










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.48 n.4 San José Dec. 2000
José Rolando Bastida-Zavala y Sergio I. Salazar-Vallejo1
Recibido: 10-IX-00 Corregido: 9-III-00 Aceptado: 24-V-00
Abstract
From the revision of more than 1 250 serpulids, 15 species from the Grand Caribbean Region were identified and characterized. Thirteen species were collected along the shores of the Yucatan Peninsula and eight were found in other localities in the Gulf of Mexico, seven others are from Cuba and comments on type specimens of two species are also included. Three morphometric analyses were made to evaluate some characters. The first on Pomatostegus stellatus (Abildgaard), and related species and subspecies: P. brachysoma Schmarda, P. macrosoma Schmarda, P. s. fruticosa Mörch, P. s. pentapoma Mörch and P. s. tetrapoma Mörch, indicated that they are conspecific. Another one on Spirobranchus de Blainville species: S. dendropoma Mörch, S. giganteus (Pallas), S. polycerus (Schmarda) and S. polycerus augeneri ten Hove; indicated that S. giganteus differs from the other three taxa. The third analysis was made on Vermiliopsis annulata (Schmarda) sensu lato; it allowed the recognition of three different forms. Several incomplete specimens might belong to an undescribed genus. A key for identification of all the species recorded in the Grand Caribbean Region is also included
Key words
Caribbean, Keys, Morphometric Variation, Polychaetes, Serpulids, Taxonomy
Los serpúlidos son poliquetos comunes en fondos duros y, por lo general, son muy llamativos por la coloración de la corona radiolar; la porción que se expone fuera del tubo justifica que por su forma se les llamen pinitos de navidad. Casi todos se asocian a fondos duros, que pueden ser luego cubiertos por esponjas u otros organismos epibiontes, excepto por las especies del género Ditrupa Berkeley, 1835 ya que viven en fondos blandos sublitorales e incluso se han confundido con los moluscos escafópodos del género Dentalium (ten Hove & Smith 1990).
Se ha considerado que no hay mayor interacción entre los serpúlidos del género Spirobranchus y las especies de coral (ten Hove 1989), pero hay selectividad de las larvas de por lo menos un serpúlido por una especie de coral (Marsden 1987, Hunte et al. 1990a, 1990b, Marsden et al. 1990). En la Gran Barrera de Coral, se considera que la relación entre estos táxoneses mutualista (deVantier et al. 1986) porque el serpúlido dificulta la depredación sobre el coral por la estrella corona de espinas (Acanthaster planci). Esta relación estrecha de dos organismos sésiles ha permitido también el que pueda estimarse la edad de estos serpúlidos (Spirobranchus) a partir del número de bandas anuales que presenta el coral que lo envuelve; de esta manera se ha observado que pueden vivir desde diez hasta 40 años (Nishi y Nishihira 1996).
La presencia de un opérculo de desarrollo variable, la formación de un tubo calcáreo sinuoso y una membrana torácica, son los caracteres distintivos principales de la familia. Otro grupo de poliquetos muy afín es el de los espirórbinos, pero se distinguen por tener el cuerpo reducido, doblado en espiral, formar un tubo enrollado en forma dextrógira o levógira y por sus mecanismos especiales para la incubación de los embriones. Hay unos pocos serpúlidos capaces de incubar sus embriones; forman una cámara o expansión del tubo (Zibrowius 1983), incuban en los radiolos, o en el tubo (Nishi 1993). Un sabélido peculiar, Calcisabella Perkins, 1991 también forma tubo calcáreo pero su patrón corporal corresponde con los Sabellidae. Por una revisión reciente (Pillai & ten Hove 1994), se ha enfatizado la utilidad de algunas estructuras internas del tubo que son útiles en la separación de las especies de Spiraserpula Regenhardt, 1961.
La autoría de la familia no está estabilizada. McIntosh (1923) atribuye la autoría a Burmeister y Grube, pero por lo menos Grube (1851) incluyó a los Sabellidae. Fauchald (1977), ten Hove & Wolf (1984) y Fauchald & Rouse 1997) atribuyen la autoría de Serpulidae a Johnston; sin embargo, Johnston (1865), al igual que de Quatrefages (1865) y Grube (1878), incluyeron a los Sabellidae (+Eriographidae) dentro de la familia Serpulidae; Hartmann-Schröder (1996) la atribuyó a Savigny (1818), sin embargo, este año corresponde a la publicación de Lamarck y la de Savigny debe ser de 1820. En cualquier caso, sea Lamarck (1818) o Savigny (1820), la agrupación es heterogénea debido a que incluye a los Magile o a los Terebellidae y Hermellidae (=Sabellariidae), respectivamente. Perkins (1998) y ten Hove (com. pers. 1999) se la atribuyen a Rafinesque (1815) por ser el primero en considerar a los gusanos con tubo calcáreo como una categoría elevada (subfamilia Serpularia), aunque por su caracterización, "concha tubular calcárea" podría incluir a los moluscos vermétidos.
La primera evaluación crítica del grupo y su reconocimiento moderno como familia la realizó Mörch (1863), el único taxon situado erróneamente en Serpulidae es Phragmatopoma, considerado ahora en la familia Sabellariidae. Sin embargo, el primero en establecer la familia Serpularia sería Risso (1826), quien caracterizó a los Serpulidae por presentar un tubo calcáreo sólido con una apertura sencilla y branquias (= radiolos) separadas protegidas por un opérculo. Por esta razón, la familia debe atribuirse a este autor.
La clasificación tradicional de los serpúlidos descansa en dos subfamilias: Serpulinae Risso, 1826 y Filograninae Rioja, 1923; esta última fue propuesta en una revisión mayor (Rioja 1923). Las subfamilias se distinguen por tener el opérculo sobre un pedúnculo modificado en la primera, o por llevarlo en un radiolo en la segunda, aunque a veces puede faltar. Sin embargo, es posible que la pérdida de opérculo sea un carácter derivado al que se hubiera llegado por líneas independientes, por lo que esta distinción no es muy robusta (cf. Lechapt 1992, Pillai & ten Hove 1994). Una tercera subfamilia, Spirorbinae Mörch, 1863, fue propuesta para englobar a los serpúlidos con tubos enrollados y Pillai (1970) la elevó a categoría de familia. Pillai (1960) adicionó una cuarta subfamilia, Ficopomatinae Pillai, 1960, para las especies cuyo collar lleva setas denticuladas y habitan en aguas salobres.
Uchida (1978) propuso un esquema de 11 subfamilias que se separan por la presencia y características del opérculo, las setas del collar y los uncinos torácicos, principalmente. La clasificación propuesta, sin embargo, apareció en una revista poco accesible y no ha sido evaluada con detalle para aclarar los límites morfológicos entre géneros afines y entre las distintas subfamilias. Un carácter importante que merece ser considerado es el tipo de fotoreceptores ya que hay variaciones interesantes (ten Hove & Pantus 1985); por supuesto, el análisis de afinidades entre los géneros, para definir las subfamilias, debe ser basado en una evaluación estandarizada de la variación morfológica (ten Hove & Jansen-Jacobs 1984) y se está realizando actualmente (Bastida-Zavala & Monks, en prep.).
Los serpúlidos son una familia diversificada entre los poliquetos sedentarios del Gran Caribe, representados por 20 géneros, 65 especies y tres subespecies (Salazar-Vallejo 1996). En el litoral oriental de México se han registrado hasta el momento 19 especies, la mayoría de las cuales fueron citadas por Enrique Rioja en sus intensas investigaciones en la vertiente del Golfo, en donde incluso describió dos especies, Hydroides mongeslopezi Rioja, 1958 e H. mucronatus Rioja, 1958, pero lamentablemente sus colecciones parecían haberse perdido (Salazar-Vallejo 1989), aunque Vivianne Solís (ICML-UNAM) los recuperó.
En este trabajo se analizaron más de 1 250 ejemplares, de 41 localidades del litoral oriental de México y de 11 localidades de Cuba (Fig. 1 y 2, Apéndice 1). Resultaron 16 especies de serpúlinos, filográninos y ficopomátinos (se excluyen los espirórbinos). Se anexan breves comentarios del material tipo de dos especies más descritas de Bermuda y Puerto Rico (Fig. 2), los cuales están depositados en el American Museum of Natural History (AMNH).
Materiales y métodos
La clave de identificación para los géneros está basada en la propuesta de de León-González et al. (1989). Para la clave a especies se complementó con las de ten Hove (1970, 1975) para los géneros Spirobranchus y Pseudovermilia Bush, 1907, y la de ten Hove & Wolf (1984) para los serpúlidos del norte del golfo de México. El catálogo de Hartman (1959) permitió ubicar más fácilmente los nombres y sinonimias. Por su parte, los listados de especies regionales de Perkins & Savage (1975), de Salazar-Vallejo (1996) y de Perkins (1998) facilitaron la selección de referencias. La caracterización de las especies está basada en las características del ejemplar más grande, mejor conservado y/o completo. Las ilustraciones se dibujaron usando cámara clara y fotografías. Las claves de localidades utilizadas para el material examinado se explican en el Apéndice 1. El número entre paréntesis es el número de ejemplares examinado. En caso de incluirse material tipo se indica primero en material examinado y se incluyen los comentarios a dichos materiales al final de las observaciones.
El material recolectado se depositó en la colección de referencia de ECOSUR (Clave INE: QNR.IN.021.0497). Los ejemplares de Cuba están depositados en el Instituto Oceanológico de Cuba (IOC); otros materiales están en la colección de poliquetos del Instituto de Ciencias del Mar y Limnología (ICML-UNAM), de la Universidad Autónoma de Nuevo León (UANL) y de la Universidad Veracruzana (UV).
Consideraciones morfométricas: Existen varias propuestas de estandarizaciones para el estudio taxonómico de los serpúlidos (ten Hove & Jansen-Jacobs 1984, ten Hove & Pantus 1985), las cuales incluyen la medición de caracteres cuantitativos así como clasificación de caracteres cualitativos. En el presente estudio realizamos un análisis morfométrico de algunas especies que merecían atención especial. Como un resultado integrado de estos análisis por especie, se encontró que varios caracteres pueden ser equivalentes y estables. De esta manera, se consideró la longitud de la membrana torácica (medida ventralmente) como una variable independiente, cuya correlación con la longitud de pedúnculo opercular (opérculo incluido) es isométrica (Fig. 3a-b). Aunque persisten variaciones entre las diferentes especies, las tendencias observadas permiten considerar a ambos caracteres igualmente utilizables como variables independientes; sin embargo, recomendamos principalmente el uso de la longitud de la membrana torácica como el carácter principal, dado que el opérculo, por su mayor exposición a los depredadores, puede llegar a faltar.
La mayoría de los serpúlidos presentan siete setígeros torácicos, cuando el número sea diferente se indicará. Si aparecen datos merísticos y estadísticos, estos se abreviarán como n = tamaño de muestra, r: = intervalo de datos, µ = promedio, ± = desviación estándar, + = más de. Otras abreviaturas son latinas: ca. = circa (cerca), cf. = confer (compárese con), s.l. = sensu lato (en el sentido amplio), s.str. = sensu stricto (en el sentido estricto). Todas las escalas de las figuras están en milímetros.
Resultados y discusión
Serpulidae Risso, 1826
Filograninae Rioja, 1923
Salmacina Claparède, 1870
Salmacina incrustans Claparède, 1870
(Fig. 4a-h)
Referencia: Rioja, 1931: 440-441, Lám. 144, Fig. 9.
Material examinado: +570 ejemplares. CON1(+100), CON3(+100), CON6(35), COZ1(62), ECH1(8), ECH3(15), EL2780(1), ESP14(1), LAG1(+100), NC3(30), NIZ3(20), SAM1(72), SF1(3), SJU1(19).
Caracterización: Animal gregario. Tubo blanco con anulaciones transversales. Animal diminuto, cuerpo amarillo. Sin opérculo. Tórax con ocho segmentos (r: 7-9). Setas del collar con hombro compuesto por 5-6 (r: 4-7) dientes grandes (Fig. 4a-b). El resto de los setígeros torácicos con setas "Apomatus" (Fig. 4c) y capilares gruesas (Fig. 4d). Uncinos torácicos subcuadrados con 8-10 dientes aguzados, el primero es más grande (Fig. 4e). Abdomen con setas geniculadas (Fig. 4f). Uncinos con varias hileras de dientes, 6-8 dientes aguzados vistos de perfil (Fig. 4g-h).
Distribución y hábitat: Océano Atlántico y Mediterráneo. En pilotes de madera con los tubos cubiertos por briozoos, en fondos mixtos, epibionte de erizos, fondos rocosos con algas, sobre corales, epibionte de ascidias en una pared de vermétidos y ostras.
Observaciones: Las diferencias con S. dysteri (Huxley, 1855), especie aparentemente cosmopolita, radican en la cantidad de dientes de las setas del collar (Rioja 1931); sin embargo, se sospecha que muchas determinaciones son erróneas (ten Hove & Wolf 1984). Rioja (1946) registró S. dysteri para el Golfo de México, pero describe las setas del collar con 6-8 dientes gruesos, similares a los de S. incrustans; posteriormente, Rioja (1959) consideró que se trata de una subespecie: S. dysteri incrustans. Hay controversia sobre el estatus del género, debido a que Salmacina sólo se separa de Filograna por carecer de opérculo. Zibrowius (1973: 80) consideró que todas las formas de ambos géneros se mantengan como F. implexa Berkeley, 1827; sin embargo, un detallado estudio morfológico y de distribución (Gee 1963) confirmó que las formas operculadas y anoperculadas son especies diferentes, que incluso ocupan zonas marinas diferentes (Salmacina exclusivamente de la zona litoral y Filograna de la sublitoral). Gee (1963) mantiene ambas formas como especies diferentes dentro de Filograna, aunque su propuesta no es robusta considerando que la presencia o ausencia del opérculo es un carácter suficiente para separarlos a género, reforzado por su segregación ecológica.
Por su parte, un estudio sobre la reproducción asexual de ambos géneros (Vannini 1975), indicó que hay diferencias clonales dentro del grupo y muestran una correlación entre el número de segmentos torácicos de progenitores y descendientes. Ten Hove & Wolf (1984) sugieren que deben revisarse detalladamente las especies de estos géneros utilizando microscopía electrónica (SEM) para comprobar la validez de las diferencias pequeñas, pero constantes, en las setas del collar y quizás también de los uncinos. De esta forma, se comprobará si hay constancia en el número de dientes en las setas del collar; pocos dientes en S. incrustans o innumerables en S. dysteri. Otra especie, S. amphidentataJones, 1962, descrita de Jamaica, presenta setas modificadas del collar de tres tamaños diferentes (ver nuestra clave).
