
1 Centro de Investigación y de Estudios Avanzados del IPN-Unidad Mérida. Km 6 Antigua a Progreso. A.P. 73 Cordemex, 97310 Mérida, Yucatán, México. Tel: (99) 81-2903 ext. 513, Fax: (99) 81-2917. E-mail: alitor@kin.ciamer.conacyt.mx


{kind=link}
{kind=link}