Ficopomatinae Pillai, 1960
Ficopomatus Southern, 1921
Ficopomatus miamensis (Treadwell, 1934)
(Fig. 4i-s)
Referencia: ten Hove & Weerdenburg, 1978: 106-109, Fig. 1f-i, 3c, 5a-c.
Material examinado: +130 ejemplares. CHE1(+100), CHE2(22), CHE3(7), LAR1(2).
Caracterización: Animal gregario. Tubo blanco con anulaciones transversales. Animal diminuto, cuerpo amarillo. Opérculo esférico, blando y amarillo (Fig. 4i); en otros ejemplares tiene forma de higo o de disco ligeramente convexo (Fig. 4j). Setas del collar de dos tipos, con serraciones agudas y parte distal finamente serrada (Fig. 4k) y capilares (Fig. 4l). El resto de los setígeros presenta setas capilares (Fig. 4m) y limbadas (Fig. 4n). Uncinos torácicos triangulares, con siete dientes aguzados en una hilera, el primero más grande (Fig. 4o-p). Abdomen con setas geniculadas (Fig. 4q). Uncinos similares a los torácicos pero con más dientes, entre 8-9 en una hilera, el primero ligeramente más grande (Fig. 4r-s).
Distribución y hábitat: Anfiamericana. Gran Caribe y Pacífico de Panamá. Lagunas costeras y canales de manglar, en aguas salobres.
Observaciones: Pillai (1960) erigió la subfamilia Ficopomatinae para agrupar a las especies con setas del collar fuertemente denticuladas y que aparentemente se encuentran restringidas a aguas salobres. Sin embargo, ten Hove & Weerdenburg (1978) realizaron una revisión de Ficopomatus y sinonimizaron Mercierella Fauvel, 1923, Mercierellopsis Rioja, 1945, Sphaeropomatus Treadwell, 1934 y Neopomatus Pillai, 1960 con F. miamensis. Consideraron que las diferencias en la forma y ornamentación de los opérculos, junto con la alta variabilidad encontrada en las setas del collar de las distintas especies, no ameritaba la separación genérica y por lo tanto también rechazaron el estatus de la subfamilia. Empero, la forma de las setas del collar es característica de este género, como ellos mismos reconocieron (ten Hove & Weerdenburg 1978: 98), lo que aunado a la fusión de la membrana torácica permite retener la subfamilia.
Serpulinae Risso, 1826
Pomatoceros Philippi, 1844
Pomatoceros cf. minutusRioja, 1941
(Fig. 5a-f)
Referencia: Zibrowius 1970: 15-16, Lám. 3, Fig. 11-15.
Material examinado: PO75004(1), CAZ1(+100).
Caracterización: Tubos blancos, muy frágiles, con costillas longitudinales y alvéolos entre las costillas (Fig. 5a). Animal diminuto, cuerpo amarillo. Pedúnculo opercular con alas cortas y delgadas a cada lado. El opérculo es un disco blanco y blando con dos tubérculos diminutos (Fig. 5b). Setas torácicas limbadas (Fig. 5c), incluyendo las del collar. Uncinos torácicos rectangulares, con 9-10 dientes agudos, el primero más grande y obtuso (Fig. 5d). Abdomen con setas "trompeta" (Fig. 5e). Uncinos similares a los torácicos, con 8-10 dientes aguzados (Fig. 5f).
Distribución y hábitat: Anfiamericana? Pacífico oriental, desde México hasta Perú, Golfo de México, Brasil. Sobre algas carnosas.
Observaciones: Estos ejemplares parecen pertenecer a P. minutus, fue descrita del Pacífico Oriental tropical y documentada en Brasil (Zibrowius 1970). Las pequeñas diferencias en el opérculo y pigmentación del tubo ameritan ser estudiadas con más detalle y comparando directamente con ejemplares del Pacífico Oriental tropical.
Pomatostegus Schmarda, 1861
Pomatostegus stellatus (Abildgaard, 1789)
(Fig. 3b, 5g-q, 6a-d)
Referencia: Rioja 1959: 293-296, Fig. 92-100.
Material examinado: 134 ejemplares. Material tipo (AMNH 1266, como Protis torquata), CHK1(6), CON2(1), CON6(3), COZ1(44), COZ2b(3), COZ3(1), CU7(2), EL2780(1), IP1(2), LAG1(8), NC3(14), NIZ1(10), NIZ2(8), SF1(2), XAH1(8), XAH2(17), XCA3(2), XCAY1(1).
Caracterización: Tubo blanco, con una cresta dorsal lobulada, en el tercio distal forma una lámina delgada que termina en una espina horizontal saliente a partir de la abertura del tubo. A cada lado del tubo y en su parte basal hay dos pares de crestas más, las cuales presentan lóbulos y espinas (Fig. 5g). Cuerpo pardo amarillento, de 45 mm (n=39, r: 6.2-78, µ=37.2 ±14.6). Tórax con 8 mm de largo (n=57, r: 1.8-12, µ=6.8 ±2.3) y 2.8 mm de ancho (n=57, r: 0.7-5.5, µ=3.5 ±1.1). Abdomen con tres segmentos asetígeros y 88 setígeros (n=44, r: 43-125, µ=93.4 ±14.6). Radiolos con 2-3 bandas de pigmento negro-azul iridiscente. Pedúnculo opercular maculado en más de dos tercios de su superficie, con 8 mm de largo incluyendo la copa opercular (n=50, r: 1.5-18, µ=8.4 ±3), y 3.2 mm de ancho en la placa basal (n=48, r: 1.1-5, µ=2.9 ±0.8). Copa opercular quitinosa, de 3 mm de longitud (n=48, r: 0.5-6, µ=2.5 ±1.1); con cuatro discos (n=48, r: 2-6, µ=4 ±1), el primero está fusionado a la placa opercular. Discos festoneados y concéntricos, unidos por un tubo con bordes espinosos, el remate de dicho tubo y las espinas forman una estrella distal, la cual es similar a la observada bajo cada uno de los discos (Fig. 5h). Setas del collar de dos tipos, "bayoneta" con hombro con incontables dientes minúsculos y limbo finamente serrado (Fig. 5i-j), el resto son capilares con denticulación muy fina desde la parte media (Fig. 5k-l). Los demás setígeros torácicos con setas limbadas (Fig. 5m) y setas "Apomatus" (parte media limbada y tercio distal finamente denticulado) (Fig. 5n). Uncinos rectangulares, con 12-16 dientes, el diente anterior es romo y más grande, el resto son aguzados (Fig. 5o). Setas abdominales geniculadas (Fig. 5p). Uncinos similares a los torácicos pero de menor tamaño, con 6-10 dientes (Fig. 5q).
Distribución y hábitat: Gran Caribe, desde Florida a Brasil. En el Golfo de México sólo está registrado para Cayo Alacranes, y en el litoral continental de Quintana Roo hasta San Felipe (Yucatán). Se le encuentra adherido a rocas y asociado a corales de fuego y escleractíneos.
Observaciones: Schmarda (1861) definió dos especies por la forma y el tamaño del cuerpo: P. brachysoma y P. macrosoma. Por su parte, Mörch (1863) estableció tres subespecies de P. stellatus con base en el número de discos operculares: P. stellatus fruticosa con cuatro discos, P. s. pentapoma con cinco discos, P. s. tetrapoma con cuatro discos. Dichas especies o subespecies no pueden mantenerse considerando la evidencia proporcionada por un análisis de 57 ejemplares de P. stellatus s.l. provenientes del litoral oriental de México y de Cuba (Fig. 6). Las especies descritas por Schmarda (P. brachysoma y P. macrosoma), no se separan por tamaño; por ejemplo, al relacionar el diámetro del primer disco opercular con la longitud del tórax (Fig. 6a), resulta evidente la relación isométrica de dicho carácter.
Sobre las subespecies de Mörch (1863) no existe relación entre el número de discos y la longitud del tórax (Fig. 6b). Los opérculos presentan una serie de discos cuyo número es independiente de la longitud de la membrana torácica. Pueden presentar desde dos hasta seis discos (n=48, µ=4 ±1) y, al parecer, esta diferencia puede deberse más a la acción eventual de los depredadores. Debido a su inestabilidad, el número de discos no puede ser utilizado para separar especies o subespecies. La relación entre la longitud del pedúnculo opercular y la longitud del tórax es isométrica (Fig. 3b), y entre los ejemplares analizados con opérculo (n=48), se encontró que éste puede estar inserto en cualquier lado (24 en el derecho, 26 en el izquierdo). Al analizar los pedúnculos de un solo lado, resulta menos isométrica la relación de la longitud del tórax con los opérculos izquierdos (Fig. 6c) que con los derechos (Fig. 6d). Esta evidencia indica que los opérculos derechos emergen primero, mientras que los izquierdos resultan de una pérdida previa del opérculo anterior.
Con la evidencia presentada, se debe usar sólo P. stellatus para las formas del Gran Caribe. Sin embargo, P. kroeyeri Mörch, 1863 del Pacífico centroamericano y P. actinoceras Mörch, 1863 de Filipinas deben retenerse por su localización.
Observaciones sobre el material tipo: Holotipo de Protis torquata Hoagland, 1919, que consiste de dos porciones de tubo (una costilla central longitudinal y dos accesorios menos marcados) y un ejemplar muy manipulado, sin opérculo, con 12.5 mm de largo incluyendo los radiolos (de 2.5 mm) y 1.5 mm de ancho. Presenta seis setígeros torácicos. Zibrowius anotó (13 de junio de 1968) "holotipo de P. torquata (.) este ejemplar no pertenece al género Protis sino al género Pomatostegus. Parece un juvenil de P. stellatus que es la única especie del Atlántico occidental tropical". Recientemente Zibrowius (com. pers.) confirmó que se trata de un juvenil de P. stellatus.
Protula Risso, 1826
Protula diomedeaeBenedict, 1887
(Fig. 7a-g)
Referencia: Benedict 1887: 547-548, Lám. 20, Fig. 1-6.
Material examinado: CU9(1).
Caracterización: Fragmento del tubo áspero, la superficie presenta tubérculos. Cuerpo amarillo de 17 mm. Corona con 22 radiolos por lado. El radiolo más largo mide 4 mm. Los primeros seis radiolos dorsales presentan procesos basales externos a nivel de la membrana radiolar (Fig. 7a). Tórax de 6 mm de largo y 2.5 mm de ancho, la membrana torácica llega hasta el séptimo setígero. Setas del collar capilares, el resto de los setígeros torácicos tienen setas limbadas (Fig. 7b-d). Uncinos subcuadrados con numerosos dientes, el primero de ellos forma una espina prominente (Fig. 7e). Parte anterior del abdomen con setas geniculadas (Fig. 7f) y la posterior con setas capilares. Uncinos similares a los torácicos, ligeramente más pequeños (Fig. 7g).
Distribución y hábitat: Costa oriental de Estados Unidos y Cuba. Fondo con ceibadal y fango.
Observaciones: Desde su descripción original no se había vuelto a encontrar la especie. Las diferencias entre las especies del género radican en las ornamentaciones del tubo y el número de radiolos. El patrón ventral de los vasos sanguíneos en el tórax ha servido para separar algunas especies europeas del mismo género tanto en ejemplares in vivo como fijados (Hanson 1948: 581, ten Hove & Pantus 1985); sin embargo, en el ejemplar estudiado no se observan los vasos sanguíneos, quizás debido al tiempo de preservación. Esta especie parece restringirse a aguas subtropicales del Gran Caribe.
Protula longiseta Schmarda, 1861
(Fig. 7h-o)
Referencia: Augener 1925: 39-42, redescripción sin Fig.
Material examinado: 2 ejemplares. NIZ1(1), XAH1(1).
Caracterización: Tubos perdidos. Cuerpo amarillo de 22 mm. Corona con 16 radiolos por lado. Tórax con 5 mm de largo y 2.3 mm de ancho, la membrana torácica llega hasta el cuarto setígero y está muy expandida. Setas del collar limbadas (Fig. 7h) y capilares (Fig. 7i). El resto de los setígeros torácicos tienen setas limbadas (Fig. 7j-k). Uncinos con 10-12 dientes aguzados, el primero de ellos más largo y obtuso (Fig. 7l). Abdomen con setas geniculadas (Fig. 7m). Uncinos más pequeños que los torácicos, con varias hileras de dientes, con 10-12 dientes vistos de perfil (Fig. 7n-o). Glándula pigidial muy pigmentada.
Distribución y hábitat: Mar Caribe. Playas mixtas o rocosas, en talos de algas.
Observaciones: Desde la redescripción de Augener (1925) no se había vuelto a registrar la especie. Augener revisó el ejemplar tipo de Schmarda y consideró a P. antennata Ehlers, 1887 como sinónimo de P. longiseta (ver nuestra clave). También sugiere que P. alba Benedict, 1887 es idéntica a P. longiseta pero como la descripción de Benedict es muy breve, deberá revisarse el material tipo de P. alba. Augener (1925, p: 40) también observó que la membrana torácica llega hasta el cuarto setígero: "Das letztere erreicht seine grösste Höhe im Bereich der 4 ersten Thoraxsegments und fällt dann nach hinten schnell an Höhe ab".
Pseudovermilia Bush, 1907
Pseudovermilia fuscostriata ten Hove, 1975
(Fig. 8a-k)
Referencia: ten Hove 1975: 79-85, Fig. 128-134, 146, 149, 151, 160, 165, 175, Lám. 5, 8a-b.
Material examinado: 5 ejemplares. AVE4(1), EL2771(2), EL2779(1), EL2792(1).
Caracterización: Tubo blanco con bandas pardas transversales, una quilla longitudinal dorsal con una ondulación anterior, con varias hileras de alvéolos y espinas (Fig. 8a-b). Cuerpo amarillo. Opérculo amarillo, hialino, multianulado, con una espina distal ancha (Fig. 8c). Otro ejemplar presenta una espina distal ramificada (Fig. 8d). Setas del collar capilares (Fig. 8e) y limbadas finamente serradas (Fig. 8f). El resto de los setígeros torácicos presentan setas "Apomatus" (Fig. 8g) y limbadas (Fig. 8h). Uncinos con 12-14 dientes aguzados en una hilera, el primero bifurcado (Fig. 8i). Abdomen con setas geniculadas anteriores (Fig. 8j) y capilares en los segmentos posteriores. Uncinos más pequeños que los torácicos, con varias hileras de dientes, diez dientes aguzados vistos de perfil, el primero más ancho y bifurcado (Fig. 8k).
Distribución y hábitat: Gran Caribe. Florida al sur de Brasil. En formaciones coralinas, sobre conchas de cangrejos ermitaños y hasta más de 100 m.
Pseudovermilia multispinosa
(Monro, 1933)
(Fig. 8l-q)
Referencia: ten Hove 1975: 72-79, Fig. 124-127, 142-143, 147-148, 152, 157, 166, 173-174, Lám. 2f, 4, 8f-g.
Material examinado: 3 ejemplares. CU8(2), EL2771(1).
Caracterización: Tubo blanco con una quilla longitudinal dorsal y varias hileras de alvéolos. Cuerpo amarillo de 12 mm (r: 7-12). Pedúnculo opercular de 4.5 mm de largo (r: 3-6), opérculo con una base amarilla opaca, blanda y una parte distal negra y dura, con una espina arborescente central (Fig. 8m). Otro ejemplar presenta una gran espina central ramificada rodeada por múltiples espinas periféricas muy delgadas y ramificadas (Fig. 8l). Tórax de 2.1 mm de largo (r: 1.9-3) y 1 mm de ancho (r: 0.9-1). Setas del collar capilares finamente serradas (Fig. 8n). El resto de los setígeros torácicos presentan setas "Apomatus" (Fig. 8o), capilares (Fig. 8p) y limbadas (Fig. 8q). Uncinos con 15 dientes aguzados en una hilera, el primero bifurcado. Abdomen con setas geniculadas anteriores y capilares en los segmentos posteriores. Uncinos con varias hileras de dientes, el primero más ancho y bifurcado.
Distribución y hábitat: Anfiamericana. Gran Caribe, desde Florida hasta el sur de Brasil. Pacífico de Panamá (isla Gorgona). En formaciones coralinas.
Pseudovermilia occidentalis
(McIntosh, 1885)
(Fig. 8r-u)
Referencia: ten Hove 1975: 59-72, Fig. 114-123, 144-145, 155-156, 159, 161-164, 170-172, Lám. 1, 2a-e, 3, 7
Material examinado: 198 ejemplares. AVE4(11), CHA13-14(22), CON3(35), COZ1(1), CU5(2), EL2777(1), ESP13(2), LAG1(68), MAJ1(1), NC2(1), NC3(45), NIZ3(3), PO75005a(1), PO75005b(2), SC(1), XCA3(1), XCAY1(1).
Caracterización: Tubo blanco con surcos transversales y una quilla longitudinal dorsal. Cuerpo amarillo. Opérculo con una base amarilla opaca, blanda y una parte distal negra y dura con una espina curva simple (Fig. 8r). Otros ejemplares no presentan la espina distal (Fig. 8s), presentan una parte distal muy prolongada (Fig. 8t), o una espina ganchuda (Fig. 8u). Setas del collar capilares finamente aserradas. El resto de los setígeros torácicos presentan setas limbadas y a partir del tercer setígero setas "Apomatus". Uncinos con 13 dientes aguzados en una hilera, el primero bifurcado. Abdomen con setas geniculadas anteriores y capilares en los segmentos posteriores. Uncinos con varias hileras de dientes, el primero más ancho y bifurcado.
Distribución y hábitat: Anfiamericana. Pacífico oriental, desde California hasta Galápagos, Islas Hawaii. Ambas costas tropicales y subtropicales del Atlántico, Islas de Cabo Verde y Santa Helena. En rocas calcáreas y coralinas, sobre fondos mixtos o en esponjas (Clathria calla).
Observaciones: Ten Hove & Smith (1990: 110) considera que la forma del Indopacífico es diferente a la forma nominal. La especie amerita una revisión y comparar ejemplares de las diferentes localidades de ambas costas de América. Las formas del Gran Caribe son P. occidentalis, pues la localidad tipo es Bermuda.
Sclerostyla ctenactis
Mörch, 1863
Referencia: ten Hove, 1973: 6-12, Fig. 1-4, 7, 20-31, Lám. 1, 3a-b, 3, 7.
Material examinado: Holotipo (AMNH 1996) de Placostegus calciferus Treadwell, 1929: 12-13. Julia Cove, Bahía Guanica, Puerto Rico, 23 VI 1915.
Características: Al ejemplar le falta el opérculo. Mide 19 mm de largo y 2.5 mm de ancho. El frasco contiene un eunícido (ca. E. cariboea) al que le falta el aparato mandibular.
Spirobranchus de Blainville, 1818
Spirobranchus dendropoma Mörch, 1863
(Fig. 9a-i, 11a-b)
Referencia: ten Hove 1970: 3-14, Fig. 3, 4-14, 27, 30-32, Lám. 1, 5C, F-G.
Material examinado: 3 ejemplares. IE1(1), IE2(2).
Caracterización: Tubo perdidos. Cuerpo amarillo. Corona formada por dos semicírculos de radiolos pálidos, aunque en los otros dos ejemplares presentan bandas rojizas. En un ejemplar (IE2) se presentan procesos interradiolares redondeados (Fig. 9c). Pedúnculo opercular amarillo. Alas pequeñas, distalmente crenuladas, ocupan un cuarto de su longitud (Fig. 9a). En otros ejemplares las crenulaciones están presentes pero tienen distinto grado de desarrollo (Fig. 9b). Opérculo con tres grupos de espinas ramificadas; dos latero-dorsales, con una y dos ramas principales (con varias espinas secundarias pequeñas); y un grupo medio-ventral, con dos ramas (con varias espinas secundarias pequeñas) (Fig. 9a). Los otros ejemplares son diferentes, con sólo dos espinas no ramificadas y una protuberancia en el lugar donde se encontraría el tercer grupo de espinas (Fig. 9b). Setas del collar típicas del género ("Spirobranchus"), con un hombro piloso y un limbo aserrado (Fig. 9d). Los demás setígeros torácicos presentan setas limbadas (Fig. 9d-f). Uncinos rectangulares con 13-14 dientes, el primero bifurcado (Fig. 9g). Abdomen con 102 setígeros. Setas abdominales "trompeta" (Fig. 9h). Uncinos similares a los torácicos pero de menor tamaño, con 12-14 dientes (Fig. 9i).
Distribución y hábitat: Gran Caribe. Sobre coral muerto o vivo (Acropora).
Observaciones: S. dendropoma fue sinonimizado por ten Hove (1970) con S. tetraceros (Schmarda, 1861); dado que la segunda es una especie australiana es conveniente conservar el nombre usado para las formas del Caribe. Asimismo, la distinción puede facilitarse con los procesos bilobulados en la base de los radiolos que ten Hove (1970) y Liñero-Arana (1999) citan para ejemplares de Bonaire y Venezuela, respectivamente; sin embargo, los procesos lobulados se presentan sólo en uno de los ejemplares revisados en este trabajo. En un análisis los tres ejemplares de esta especie no se separan de S. polycerus ni de S. p. augeneri, y la tendencia que siguen es similar a S. polycerus (Fig. 11a-b). Véanse los comentarios en S. giganteus.
Spirobranchus giganteus (Pallas, 1766)
(Fig. 3b, 9j-p, 11a-b)
Referencia: ten Hove 1970: 14-22, 49, Fig. 35-62, Lám. 2A-B, 5A, D.
Material examinado: 26 ejemplares. COZ2b(6), COZ3(13), CU10(1), IP1(1), NIZ2(5).
Caracterización: Tubo blanco y liso, con una quilla dorsal muy suave que se prolonga, distalmente desde la boca del tubo, en una espina morada dirigida hacia delante o con una ligera inflexión hacia arriba. También presenta dos costillas basales, una a cada lado del tubo, que terminan en una espina dirigida hacia delante. En corte transversal el tubo tiene forma triangular. El cuerpo es amarillo ocre de 73.5 mm (n=11, r: 38.5-75, µ=60.6 ±12.3). Corona con dos espirales de radiolos morados, aunque en los demás organismos el color es variable. Pedúnculo opercular pálido, con 19 mm de largo (n=15, r: 12-19, µ=15.9 ±1.9) y 6.5 mm de ancho en la placa basal calcárea (n=15, r: 5-8, µ=6.3 ±0.7). Alas distalmente enteras, ocupan dos tercios de su longitud. Opérculo formado por tres espinas ramificadas, proyectadas diagonalmente en dirección ventral. Dos son latero-dorsales, con una espina secundaria dorsal y varias espínulas secundarias pequeñas. La tercera espina es medio-ventral, distalmente bifurcada (Fig. 9j); en otros ejemplares esta espina puede faltar o tener diferentes grados de desarrollo. Tórax con 19 mm de largo (n=15, r: 6.5-19, µ=13.5 ±3.3) y 6 mm de ancho (n=15, r: 5-12, µ=6.5 ±1.8). Setas del collar "Spirobranchus" (Fig. 9k-l); en otros ejemplares el haz de setas es rebasado por el tejido del collar, aparentando no presentar setas del collar, salvo por un minúsculo orificio. Los demás setígeros torácicos con setas limbadas (Fig. 9m). Uncinos rectangulares, con 20-24 dientes, el primero obtuso (Fig. 9n). Presenta 225 setígeros abdominales (n=9, r: 137-225, µ=180.2 ±25.5). Setas abdominales "trompeta" (Fig. 9o). Uncinos similares a los torácicos, pero de menor tamaño, con 12-14 dientes, el primero es obtuso (Fig. 9p).
Distribución y hábitat: Gran Caribe. Dentro y sobre corales escleractíneos y corales de fuego.
Observaciones: Esta especie ha suscitado confusión por su variabilidad morfológica (ten Hove 1970, Fiege & ten Hove 1999). Por ello, se realizó un análisis que incluye a las otras especies del género. Se tomaron medidas a 28 ejemplares: 3 S. dendropoma, 15 S. giganteus, 3 S. polycerus y 1 S. p. augeneri. En todas las relaciones morfométricas se separa claramente S. giganteus del resto (Fig. 11a-b). Sin embargo, no ocurre lo mismo entre las otras especies, aunque esto se debe quizá a la cantidad de ejemplares disponibles. Los caracteres de S. giganteus presentan estabilización con respecto a la longitud del tórax: altura perpendicular de las espinas principales, ancho del tórax, longitud del pedúnculo opercular (Fig. 3b), ancho del pedúnculo opercular (Fig. 11a), y número de segmentos abdominales (Fig. 11b). En cambio, la longitud del radiolo no parece tener relación con la del tórax, lo cual puede deberse a que es una estructura blanda expuesta a la depredación (e.g. mordisqueo de peces). Los datos aparecen dispersos, indicando tal vez una variada tasa de regeneración en estas estructuras.
Spirobranchus polycerus (Schmarda, 1861)
(Fig. 10a-g, 11a-b)
Referencias: ten Hove 1970: 32-38, Fig. 74-102, 122-123, Lám. 3B, 4A-B, D, 5B; Marsden, 1992: 14-18.
Material examinado: 34 ejemplares. AVE4(1), COZ2b(1), COZ3(31), XAM1(1).
Caracterización: Tubo blanco y liso, con una quilla dorsal muy suave que se prolonga distalmente, desde la boca del tubo, en una espina dirigida hacia delante. El tubo también presenta dos costillas basales, cada una a los lados del tubo, que terminan en una espina dirigida hacia delante. En corte transversal el tubo tiene forma triangular. Cuerpo verde olivo de 40 mm (n=9, r: 18-40, µ=33 ±7.5). Corona formada por dos espirales de radiolos verde-amarillento. Pedúnculo opercular pálido, con 5 mm de largo (n=9, r: 3.5-7, µ=4.9 ±7.5) y 3 mm de ancho en la placa basal calcárea (n=9, r: 1.5-3, µ=2.4 ±0.5). Alas enteras, ocupan dos tercios de su longitud. Opérculo con siete espinas trifurcadas en arreglo circular y algunas espinas secundarias (Fig. 10a-b); hay algunos con espinas bifurcadas o incluso con espinas en regeneración. Tórax con 7 mm de largo (n=9, r: 4-8, µ=6.2 ±1.3) y 3 mm de ancho (n=9, r: 1.9-3.5, µ=2.7 ±0.5). Setas del collar "Spirobranchus" (Fig. 10c), presentándose en haces muy numerosos. El resto de los setígeros torácicos presentan setas limbadas (Fig. 10d) en haces numerosos. Uncinos subtriangulares, con 12-15 dientes aguzados, el primero ligeramente mayor que los demás (Fig. 10e). Abdomen con 109 setígeros abdominales (n=9, r: 85-120, µ=107 ±11.6). Setas abdominales "trompeta" (Fig. 10f). Uncinos similares a los torácicos pero de menor tamaño, con 11-13 dientes aguzados, el primero es ligeramente mayor y obtuso (Fig. 10g).
Distribución y hábitat: Mar Caribe. Sobre roca coralina y corales de fuego.
Observaciones: El análisis indica que esta especie se separa de S. giganteus, tanto en la relación longitud del tórax/ancho del pedúnculo opercular como en la de longitud del tórax/número de segmentos abdominales (Fig. 11a-b). Véanse los comentarios en S. giganteus.
Spirobranchus polycerus augeneri
ten Hove, 1970
(Fig. 10h-o, 11a)
Referencias: ten Hove 1970: 38-42, Fig. 109-118, 119-121, Lám. 3A-B, 4C; Marsden, 1992: 14-18.
Material examinado: COZ2b(1).
Caracterización: Tubo blanco, inmerso en roca coralina. Ejemplar incompleto, cuerpo amarillo. Corona con dos semicírculos de radiolos pálidos. Pedúnculo opercular pálido, con 4.8 mm de largo y 1.8 mm de ancho en la placa basal calcárea. Alas distalmente enteras, ocupan un tercio de su longitud. Opérculo con tres espinas ramificadas, dos triangulares en posición latero-dorsal y la tercera más pequeña, en posición medio-dorsal (Fig. 10h-i). Tórax con 4.6 mm de largo y 1.8 mm de ancho. Setas del collar limbadas, con serraciones a lo largo del limbo (Fig. 10j) y además setas "Spirobranchus", con hombro piloso y un limbo finamente serrado (Fig. 10k). Los demás setígeros torácicos presentan setas limbadas (Fig. 10l). Uncinos rectangulares, con 15 dientes, el primero obtuso y ligeramente mayor que los demás (Fig. 10m). Setas abdominales "trompeta" (Fig. 10n). Uncinos similares a los torácicos pero de menor tamaño, con 10-13 dientes (Fig. 10o).
Distribución y hábitat: Mar Caribe. Sobre roca coralina.
Observaciones: En un análisis con datos morfométricos, el único ejemplar de esta especie no se separa de la especie parental, S. polycerus, ni de S. dendropoma (Fig. 11a). Sin embargo, Marsden (1992) mostró que dos formas asignables a Spirobranchus polycerus (Schmarda, 1861) están aisladas reproductivamente y deben reconocerse como especies independientes (ver clave). Las formas son reconocidas como subespecies por ten Hove (1970): S. p. augeneri con dos cuernos y S. p. polycerus con siete cuernos. Aunque hay formas intermedias, el porcentaje es de alrededor del 5% (Marsden 1992). Por estas diferencias y con ejemplares en buen estado, se debe elevar esta subespecie como una especie independiente.
Vermiliopsis Saint-Joseph, 1894
Vermiliopsis annulata s.l. (Schmarda, 1861)
(Fig. 3a, 12a-h, 13a-b)
Referencias:ten Hove & Wolf 1984: 55.33-34, Fig. 55.28; ten Hove & San Martín 1995: 21-22.
Material examinado de Vermiliopsis: 49 ejemplares. Holotipo (AMNH 2396) de Vermilia glandulata Treadwell, 1936: 36-39, 65. Gurnets Rock, Bermuda, 35 pies, 19 VIII 1931. =V. bermudensis (Bush, 1904) fide Hartman (1956: 259), sin análisis ni discusión. Las formas pueden caracterizarse por la forma del opérculo.
V. annulata embudo
(Fig. 12a-o, 13a-b)
Material examinado: 11 ejemplares. CHA13-14(1), CON3(1), COZ1(1), CU11(2), F5(3), LAG1(1), NC2(1), NIZ1(1).
Caracterización: Tubo blanco, enrollado, con anulaciones tenues y boca muy amplia (Fig. 12a), otros tubos tienen cinco quillas longitudinales (Fig. 12b) y otros peristomas. Cuerpo amarillo de 21 mm (n=7, r: 6-24, µ=14.2 ±7.7). Pedúnculo opercular de 6 mm (n=10, r: 1.6-6, µ=3.5 ±1.4) y copa opercular de 2.6 mm (n=10, r: 0.4-2.7, µ=1.7 ±0.8), con forma de embudo (Fig. 12c), pardo oscuro, con siete discos fusionados (n=10, r: 2-12, µ=6.2 ±3.2), aunque existe cierta variación en los otros ejemplares (Fig. 12d-f), incluso con una proyección distal (Fig. 12g). Tórax de 6.2 mm de largo (n=9, r: 1.5-6.2, µ=3.2 ±1.7) y 2.2 mm de ancho (n=9, r: 0.5-2.2, µ=1.3 ±0.7). Membrana torácica llega hasta el quinto setígero (n=10, r: 4-6). Setas del collar capilares (Fig. 12h) y limbadas muy delgadas (Fig. 12i); el resto de los setígeros torácicos tienen setas limbadas (Fig. 12j) y "Apomatus" (Fig. 12k-l). Uncinos torácicos rectangulares con una hilera de 12-14 dientes aguzados, el primero ligeramente más grande (Fig. 12m). Abdomen con setas geniculadas (Fig. 12n), primeros segmentos abdominales asetígeros. Uncinos similares a los torácicos pero más pequeños, con una hilera de 10-12 dientes (Fig. 12o).
V. annulata Sarakof
(Fig. 12p-s, 13a-b)
Material examinado: 4 ejemplares. BUE1(1), COZ1(1), NC3(1), XAH2(1).
Caracterización: Tubo blanco con cinco quillas longitudinales apenas reconocibles (Fig. 12p) y con peristomas. Cuerpo amarillo de 15 mm (n=3, r: 7.2-15, µ=10.1 ±4.3). Pedúnculo opercular de 4.3 mm (n=4, r: 2-5.5, µ=3.7 ±1.5), copa opercular de 1.5 mm (n=4, r: 0.8-2, µ=1.5 ±0.5), es semiesférico, con una parte basal blanca y blanda y una sección distal hialina-amarilla y con forma de sarakof (sombrero de cazador) (Fig. 12q), con cinco discos fusionados (n=4, r: 2-5, µ=4 ±1.4). En los demás ejemplares existen ligeras variantes en la forma (Fig. 12r-s). Tórax de 3.2 mm de largo (n=3, r: 2-3.2, µ=2.5 ±0.6) y 1.9 mm de ancho (n=3, r: 0.5-1.9, µ=1.2 ±0.7). Membrana torácica llega hasta el quinto setígero (n=3, r: 4-5). Setas del collar capilares y limbadas. El resto de los setígeros torácicos presentan setas limbadas y "Apomatus". Uncinos con diez dientes aguzados en una hilera, el primero es ancho y truncado. Abdomen con setas geniculadas anteriores y capilares en los segmentos posteriores, primeros segmentos abdominales asetígeros. Uncinos abdominales posteriores con varias hileras de dientes, el primero más ancho y aguzado.
V. annulata trapezoidal
(Fig. 12t-h, 13a-b)
Material examinado: 33 ejemplares. AVE3(1), BA1(1), BUE1(1), CHA13-14(5), CON3(16), CU4(3), EL2774(1), NC1(1), NC2(1), PO75006(1), XAH3(2).
Caracterización: Tubo blanco con tres quillas longitudinales y peristomas prominentes (Fig. 12t-u). Cuerpo amarillo de 7 mm (n=20, r: 2.6-18.3, µ=8.4 ±4.5). Pedúnculo opercular de 2 mm (n=22, r: 0.9-5.3, µ=2.3 ±1.2), copa opercular de 0.6 mm (n=22, r: 0.2-2.3, µ=1 ±0.6), semiesférico, con una parte basal blanca y blanda y una sección distal amarilla con forma trapezoidal (Fig. 12v-w), sin rastros de discos fusionados, aunque otros ejemplares (Fig. 12x) tienen hasta cinco discos fusionados (n=22, r: 1-5, µ=2.1 ±1.2). Tórax de 1.8 mm de largo (n=22, r: 0.8-4, µ=2 ±0.9) y 0.4 mm de ancho (n=22, r: 0.2-2.2, µ=0.7 ±0.6). Membrana torácica llega hasta el cuarto setígero (n=22, r: 3-5). Setas del collar capilares finamente serradas (Fig. 12y) y limbadas (Fig. 12z). El resto de los setígeros torácicos presentan setas limbadas (Fig. 12a) y "Apomatus" (Fig. 12b-c). Uncinos con 10-11 dientes aguzados en una hilera, el primero es ancho y truncado o en ocasiones aguzado (Fig. 12d-e). Abdomen con setas geniculadas anteriores (Fig. 12h) y capilares en los segmentos posteriores, primeros segmentos abdominales asetígeros. Uncinos abdominales posteriores con varias hileras de dientes, el primero más ancho y obtuso (Fig. 12f-g).
Observaciones: Los caracteres tradicionales (opérculo, setas del collar) no han permitido separar a las especies, aunque hay evidencia de que la ornamentación de los tubos puede indicar diferencias interespecíficas (ver Clave). En el Caribe se han descrito siete especies: V. annulata, V. annulituba (Augener, 1906), V. bermudensis (Bush, 1904), V. amblia (Bush, 1907), V. intermedia (Bush, 1907), V. ehlersiana (Bush, 1910) y V. glandulata Treadwell, 1936. Los límites morfológicos entre ellas no son claros (ten Hove & Wolf 1984). Zibrowius (1970) comentó sobre esta confusión y mantiene las formas del Gran Caribe como Vermiliopsis sp. Por su parte, ten Hove y Wolf (1984) proponen que las formas de esta región se nombren temporalmente como V. annulata mientras se aclaran los límites morfológicos entre las especies. Consideran que por lo menos deben existir dos especies con ecología y tubos diferentes: V. annulata con cinco quillas longitudinales y otra especie, aún indescrita, con tres quillas (ten Hove & San Martín 1995).
Nuestros ejemplares no permiten separación entre especies identificables por el número de quillas longitudinales del tubo y las características del opérculo. Los tubos por lo general están fragmentados, algunos no presentan rastros de quillas, otros presentan tres o cinco quillas. La mayoría también presentan peristomas bien marcados. La relación entre la longitud del tórax y la del pedúnculo opercular, considerando las tres formas, es isométrica (Fig. 3a), pero no muestra separación de grupos (Fig. 13a). Sin embargo, el número de discos operculares segrega a las trapezoidales (Fig. 13b), las cuales son más pequeñas y presentan pocos discos fusionados. Por el contrario, los ejemplares con opérculo embudo son más grandes y con mayor número de discos. Los ejemplares con opérculos sarakof son intermedios. Aunque las líneas de tendencia son diferentes, existe mucho traslape en los datos, lo cual no permite una separación definitiva entre las tres formas (Fig. 13b). Es necesario evaluar las características anteriores con mas ejemplares. Asimismo, debe realizarse una revisión del género que incluya la búsqueda de nuevos caracteres morfológicos (y análisis moleculares de tejidos frescos), toda vez que las características del tubo pueden prestarse a confusión por la acción de los epibiontes. También debe observarse la relación que puede haber entre las especies de Vermiliopsis y el sustrato donde se fijan, así como sus patrones reproductivos.
Observaciones de V. glandulata (Fig. 12i): De 10 mm de largo y 1 mm de ancho torácico, sin radiolos. Pedúnculo opercular anulado, junto con la copa tiene 4 mm de largo. Base de la copa opercular blanda y blanca, la parte media distal presenta ocho anillos de quitina amarillo oscuro, y el extremo distal está roto (le faltan algunos anillos). Ten Hove examinó el ejemplar y anotó en una etiqueta: "A partir de la descripción el ejemplar marcado como tal por mí, debe ser el holotipo. El otro ejemplar, dañado, puede ser el segundo mencionado por Treadwell y entonces es de otra localidad, sea "Castle Harbor Reef".
Distribución y hábitat: Gran Caribe. Fondos mixtos y fondos rocosos con algas, pastos marinos (Thalassia), o corales.
Género A
(Fig. 14a-h)
Material examinado: AVE4(16).
Caracterización: Animal gregario. Algunos fragmentos del tubo son lisos pero otros fragmentos presentan 2-3 quillas longitudinales. Todos los ejemplares están incompletos, su cuerpo es amarillo. Corona con cuatro radiolos por lado, con una expansión cilíndrica distal en dos de los radiolos, a manera de opérculos (Fig. 14a). Tórax con nueve segmentos torácicos (n=14, r: 7-9, µ=7.8 ±0.8). Membrana torácica poco desarrollada, llega hasta el quinto setígero torácico. Setas del collar limbadas (Fig. 14b). El resto de los setígeros torácicos tienen setas limbadas (Fig. 14d) y "Apomatus" (Fig. 14c) a partir del quinto setígero (n=15, r: 2-5, µ=2.9 ±0.9). Uncinos rectangulares con 12-15 dientes aguzados dispuestos en una hilera (Fig. 14e-f). Abdomen con setas geniculadas (Fig. 14g). Uncinos más pequeños que los torácicos, con 9-10 dientes aguzados, el primero más largo y obtuso (Fig. 14h).
Distribución y hábitat: Caribe mexicano. Sobre rocas coralinas.
Observaciones: Son pocos los géneros de serpúlidos que presenten más de siete setígeros torácicos; sin embargo, la combinación de caracteres de los ejemplares revisados no permite ubicarlos dentro de alguno de estos géneros. Amphiserpula Uchida, 1978 tiene 9-10 setígeros torácicos pero el opérculo es similar al de Serpula; Filograna Oken, 1815 presenta 5-12 setígeros torácicos pero sus setas del collar son modificadas, con un limbo distal y un hombro denticulado; Filogranella Ben-Eliahu & Dafni, 1979 tiene 11-14 setígeros torácicos pero no tiene opérculo; Membranopsis Bush, 1910 presenta nueve setígeros torácicos, se desconoce si tiene opérculo, pero presenta una membrana torácica muy desarrollada semejante a la de Protula (cf. Uchida 1978); Salmacina tiene entre 5-9 setígeros torácicos pero sus setas del collar son modificadas, con un limbo distal y un hombro denticulado y no tiene opérculo; Salmacinopsis Bush, 1910, tiene nueve setígeros torácicos pero no tiene opérculo; Spiraserpula tiene 6-9 setígeros torácicos pero su opérculo es como los de Serpula. Necesitamos mejores materiales para completar la caracterización.
Clave para los serpúlidos del Gran Caribe
(* no registrados en la región)
| 1 | Sinopérculo | 2 |
| - | Con opérculo | 7 |
| 2(1) | Setas del collar limbadas o capilares; uncinos con diente principal muy largo | 3 |
| - | Setas del collar modificadas, con limbo distal; uncinos con diente principal no tan largo | 6 |
| 3(2) | Siete a nueve setígeros torácicos | 4 |
| - | Once a 14 setígeros torácicos | Filogranella sp.1 |
| 4(3) | Siete setígeros torácicos; corona radiolar con más de 10 radiolos | 5 |
| - | Nueve setígeros torácicos; corona radiolar con hasta 5 pares de radiolos | Salmacinopsis setosa2 |
| - | Nueve setígeros torácicos; corona radiolar desconocida; | Membranopsis inconspicua3 |
| 5(4) | Setas del collar capilares (sin limbos); corona con 20 radiolos por lado; uncinos torácicos con hasta 16 dientes | Subprotula appendiculata4 |
| - | Setas del collar limbadas | Protula27 |
| 6(2) | Setas abdominales geniculadas | Salmacina... 32 |
| - | Setas abdominales casi rectas; corona con hasta 23 radiolos por lado | Protis arctica (Hansen, 1878)5 |
| 7(1) | Opérculo sobre un radiolo | 8 |
| - | Opérculo sobre un pedúnculo modificado | 12 |
| 8(7) | Cinco setígeros torácicos | Josephella marenzelleri |
| - | Seis a 12 setígeros torácicos | 9 |
| 9(8) | Setas del collar simples y limbadas aguzadas | 10 |
| Setas del collar modificadas, con limbo distal | 11 | |
| 10(9) | Sólo 7 setígeros torácicos, membrana torácica hasta el séptimo setígero torácico; opérculo esférico | Apomatus sp. A ten Hove & Wolf |
| Siete a nueve setígeros torácicos; membrana torácica hasta el quinto setígero torácico; opérculo ovoidal con una proyección distal | Género A | |
| - | Nueve setígeros torácicos, membrana torácica hasta el quinto setígero torácico | Membranopsis inconspicua3 |
| 11(9) | Opérculo como un cono ligeramente deprimido dispuesto sobre un engrosamiento esférico del pedúnculo opercular | Filogranula calyculata |
| Opérculo como una esfera con una depresión media, pedúnculo opercular sin engrosamiento subdistal | Filograna huxleyi6 | |
| 12(7) | Pedúnculo opercular con alas o espinas | 13 |
| - | Pedúnculo opercular sin alas ni espinas | 17 |
| 13(12) | Pedúnculo opercular con 4 espinas o procesos digitados radiales; opérculo abocinado, crenulado, formado por muchas proyecciones radiales fusionadas | Crucigera websteri |
| - | Pedúnculo opercular con alas | 14 |
| 14(13) | Collar sin setas; opérculo sin proyecciones | Pomatoleios* |
| - | Collar con setas; opérculo variable | 15 |
| 15(14) | Opérculo plano o sin espinas; setas del collar limbadas | Pomatoceros... 37 |
| - | Opérculo con espinas o discos; setas del collar modificadas | 16 |
| 16(15) | Opérculo arborescente, con espinas semejantes a cuernos de venado | Spirobranchus... 34 |
| - | Opérculo con una columna de discos concéntricos | Pomatostegus stellatus |
| 17(12) | Collar sin setas | 18 |
| - | Collar con setas | 20 |
| 18(17) | Tubo libre con forma de colmillo; opérculo globular, distalmente plano y liso, raramente con una espina central; setas abdominales capilares, sin setas geniculadas | Ditrupa arietina* |
| - | Tubo cementado, con anulaciones | 19 |
| 19(18) | Tubo opaco sin quilla dorsal, a veces con cápsulas de incubación; abdomen con 11-15 setígeros; setas abdominales geniculadas; opérculo con espinas romas, a veces bífidas, si está colapsado las espinas parecen cubiertas por una membrana transparente | Rhodopsis pusilla7 |
| - | Tubo hialino con quilla dorsal, sin cápsulas de incubación; abdomen con 60 setígeros; una banda de ocelos en lugar de las setas del collar; setas abdominales "trompeta" con diente largo; opérculo desconocido | Placostegus incomptus |
| 20(17) | Setas abdominales "trompeta" | 21 |
| - | Setas abdominales geniculadas | 23 |
| 21(20) | Opérculo como un embudo sencillo; setas del collar dentadas en la base | 22 |
| - | Opérculo complejo, con un embudo basal y un verticilo distal | Hydroides... 39 |
| 22(21) | Tubo sin costillas longitudinales internas | Serpula... 53 |
| - | Tubo con costillas u otras estructuras longitudinales internas, por lo menos en las porciones más antiguas del mismo | Spiraserpula... 54 |
| 23(20) | Tórax sin setas falciformes ("Apomatus") | 24 |
| - | Tórax con setas falciformes | 26 |
| 24(23) | Pedúnculo opercular blando, no calcificado | 25 |
| - | Pedúnculo opercular calcificado; uncinos torácicos con dientes en una hilera, los abdominales con dientes en varias hileras | Sclerostyla... 61 |
| 25(24) | Siete setígeros torácicos; setas abdominales geniculadas; uncinos abdominales con menos de 15 dientes vistos de perfil | Ficopomatus... 63 |
| - | Seis setígeros torácicos; setas abdominales distalmente geniculadas; uncinos abdominales con más de 15 dientes vistos de perfil; corona con 12 radiolos por lado | Hyalopomatus langerhansi |
| 26(23) | Membrana torácica llega hasta los setígeros 3-6; uncinos torácicos con diente anterior entero | Vermiliopsis8... 64 |
| - | Membrana torácica llega al setígero 2; uncinos torácicos con diente anterior bifurcado | Pseudovermilia... 66 |
| 27(5) | Tubo con quillas longitudinales; corona con 15 radiolos por lado | P. longiseta9 |
| - | Tubo sin quillas longitudinales | 28 |
| 28(27) | Tubo liso o tuberculado | 29 |
| - | Tubo anulado | 31 |
| 29(28) | Tubo liso; corona con 12-15 radiolos por lado | P. tubularia |
| - | Tubo con tubérculos (textura áspera); corona con más de 15 radiolos | 30 |
| 30(29) | Corona con 20 radiolos por lado | P. diomedeae |
| - | Corona con 30-40 radiolos por lado | P. balboensis |
| 31(28) | Corona con 12 radiolos por lado | P. alba |
| - | Corona con 25-50 radiolos por lado | P. submedia10 |
| 32(6) | Setas del collar con tres tamaños muy diferentes | S. amphidentata |
| - | Setas del collar muy similares entre sí | 33 |
| 33(32) | Setas del primer setígero con incontables dientes pequeños | S. dysteri11 |
| - | Setas del primer setígero con 4-7 dientes grandes | S. incrustans11 |
| 34(16) | Pedúnculo opercular con alas cortas, generalmente fimbriadas; opérculo con 3 grupos de cuernos ramificados | S. dendropoma |
| - | Pedúnculo opercular con alas grandes, no fimbriadas | 35 |
| 35(34) | Opérculo con 2-3 cuernos grandes y delgados, generalmente ramificados, cuyo patrón de crecimiento los aleja de la base opercular | S. giganteus12 |
| - | Opérculo con cuernos anchos, extendidos sobre la base opercular | 36 |
| 36(35) | Opérculo con unos 7 cuernos con puntas bífidas o trífidas | S. polycerus |
| - | Opérculo con 2-3 cuernos, a veces con las puntas hendidas | S. polycerus augeneri |
| 37(15) | Tubo con alvéolos entre las costillas longitudinales; opérculo con 2 tubérculos diminutos | P. cf. minutus |
| - | Tubo sin alvéolos; opérculo variable | 38 |
| 38(37) | Opérculo piriforme, con una proyección | P. americanus |
| - | Opérculo variado, puede presentar tres rebordes o cuernos | S. polycerus13 |
| 39(21) | Espinas del verticilo opercular con espínulas laterales pares | grupo Hydroides14... 40 |
| - | Espinas del verticilo opercular sin espínulas laterales, a veces con alas grupo Eupomatus14 | 45 |
| 40(39) | Espinas con un solo par de espínulas laterales | 41 |
| - | Espinas con varios pares de espínulas laterales basales | 43 |
| 41(40) | Embudo opercular formado por radios; espinas incurvadas, aguzadas | 42 |
| - | Embudo opercular formado por radios; espinas rectas, punta roma | H. mucronatus |
| - | Embudo opercular sin radios pero con 3-6 procesos obtusos; espinas rectas, punta roma | H. cf. mucronatus |
| 42(41) | Espinas con una proyección dorsal redondeada, muy incurvadas en ángulo recto (semejan la cabeza de un buitre) | H. parvus15 |
| - | Espinas sin proyección dorsal, incurvadas de forma suave; embudo opercular con unos 16 radios | H. bispinosus15 |
| 43(40) | Verticilo con sólo 5 espinas; espinas ensanchadas basalmente | Hydroides sp. (Cuba) |
| - | Verticilo con más de 5 espinas; espinas separables en dos porciones, basal oscura con espínulas laterales y distal clara, todas incurvadas | H. mongeslopezi |
| - | Verticilo con más de 5 espinas; espinas simples con espínulas laterales | 44 |
| 44(43) | Espinas del verticilo dirigidas hacia afuera | H. elegans |
| - | Espinas del verticilo incurvadas | Hydroides sp. A ten Hove & Wolf |
| 45(39) | Espinas con alas laterales; espínulas internas del verticilo con tubérculo subdorsal | H. alatalateralis16 |
| - | Espinas sin alas laterales o muy poco desarrollados; espínulas internas del verticilo sin tubérculo subdorsal | 46 |
| 46(45) | Embudo opercular con radios distalmente expandidos lateralmente, con forma de T | H. gairacensis |
| - | Embudo opercular con radios aguzados | 47 |
| 47(46) | Radios del embudo opercular con una protuberancia interna distal | H. floridanus16 |
| - | Radios del embudo opercular sin protuberancia distal | 48 |
| 48(47) | Verticilo con espinas dirigidas hacia arriba, cada espina roma, ligeramente capitada | H. microtis |
| - | Verticilo con algunas espinas incurvadas | 49 |
| 49(48) | Verticilo con todas las espinas dirigidas hacia afuera | H. protulicola |
| - | Verticilo con espinas incurvadas, por lo menos las mayores | 50 |
| 50(49) | Espinas con puntas dilatadas, en forma de T | H. diramphus |
| - | Espinas aguzadas | 51 |
| 51(50) | Espinas sin espínulas internas; las mayores geniculadas, las menores dirigidas hacia afuera | H. dianthus |
| - | Espinas geniculadas, con espínulas internas | 52 |
| 52(51) | Espinas con una larga espínula | H. spongicolus |
| - | Espinas con dos espínulas cortas internas | H. sanctaecrucis |
| 53(22) | Embudo opercular con 20-40 radios; pedúnculo liso; tubo con 5-7 costillas longitudinales, a veces cada costilla parece llevar esínulas | Serpula vermicularis17 |
| - | Embudo opercular con 26-38 radios; pedúnculo liso; tubo pardo con 9 costillas longitudinales | Serpula sp. |
| - | Embudo opercular con 18 radios; pedúnculo con un pliegue; tubo desconocido | Serpula sp. A ten Hove & Wolf |
| 54(22) | Tubo con costilla interna dorsal o ventral | 55 |
| - | Tubo con costillas internas dorsal y ventral | 56 |
| 55(54) | Tubo con costilla dorsal no serrada, en forma de V invertida | Spiraserpula singularis |
| - | Tubo con costilla ventral lisa, raramente con espinas dorsales | S. nudicristata |
| 56(54) | Costilla dorsal serrada, la ventral en forma de Y | 57 |
| - | Costilla dorsal no serrada, la ventral serrada | 58 |
| 57(56) | Hasta 7 pares de radiolos; uncinos torácicos sin tubérculos laterales; más de 100 segmentos abdominales | S. ypsilon |
| - | Hasta 11 pares de radiolos; uncinos torácicos con tubérculos laterales; menos de 100 segmentos abdominales | S. paraypsilon |
| 58(57) | Tubo con costillas accesorias | 59 |
| - | Tubo sin costillas accesorias | 60 |
| 59(58) | Costillas accesorias dorsolaterales; tubo blanco; con opérculo | S. plaiae |
| - | Costillas accesorias laterales; tubo rosado o con bandas longitudinales rosadas; sin opérculo | S. caribensis |
| 60(58) | Con opérculo; abdomen con unos 40 setígeros | S. karpatensis |
| - | Sin opérculo; abdomen con unos 50 setígeros | S. zibrowii |
| 61(24) | Tubo con 6-12 costillas longitudinales irregulares; embudo opercular somero | Sclerostyla differens |
| - | Tubo con tres o menos costillas longitudinales; embudo opercular profundo | 62 |
| 62(61) | Tubo con anulaciones prominentes | S. semiannulata |
| - | Tubo con unas 3 costillas longitudinales regulares; embudo | S. ctenactis |
| 63(25) | Opérculo cóncavo con espinas incurvadas (subtropical) | F. enigmaticus |
| - | Opérculo ligeramente cóncavo sin espinas (tropical) | F. miamensis |
| 64(26) | Tubo con anulaciones marcadas (peristomas), sin crestas longitudinales | V. annulituba18 |
| - | Tubo con crestas longitudinales, raramente con anulaciones | 65 |
| 65(64) | Tubo con crestas longitudinales dentadas, raramente con anulaciones; opérculo con 1-3 discos anchos fusionados | V. annulata19 |
| - | Tubo con 5 crestas longitudinales lisas, 2 mayores alternas, sin anulaciones; opérculo con más de 5 discos fusionados | V. bermudensis20 |
| - | Tubo con 5 crestas longitudinales lisas, poco marcadas, sin anulaciones; opérculo con menos de 5 discos fusionados | V. intermedia |
| 66(26) | Opérculo negro o pardo oscuro | 67 |
| - | Opérculo no negro, endurecido o blando | 68 |
| 67(66) | Tubo con cresta e hileras longitudinales de alvéolos; opérculo con espinas arborescentes | P. multispinosa |
| - | Tubo con cresta longitudinal y surcos transversales, a veces con anillos; opérculo con una espina curva simple, con varias espinas menores o sin espinas | P. occidentalis |
| 68(66) | Tubo con bandas pardas transversas o completamente pardo; opérculo con surcos anulados profundos, a veces con una espina distal simple o ramificada | P. fuscostriata |
| - | Tubo blanco; opérculo con surcos anulados sutiles o liso, transparente o blanco | 69 |
| 69(68) | Tubo con anulaciones, pueden faltar; opérculo con surcos anulados, a veces con una espina distal | P. holcopleura |
| - | Tubo con 2-3 crestas longitudinales dentadas; opérculo liso | P. madracicola |
Notas
2 Ten Hove (com. pers. 1999) revisó el material tipo y confirmó la presencia de membrana torácica (llega hasta el sexto setígero) y más de cinco pares de radiolos. La única característica de esta especie es la hilera sencilla de pínulas en los radiolos (en lugar de doble); este carácter necesita confirmación.
3 Descrita por Bush (1910) para Bermuda. El material tipo está dañado y no se puede precisar si tenía opérculo; sin embargo, su membrana torácica es muy desarrollada, semejante a la de Protula; fue notado por Fauchald (1977) y Uchida (1978). Por nuestro desconocimiento, la especie tiene dos salidas en la clave.
4 Incluye Subprotula longiseta Bush, 1910 y Protula longisetoides Rioja, 1942.
5 Zibrowius (1969) consideró que P. simplex Ehlers, 1887 es sinónimo menor y que las diferencias usadas para la separación no son significativas.
6 Filograna es un género muy cercano a Salmacina, pero se diferencía de este por la presencia de un opérculo (Gee 1963).
7 Redescripción en Ben-Eliahu & ten Hove (1989).
8 Las diferencias entre las especies son muy sutiles y deben revisarse los límites morfológicos de las mismas (cf. Zibrowius 1970); sin embargo, se incluyen las especies reconocibles (V. amblia fue registrada en el Caribe pero no podemos distinguirla por que no se describió el tubo).
9 Podría incluir a P. antennata Ehlers, 1887.
10 Descrita por Augener (1906), Hartman (1959) la supuso sinónima de P. diomedeae pero son distintas.
11 Debe revisarse la variación en la denticulación de las setas del collar de estas dos especies tal como lo recomiendan ten Hove & Wolf (1984).
12 S. giganteus s.l. es separable en cuando menos tres complejos de especies: S. giganteus, S. corniculatus (Grube, 1862) y S. gardineri (Pixell, 1913), cada una con varias especies (Fiege y ten Hove 1999).
13 Es poco probable que los registros previos de P. triqueter (Linnaeus, 1767) en la región sean válidos. Fue descrita para el Atlántico europeo; el registro de Jones et al. (1986) en realidad se trata de S. polycerus (ten Hove, com. pers. 1999).
14 Estos grupos reflejan el uso tradicional pero no son unidades naturales.
15 Se han confundido H. bispinosus e H. parvus con H. cruciger, sin embargo, ésta última se restringe al Pacífico Oriental tropical (ten Hove, com. pers. 1999).
16 H. alatalateralis y H. floridanus Bush, 1910 (= "H. uncinatus" de Ehlers 1887) forman un complejo de especies. Al parecer se diferencían por su distribución geográfica, la primera restringida a las aguas tropicales del Caribe y la segunda a aguas subtropicales y templadas (ten Hove, com. pers. 1999).
17 S. sombreriana McIntosh, 1885 fue descrita con un ejemplar deteriorado y sin opérculo por lo que es cuestionable. Ten Hove transfirió tentativamente el material tipo a Hyalopomatus langerhansi Ehlers, 1887, pero no se ha realizado una argumentación formal (Ben-Eliahu & Fiege 1996).
18 Descrita por Augener (1906), se ha confundido con V. infundibulum debido a que ambas especies presentan tubo con anulaciones, pero la segunda fue descrita originalmente del Mediterráneo.
La recolecta fue autorizada por SEMARNAP (permiso DOO750-1578). Leopoldo Cosgalla (Quelonios A.C.) apoyó en la recolecta en Champotón y junto con Wilbert Borges (ITMAR-Champotón) nos brindaron espacio para trabajar. Adrián Marfil apoyó en la recolecta en Ría Lagartos. Se agradece la hospitalidad y apoyo para la realización de las recolectas en Isla Contoy al personal del Parque Nacional, en especial a Mario Lara, César Sánchez, Omar Ortiz, Adriana Amador, Dorina Basurto y Andrés Mariano. Asimismo, agradecemos las facilidades para recolectar en Isla Cozumel al personal del Parque Marino Nacional, en especial a Elvira Carvajal, Juan Domínguez y Fernando Orozco; el vehículo utilizado en la isla fue amablemente proporcionado por Alberto Sleme (Empresas Turísticas Nacionales). El material de Cuba fue prestado gentilmente por Diana Ibarzábal (IO-Cuba) y otros ejemplares del golfo de México fueron prestados por Angel de León-González (UANL). Se agradece el apoyo para realizar una estancia y revisar colecciones a Patricia Gómez (ICML-UNAM), Francisco Solís (ICML-UNAM), Vivianne Solís (ICML-UNAM) y Juan Manuel Vargas (UV). Elba Escobar y Luis Soto (ICML-UNAM) proporcionaron los materiales del "Edwin Link". Humberto Bahena imprimió las fotografías utilizadas en este estudio. El trabajo de campo y las visitas a las colecciones nacionales fueron financiadas por el Programa de Apoyo a Tesis de Maestría de ECOSUR, recursos fiscales y por un proyecto CONACYT (4120P-N9607). El apoyo de Scott Monks y Griselda Pulido para la digitalización de las figuras, así como la asistencia en la edición de las mismas por Luis Carrera-Parra fue vital para la conclusión del presente trabajo. Angel de León-González hizo comentarios sobre una versión previa de este trabajo. JRBZ agradece el tiempo que por derecho correspondía a su pareja, Socorro García-Madrigal e hijas Karla y Marcela. El segundo autor visitó el American Museum of Natural History y agradece el apoyo recibido a Judith Winston y Jerry Thurmond; la visita fue financiada en parte por la Lerner-Gray Fund for Marine Research del AMNH. La revisión crítica de Harry ten Hove (Universidad de Amsterdam) y de dos árbitros anónimos permitió mejorar la calidad del escrito; el primero también envió información crítica.
Resumen
A partir de la revisión de más de 1 250 ejemplares de serpúlidos, 15 especies de la región del Gran Caribe fueron identificadas y caracterizadas. Trece especies fueron recolectadas en las costas de la Península de Yucatán y ocho fueron encontradas en otras localidades en el Golfo de México, otras siete son de Cuba y se incluyeron comentarios sobre el material tipo de dos especies más. Se realizaron tres análisis morfométricos para evaluar algunos caracteres. El primero sobre Pomatostegus stellatus (Abildgaard) y especies y subespecies relacionadas: P. brachysoma Schmarda, P. macrosoma Schmarda, P. s. fruticosa Mörch, P. s. pentapoma Mörch y P. s. tetrapoma Mörch, indicando que estas son con-específicas. Otro fue sobre las especies de Spirobranchus de Blainville: S. dendropoma Mörch, S. giganteus (Pallas), S. polycerus (Schmarda) y S. polycerus augeneri ten Hove; se demostró que S. giganteus es diferente de los otros tres táxones. El tercer análisis fue realizado sobre Vermiliopsis annulata (Schmarda) s.l., y permitió reconocer tres formas diferentes. Asimismo, varios ejemplares incompletos probablemente pertenecen a un género indescrito. Se incluye una clave de identificación para todas las especies registradas en la región del Gran Caribe
Referencias
Augener, H. 1906. Westindische Polychaeten. Bull. Mus. Comp. Zool. Harv. Coll. 43: 90-196. [ Links ]
Augener, H. 1925. Üeber Westindische und einige andere polychaeten-typen von Grube (Ærsted), Kröyer, Mörch und Schmarda. Univ. Zool. Mus. Copenhagen, Pub. 39: 1-47. [ Links ]
Bastida-Zavala, J.R. & S. Monks. En prep. Phylogenetic relationships among the serpulid genera (Polychaeta: Serpulidae). Zool. Scripta. [ Links ]
Benedict, J.E. 1887. Descriptions of ten species and one new genus and one new genus of Annelids from the dredgings of the the U.S. Fish Comission steamer "Albatross". Proc. U.S. Natl. Mus. 9: 547-553. [ Links ]
Ben-Eliahu, M.N. & J. Dafni. 1979. A new reef-building serpulid genus and species from the Gulf of Elat and the Red Sea, with notes on other gregarious tubeworms from Israeli waters. Israel. J. Zool. 28: 199-208. [ Links ]
Ben-Eliahu, M.N. & D. Fiege. 1996. Serpulid tube-worms (Annelida: Polychaeta) of the Central and Eastern Mediterranean with particular attention to the Levant Basin. Senckenb. Marit. 28: 1-51. [ Links ]
Ben-Eliahu, M.N. & ten Hove, H.A. 1989. Redescription of Rhodopsis pusilla Bush, a little known but widely distributed species of Serpulidae (Polychaeta). Zool. Scripta 18: 381-395. [ Links ]
de León-González, J.A., S.I. Salazar-Vallejo & H. Salaices-Polanco. 1989. Claves ilustradas para familias y géneros, p. 34-131. In Poliquetos (Annelida: Polychaeta) de México. Libros Universitarios, UABCS, La Paz, México. [ Links ]
de Quatrefages, A. 1865. Histoire Naturelle des Annéles Marines et d'Eau Douce: Annéelides et Géphyriens. Vol. 1. Librairie Encyclopédique Roret, Paris. 588 p. [ Links ]
deVantier, L.M., R.E. Reichelt & R.H. Bradbury. 1986. Does Spirobranchus giganteus protect host Porites from predation by Acanthaster planci: predator pressure as a mechanism of coevolution? Mar. Ecol. Progr. Ser. 32: 307-310. [ Links ]
Fauchald, K. 1977. The Polychaete Worms, definitions and keys to the orders, families and genera. Nat. Hist. Mus. Los Angeles Cty., Sci. Ser. 28: 1-190. [ Links ]
Fauchald, K. & G. Rouse. 1997. Polychaete systematics: Past and present. Zool. Scripta 26: 71-138. [ Links ]
Fauvel, P. 1927. Polychètes Sédentaires et Addenda aux Polychètes Errantes, Archiannélides, Myzostomaires. Faune de France 16: 1-494. [ Links ]
Fiege, D. & H.A. ten Hove. 1999. Redescription of Spirobranchus gaymardi (Quatrefages, 1866) (Polychaeta: Serpulidae) from the Indo-Pacific with remarks on the Spirobranchus giganteus complex. Zool. J. Linn. Soc. 126: 355-364. [ Links ]
Fosså, & A.J. Nilsen. 1996. Korallenriff-aquarium 5. Einzellige organismen, schwämme, marine würmer und weichtiere im korallenriff und für das Korallenriff-Aquarium. Birgit Schmettkamp , Alemania. 352 p.
Gee, J.M. 1963. On the taxonomy and distribution in South Wales of Filograna, Hydroides and Mercierella (Polychaeta: Serpulidae). Ann. Mag. Nat. Hist., ser. 13, 6: 705-715. [ Links ]
Grube, A.E. 1851. Annulaten, p. 1-24. In Reise in den äussersten Norden und Osten Sibiriens während der Jahre 1843 und 1844, mit allerhöchster Genehmigung auf Veranstaltung der kaiserlichen Akademie der wissenschaften zu St. Petersburg. Heraus. Middendorff, 2. [ Links ]
Grube, A.E. 1878. Annulata Semperiana. Beitrage zur kenntnis der Annelidenfauna der Philippinen nach den von Herrn Prof. Semper mitgebrachten sammlungen. Mém. Acad. Imp. Sci. St. Pétersb. 7ème. Sér., 25: 1-300. [ Links ]
Hanson, J. 1948. The genera Apomatus and Protula (Polychaeta, Serpulidae). J. Mar. Biol. Ass. U.K. 27: 581-584. [ Links ]
Hartman, O. 1956. Polychaetous annelids erected by Treadwell, 1891 to 1948, together with a brief chronology. Bull. Amer. Mus. Nat. Hist. 109: 243-310. [ Links ]
Hartman, O. 1959. Catalogue of the polychaetous annelids of the World. Allan Hancock Found. Publ. Occ. Pap. 23: 1-628. [ Links ]
Hartmann-Schröder, G. 1996. Annelida, Borstenwürmer, Polychaeta. Die Tierwelt Deutschlands 58, Fischer, Jena. 648 p. [ Links ]
Hunte, W., B.E. Conlin & J.R. Marsden. 1990a. Habitat selection in the tropical polychaete Spirobranchus giganteus, 1. Distribution on corals. Mar. Biol. 104: 87-92. [ Links ]
Hunte, W., J.R. Marsden & B.E. Conlin. 1990b. Habitat selection in the tropical polychaete Spirobranchus giganteus, 3. Effects of coral species on body size and body proportions. Mar. Biol. 104: 101-107. [ Links ]
Jones, M.L., M.E. Petersen & W. Sterrer. 1986. Phylum Annelida, p. 232-258. In Sterrer, W. (ed.). Marine fauna and flora of Bermuda: A systematic guide to the identification of marine organisms. Wiley, Nueva York. 742 p. [ Links ]
Johnston, G. 1865. A catalogue of the British non-parasitical worms in the collection of the British Museum. London. 366 p. [ Links ]
Lamarck, J.B. Histoire naturelle des animaux sans vertèbres... lexposition des principes fondamentaux de la Zoologie. Deterville Libr. Verdiere Libr., Paris, 5. 612 p. [ Links ]
Lechapt, J.-P. 1992. Neovermilia anoperculata, nouvelle espèce bathyale de Nouvelle-Calédonie (Polychaeta, Serpulidae). Bull. Mus. Natl. Hist. Nat. Paris, 4a sér, sec. A, 14: 443-448. [ Links ]
Liñero-Arana, I. 1999. Serpulidae (Annelida: Polychaeta) de la costa nororiental de Venezuela. Bol. Inst. Oceanogr. Venezuela (38): 00-00. [ Links ]
Marsden, J.R. 1987. Coral preference behaviour by planktotrophic larvae of Spirobranchus giganteus corniculatus (Serpulidae: Polychaeta). Coral Reefs 6: 71-74. [ Links ]
Marsden, J.R. 1992. Reproductive isolation in two forms of the serpulid polychaete, Spirobranchus polycerus (Schmarda) in Barbados. Bull. Mar. Sci. 51: 14-18. [ Links ]
Marsden, J.R., B.E. Conlin & W. Hunte. 1990. Habitat selection in the tropical polychaete Spirobranchus giganteus, 2. Larval preference for corals. Mar. Biol. 104: 93-99. [ Links ]
McIntosh, W.C. 1923. A monograph of the British marine annelids. Vol. 4, Part 2: Polychaeta: Sabellidae to Serpulidae. With additions to the British marine Polychaeta during the publication of the monograph. London, Ray Soc. 4: 251-538. [ Links ]
Monro, C.C.A. 1933. On a collection of Polychaeta from Dry Tortugas, Florida. Ann. Mag. Nat. Hist., ser. 10, 12: 244-269. [ Links ]
Mörch, O.A.L. 1863. Revisio critica Serpulidarum et bidrag til Rørormens Naturhistorie. Naturhist. Tidsskr. Copenhagen 3: 347-470. [ Links ]
Nishi, E. 1993. Notes on reproductive biology of some serpulid polychaetes of Sesoko Island, Okinawa, with brief accounts of setal morphology of three species of Salmacina and Filograna implexa. Mar. Foul. 10: 11-16. [ Links ]
Nishi, E. & M. Nishihira. 1996. Age-estimation of the Christmas Tree worm Spirobranchus giganteus(Polychaeta, Serpulidae) living buried in the coral skeleton from the coral-growth band of the host coral. Fish. Sci. 62: 400-403. [ Links ]
Perkins, T.H. 1998. Checklist of shallow-water marine polychaetous Annelida of Florida, p. 79-122. In Camp, D.K., W.G. Lyons & T.H. Perkins (eds.). Checklist of selected shallow-water marine invertebrates of Florida. Florida Mar. Res. Inst. Tech. Rep. 3, St. Petersburg. [ Links ]
Perkins, T.H. & T. Savage. 1979. A bibliography and checklist of polychaetous annelids of Florida, the Gulf of Mexico, and the Caribbean Region. Florida Mar. Res. Publ. 14: 1-62. [ Links ]
Pillai, T.G. 1960. Some marine and brackish-water serpulid Polychaeta from Ceylon, including new genera and species. Ceylon J. Sci. (Bio. Sci.) 3: 1-40. [ Links ]
Pillai, T.G. & H.A. ten Hove. 1994. On Recent species of Spiraserpula Regenhardt 1961, a serpulid polychaete genus hitherto known only from Cretaceous and Tertiary fossils. Bull. Nat. Hist. Mus. 60: 39-104. [ Links ]
Rafinesque, C.S. 1815. Analyse de la nature ou tableau de lUniverse et des corps organise. Palermo. [ Links ]
Rioja, E. 1923. Estudio sistemático de las especies ibéricas del Suborden Sabelliformia. Trab. Mus. Nal. Hist. Nat. Madrid, Ser. Zool., 48: 1-144. [ Links ]
Rioja, E. 1931. Estudio de los poliquetos de la Península Ibérica. Mem. Acad. Cienc. Exact. Fís. Nat. (Madrid), ser. Cienc. Nat. 2: 1-471. [ Links ]
Rioja, E. 1946. Estudios anelidológicos, 14. Observaciones sobre algunos poliquetos de las costas del Golfo de México. An. Inst. Biol. UNAM 17: 193-203. [ Links ]
Rioja, E. 1959. Estudios anelidológicos, 22. Datos para el conocimiento de la fauna de anélidos poliquetos de las costas orientales de México. An. Inst. Biol. UNAM 29: 219-301. [ Links ]
Risso, A. 1826. Histoire naturelle de principales productions de lEurope meridionale et particulièrement de celles des environs de Nice et des Alpes Maritimes. Paris, F.G. Levrault Libr., 4: 1-439. [ Links ]
Salazar-Vallejo, S.I. 1989. Enrique Rioja y su contribución al estudio de los poliquetos (Annelida: Polychaeta) en México. Brenesia 30: 39-65. [ Links ]
Salazar-Vallejo, S.I. 1996. Lista de especies y bibliografía de los poliquetos (Polychaeta) del Gran Caribe. An. Inst. Biol., ser. Zool., UNAM 67: 11-50. [ Links ]
Savigny, J.L. 1820. Système des Annelides principalement de celles des côtes de LÉgypte et de la Syrie, offrant les caractères tant distinctifs que naturels des Ordres, Familles et Genres, avec la description des espèces, p. 3-128 In Description de LÉgypte, Histoire Naturelle, 1. [ Links ]
Schmarda, L.K. 1861. Neue wirbellose Thiere beobachtet und gesammelt auf einer Reise um die Erde 1853 bis 1857. 1. Turbellarian, Rotatorien und Anneliden. 2: 1-164. [ Links ]
ten Hove, H.A. 1970. Serpulinae (Polychaeta) from the Caribbean: I. The genus Spirobranchus. Stud. Fauna Curaçao Caribb. Isl. 32: 1-57. [ Links ]
ten Hove, H.A. 1975. Serpulinae (Polychaeta) from the Caribbean: III. The genus Pseudovermilia. Stud. Fauna Curaçao Caribb. Isl. 47: 46-110. [ Links ]
ten Hove, H.A. 1989. Serpulinae (Polychaeta) from the Caribbean: IV. Pseudovermilia madracicola sp.n., a symbiont of corals. Stud. Honour Dr. Pieter Wagenaar Hummelinck 123: 135-144. [ Links ]
ten Hove, H.A. 1994. Serpulidae (Annelida: Polychaeta) from the Seychelles and Almirante Islands, p. 107-116. In van der Land, J. (ed.), Oceanic Reef of the Seychelles. Cruise Reports Neth. Indian Ocean Program, II, Nat. Hist. Mus. Leiden. [ Links ]
ten Hove, H.A. & M.J. Jansen-Jacobs. 1984. A revision of the genus Crucigera (Polychaeta; Serpulidae); a proposed methodical approach of serpulids, with special reference to variation in Serpula and Hydroides, p. 143-180. In Proc. 1st Polychaete Conf., Sydney. [ Links ]
ten Hove, H.A. & F.J.A. Pantus. 1985. Distinguishing the genera Apomatus Philippi, 1844 and Protula Risso, 1826 (Polychaeta: Serpulidae). A Further plea for a methodical approach to serpulid taxonomy. Zool. Meded. 59: 419-437. [ Links ]
ten Hove, H.A. & G. San Martín. 1995. Serpulidae (Polychaeta) procedentes de la I Expedición Cubano-Española a la Isla de la Juventud y Archipiélago de los Canarreos (Cuba). Stud. Nat. Hist. Carib. Region. 72: 13-24. [ Links ]
ten Hove, H. A. & R.S. Smith. 1990. A redescription of Ditrupa gracillima Grube 1878 (Polychaeta, Serpulidae) from the Indo-Pacific, with a discussion of the genus. Rec. Aust. Mus. 42: 101-118. [ Links ]
ten Hove, H.A. & J.C.A. Weerdenburg. 1978. A generic revision of the brackish-water serpulid Ficopomatus Southern 1921 (Polychaeta: Serpulinae), including Mercierella Fauvel 1923, Sphaeropomatus Treadwell 1934, Mercierellopsis Rioja 1945 and Neopomatus Pillai 1960. Biol. Bull. 154: 96-120. [ Links ]
ten Hove, H.A. & P.S. Wolf. 1984. Family Serpulidae Johnston, 1865, p. 55.1-55.34. In J.M. Uebelacker & P.G. Johnson (eds). Taxonomic Guide to the Polychaetes of the Northern Gulf of Mexico, Barry A. Vittor, Mobile, 7 vols. [ Links ]
Treadwell, A.L. 1929. New species of polychaetous annelids in the collections of the American Museum of Natural History, from Porto Rico, Florida, Lower California, and British Somaliland. Amer. Mus. Novit. 392: 1-13. [ Links ]
Treadwell, A.L. 1936. Polychaetous annelids from the vicinity of Nonsuch Island, Bermuda. Zoologica 21: 49-68. [ Links ]
Uchida, H. 1978. Serpulid tube worms (Polychaeta, Sedentaria) from Japan with the systematic review of the group. Bull. Mar. Park Res. Stat. Sabiura 2: 1-98. [ Links ]
Vannini, E. 1975. Cicli reproduttivi nei policheti Salmacina dysteri e Salmacina incrustans. Pubbl. Staz. Zool. Napoli 39: 335-346. [ Links ]
Zibrowius, H.W. 1969. Review of some little known genera of Serpulidae (Annelida: Polychaeta). Smithson. Contr. Zool. 42: 1-22. [ Links ]
Zibrowius, H.W. 1970. Contribution a l'ètude des Serpulidae (Polychaeta Sedentaria) du Brésil. Bol. Inst. Oceanogr. S. Paulo 19: 1-32. [ Links ]
Zibrowius, H.W. 1973. Serpulidae (Annelida Polychaeta) des côtes Ouest de lAfrique et des archipels voisins. Ann. Mus. R. Afrique Centrale, Belgique ser. 8, Sci. Zool. 208: 1-93. [ Links ]
Zibrowius, H.W. 1983. Chitinopoma arndti n. sp., an incubating bathyal serpulid polychaete from Saint-Paul island, Southern Indian Ocean. Tethys 11: 21-24. [ Links ]
Datos de los materiales recolectados. En corchetes está la clave de los sitios indicados en la Fig. 1 y 2.
AMNH 1996: Julia Cove, Bahía Guanica, Puerto Rico, sobre una concha 23 VI 1915, R.W. Miner. [54]
AMNH 2396: Gurnets Rock, Bermuda, 10 m, 19 VIII 1931. [52]
AVE3: Playa Aventuras, DIF, QR4, roca mixto, 21 III 1992, SISV. [13]
AVE4: Playa Aventuras, DIF, fondo arenoso y coralino, los corales (Acropora spp.) estaban cubiertos de sedimento y algas filamentosas, 1.5 m, 28 II 1999, SISV y JRBZ. [13]
BA1: Bahía Ascención, entre Thalassia, 28 I 1986. [15]
BUE1: Buenavista, 0.85 m, 27 IX 1996. [17]
CAZ1: Cazones, 7 X 1981, S. Salcedo Martínez (UANL). [30]
CHA13-14: La Manteca, 10 km frente a Champotón, fondo arenoso con pastos (Syringodium, Thalassia) y parches rocosos, 6 m, 16 II 1999, SISV y JRBZ. [36]
CHE1: Bahía de Chetumal, muelle de Chetumal, 3 XI 1990, SISV. [22]
CHE2: Bahía de Chetumal, muelle de Chetumal, 3 XI 1990, S. Jiménez. [22]
CHE3: Bahía de Chetumal, Luis Echeverría, 21 IX 1996, L.F. Carrera-Parra, N.E. González y SISV. [23]
CHK1: Chankanaab, Isla Cozumel, 2 IV 1992, SISV. [27]
CON1: Isla Contoy, raspado en los pilotes del muelle de madera, incrustantes, 0-3 m, 21 II 1999, SISV y JRBZ. [5]
CON2: Isla Contoy, campamento norte, arribazón de pastos, conchas y corales, 22 II 1999, SISV y JRBZ. [5]
CON3: Isla Contoy, playa Camping, fondo mixto, con grandes moluscos (Strombus sp.) y esponjas, 0.5-1 m, 22 II 1999, SISV y JRBZ. [5]
CON6: Isla Contoy, playa Camping, intermareal, 23 II 1999, SISV y JRBZ. [5]
COZ1: El Cedral, Cozumel, fondo mixto, con rocas, algas y pastos, 1.5-2 m, 24 II 1999, SISV y JRBZ. [29]
COZ2b: Playa Corona, Cozumel, fondo rocoso, con corales de fuego (Millepora complanata) y corales cerebro (Diploria sp.), 1-2 m, 25 II 1999, SISV y JRBZ. [28]
COZ3: Playa Azul, Cozumel, parte expuesta del muelle de cemento, tapizado de corales de fuego (Millepora alcicornis, M. complanata y M. squarrosa?), 0-2 m, 26 II 1999, SISV y JRBZ. [24]
CU4: Cayo Juan García, Golfo de Batabanó (Cuba), 5 m, III 1988 (IOC). [44]
CU5: Cayo Diego Pérez, Golfo de Batabanó (Cuba), 15 m, 20 VII 1988 (IOC). [44]
CU7: Arrecife de Punta del Este, Isla de la Juventud, Golfo de Batabanó (Cuba), 5 m, 16 III 1978 (IOC). [43]
CU8: Cayo Cantiles, Golfo de Batabanó (Cuba), zona de embate de la meseta arrecifal, 2 m, 15 VIII 1988 (IOC). [46]
CU9: Bahía Jururú, Provincia de Holguín, NE de Cuba, E-9, canal de entrada de la bahía, IV 1988 (IOC). [48]
CU10: Cayo Juan García, Golfo de Batabanó (Cuba), arrecife, 10 m, VII 1988 (IOC). [44]
CU11: N de cayo Rosario, golfo de Batabanó (Cuba), E-8, arrastre, 3.5 m, 30 V 1985 (IOC). [45]
ECH1: Isla Mujeres, epifauna de las espinas de Eucidaris tribuloides, 2 m, V 1962, J.L. Cifuentes. [7]
ECH3: Frente al ojo de agua, Puerto Morelos, epifauna de las espinas de E. tribuloides, 2 m, 17 II 1995, F. Laguarda, F. Solís, M. Torres, R. Ramírez. [10]
EL2771: Cozumel, 3 km al S de bahía. Caleta (20º25.97N, 87º02.08W), 18 VIII 1990. [26]
EL2774: Cayo Norte, Banco Chinchorro, (18º45.63N, 87º15.84W), 60 m, 20 VIII 1990. [19]
EL2777: SE de Banco Chinchorro (18º26.02N, 87º18.82W), 230 m, 21 VIII 1990. [20]
EL2779: S de Banco Chinchorro, sobre concha de ermitaño diogénido, 4 m, 22 VIII 1990, E. Escobar. [21]
EL2780: N de cayo Blackford, Banco Chinchorro (18º30.94N, 87º26.61W), 43 m, 22 VIII 1990. [21]
EL2792: E de Isla Mujeres, ca. 13 km de la isla (21º14N, 86º36W), 130 m, 28 VIII 1990. [6]
ESP13: Punta Allen, en esponja (Clathria calla), 2-3 m, 24 IV 1992. [14]
ESP14: Boca Paila, en esponja (Agelas dispar) con un crustáceo, 2-3 m, 24 IV 1992. [14]
F5: Cabo Catoche, rastra camaronera, 6 m, 12 X 1983, V. Solís y N. Méndez. [4]
IE1: Isla de Enmedio, UV25, en coral, 8 m, 6 VI 1992 (UV). [33]
IE2: Isla de Enmedio, UV24, en coral vivo (A. cervicornis), 3 VII 1992 (UV). [33]
IP1: Isla Pérez (=Cayo Alacranes), 7 VII 1977, U. Garza (UANL). [41]
LAG1: Trinchera de Nictilic, 1.5-2 km frente a la bocana de Ría Lagartos, fondo mixto con pastos (Syringodium, Thalassia), algas (Caulerpa prolifera), esponjas y algunos corales aislados (Solenastrea bournoni), 2-4 m, 18 II 1999, SISV y JRBZ. [2]
LAR1: Larios, Tecolutla, en ostión, 4 VIII 1985, SISV y J.C. Stivalet. [31]
MAJ1: Majagual, E23, 1 X 1996, SISV y N.E. González. [16]
NC1: Laguna Nichupté, 18 X 1987, J.J. Oliva. [8]
NC2: Laguna Nichupté, E5M671, 28 X 1987. S. Jiménez y J.J. Oliva. [8]
NC3: Laguna Nichupté, canal de entrada norte, 200 m de SEMARNAP (Cancún), fondo rocoso con algas (Acetabularia, Halimeda opuntia y Laurencia sp.) y esponjas, 0.1-2 m, 20 II 1999, SISV y JRBZ. [8]
NIZ1: Punta Nizuc, Roca 4 y 6, 31 VIII 1997, SISV, L.F. Carrera-Parra y M.A. Ruiz-Zárate. [9]
NIZ2: Punta Nizuc, Roca 7, 8 y 9, 1 IX 1997, SISV, L.F. Carrera-Parra y M.A. Ruiz-Zárate. [9]
NIZ3: Punta Nizuc, fondo rocoso y coralino, 2 m, 20 II 1999, SISV y JRBZ. [9]
PO75004: Sonda de Campeche, IMCA IV, est. 41, (19º43.1N, 91º45.8W), 42.4 m, 3 X 1989 (ICML-UNAM). [37]
PO75005a: Cayo Alacranes (=Isla Pérez), B/O Justo Sierra, DIN II, est. 1, draga, 37 m, 28 X 1990 (ICML-UNAM). [41]
PO75005b: Cayo Arenas, B/O Justo Sierra, DIN II, buceo, 4 m, 3 XI 1990 (ICML-UNAM). [38]
PO75006: Cayo Alacranes (=Isla Pérez), B/O Justo Sierra, DIN II, est. 1, draga, 37 m, 28 X 1990 (ICML-UNAM). [41]
SAM1: Punta Sam, QR6, rocas expuestas, 1 IV 1992. [8]
SC: Sonda de Campeche, Crucero BOA, est. 1, 78 m, 2 VIII 1996, F. Vázquez. [56]
SF1: San Felipe, cerca de una bocana artificial de Ría Lagartos, fondo mixto, con algunos corales pastos (Syringodium) y algas (Caulerpa prolifera y C. pinnata), 2 m, 19 II 1999, SISV y JRBZ. [1]
SJU1: San Juan de Ulúa, UV55, pared con incrustantes (vermétidos, ostras y ascidias), 1-1.5 m, 11 VIII 1996, M.A. Muro (UV). [32]
XAH1: Xahuayxol, Roca 52, 1 VI 1997, SISV y L.F. Carrera-Parra. [17]
XAH2: Xahuayxol, Roca 53, 1 VI 1997, SISV y L.F. Carrera-Parra. [17]
XAH3: Xahuayxol, 0.7 m?, 28 IX 1996. [17]
XAM1: Xamach, 28 II 1986, E. Donath. [13]
XCA3: Xcacel, QR8, rocas, 3 IV 1992, SISV. [12]
XCAY1: Xcayal, bolsa 115, 4 XI 1990, S. Jiménez. [18]
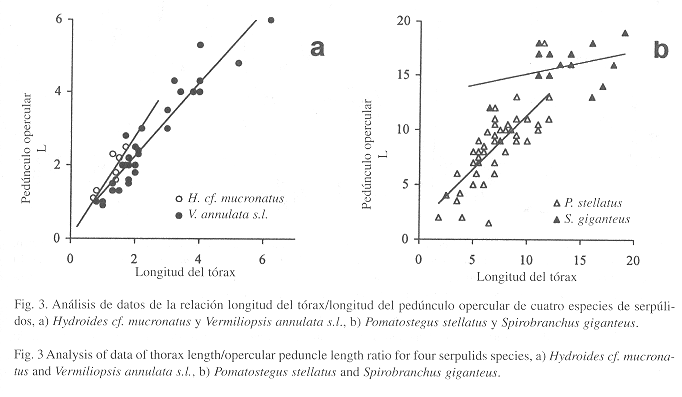
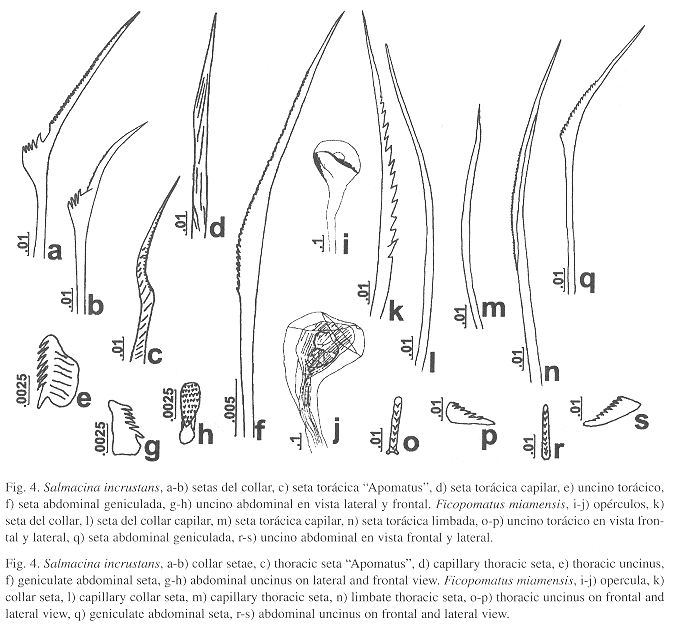
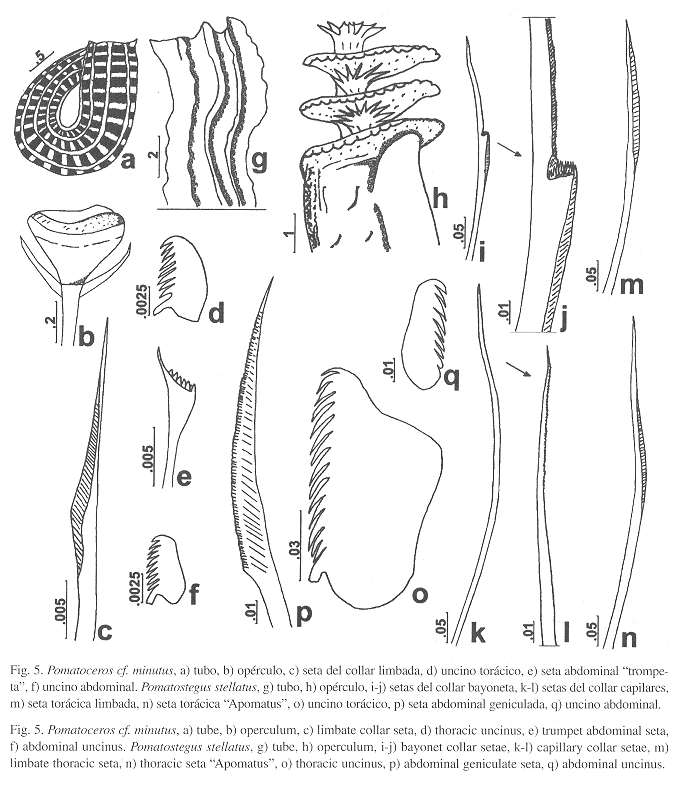
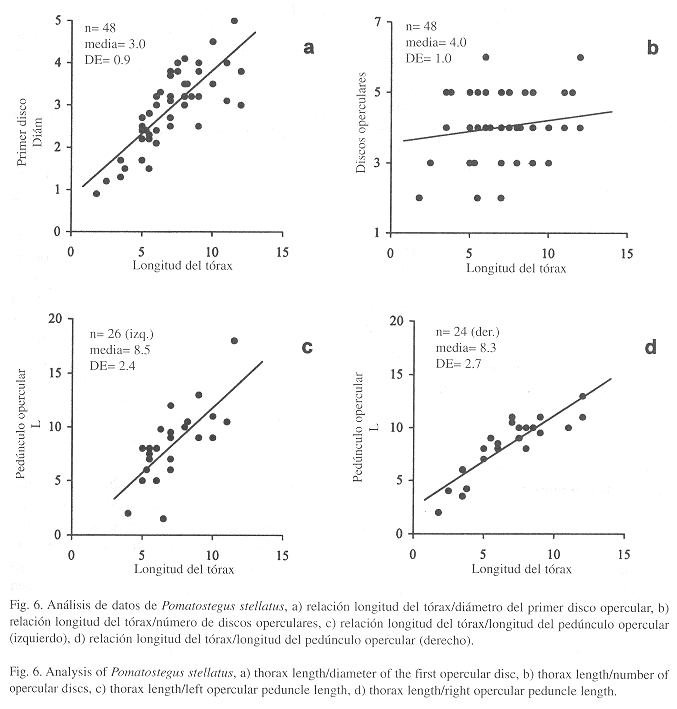
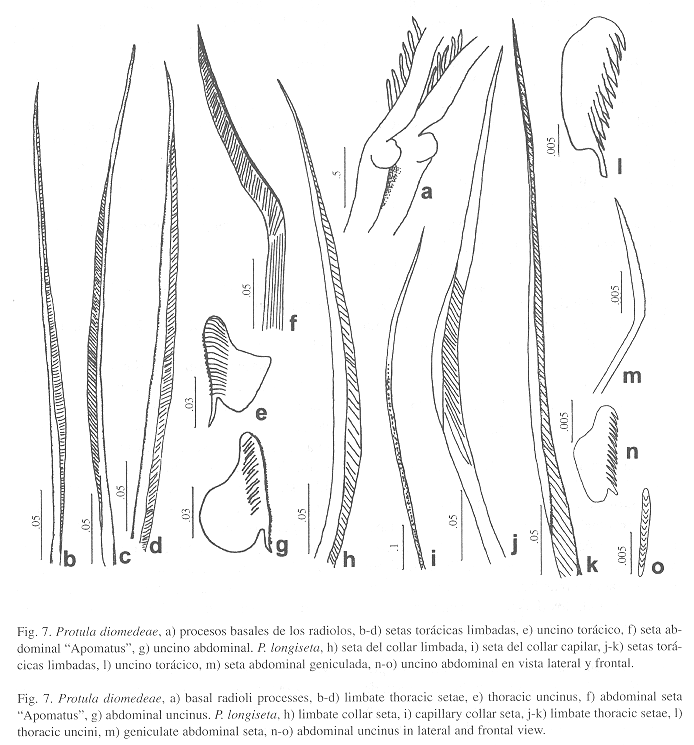
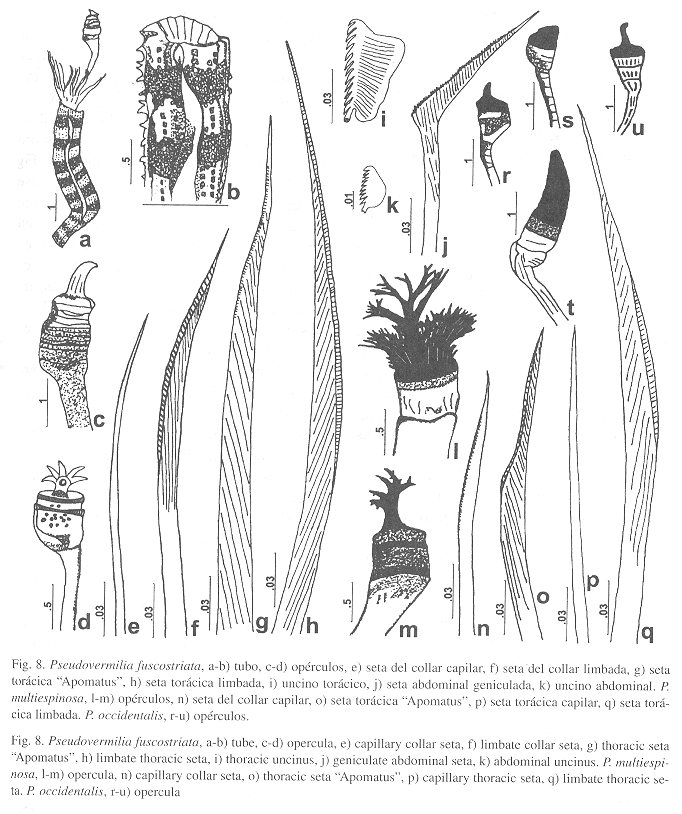


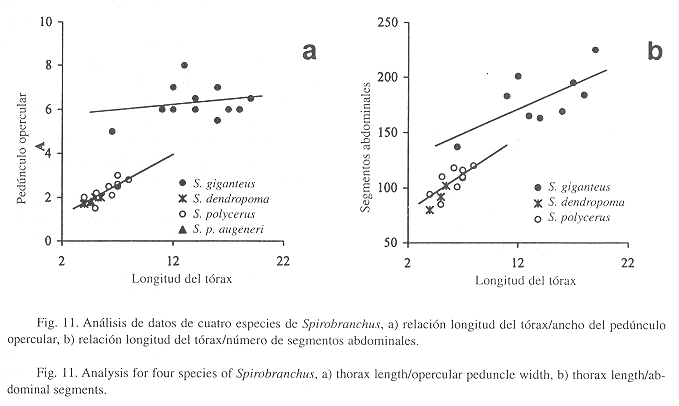
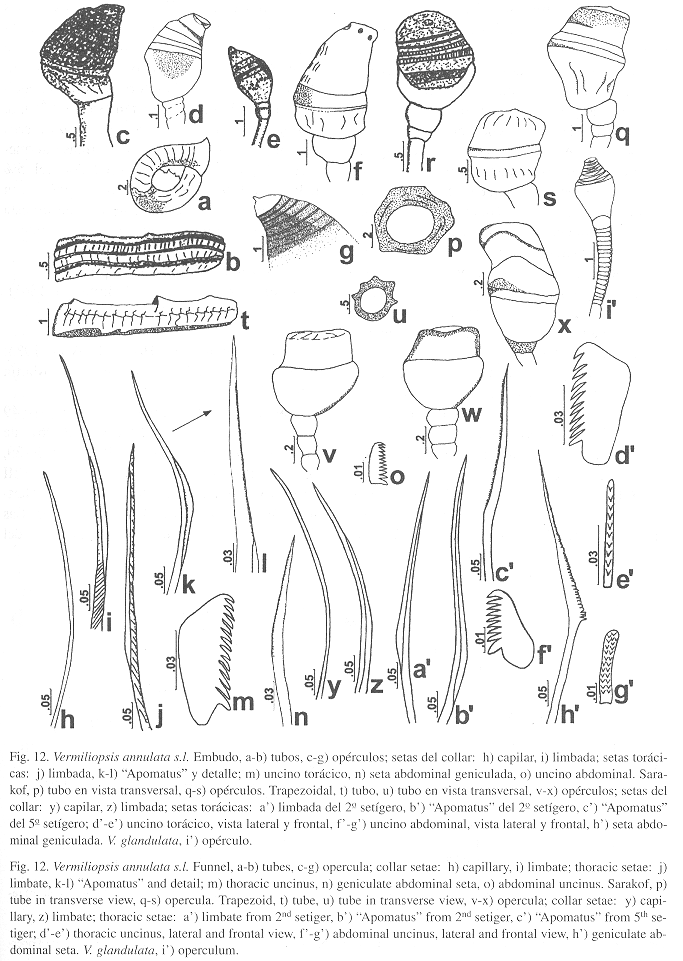
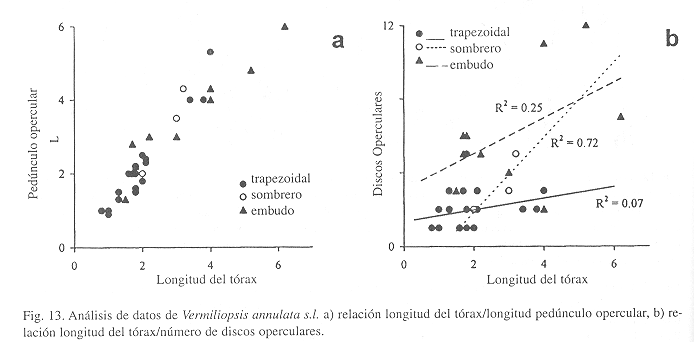
1 El Colegio de la Frontera Sur, Unidad Chetumal. Dep. de Ecología Acuática. Apdo. Postal 424, Chetumal, C.P. 77000, QROO, México, Fax (983)204-47, rolando@ecosur-qroo.mx

