










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.47 n.3 San José Sep. 1999
de la costa nororiental de Venezuela
Lilia J. Ruiz Rogelia Figueroa M. Antulio Prieto A. 1
Recibido 24-VIII-1998. Corregido 10-III-1999. Aceptado 18-III-1999.
Abstract
Reproductive biology was studied in Lactophrys quadricornis, from samples collected monthly from May 1990 to April 1991 in the east coast of Venezuela. Sex ratio was 1:1; half the population matured at total lenght of 198 mm for males and 222 mm for females. Minimal sizes for maturity was 160 and 170 mm of L.T in males and females, respectively. The Gonadosomatic Index (IG) and gonadal maturity stages sugest year around reproduction with two peaks: from June through September, and from January through February. Weight, ovary lenght and frequency of females in stages IV and V, are the evidence. Asynchronous development of the ovocytes with diameters between 0.399 to 0.930 mm, indicates that spawning is fractionate. Mean fecundity was 112 536 ± 57 047.97 eggs, and increases with lenght and weight of the fish and weight and length of the ovaries.
Key words
Lactophrys quadricornis, fish, reproduction, Venezuela.
Lactophrys quadricornis es una especie de pez de la familia Ostraciidae que se caracteriza por presentar el cuerpo corto, cuboide y estar encerrado en un caparazón constituido por escamas de láminas hexagonales firmemente unidas entre sí, excepto en el pedúnculo caudal (Cervigón 1966). Habita en fondos someros, coralinos, rocosos o arenosos, hasta cerca de 80 m de profundidad. Las tallas máximas reportadas para esta especie oscilan entre 400 y 450 mm de longitud total. Su distribución geográfica comprende ambos lados del Atlántico; en el Atlántico occidental desde Bermudas y Massachusetts, EEUU, todo el Golfo de México hasta Río de Janeiro (Fisher 1978).
Esta especie se consume en fresco y se considera como un excelente alimento; constituye uno de los platos típicos en la Isla de Margarita, ya que su carne es de buena calidad (Cervigón 1966).
En Venezuela no se tiene información sobre la biología de esta especie, a excepción de los trabajos de Cervigón (1966, 1996) sobre taxonomía. Debido a esto y a la importancia del recurso como fuente de alimento local, se justifica un estudio sobre aspectos reproductivos de L. quadricornis.
Materiales y Métodos
El estudio se basó en ejemplares de Lactophrys quadricornis, de ambos sexos, y diferentes tallas, los cuales fueron obtenidos de las capturas realizadas mensualmente, por la flota de barcos con sede en Cumaná, desde mayo de 1990 hasta abril de 1991, en el área pesquera comprendida entre la zona adyacente a la Isla de Margarita y el Estado Sucre, Venezuela (10º 30´ N - 62º 30´ W y 10º 50´ N - 64º 30´ W). A cada ejemplar se le determinó la longitud total (LT) y estándar (L. est) con precisión de 1 mm y el peso húmedo con precisión de 0.1 g.
El sexo se determinó mediante observación directa de las gónadas y los estadios de madurez sexual se establecieron siguiendo el criterio de Laevastu (1972); posteriormente se estableció una escala para la especie, de acuerdo a las características observadas. Además, se determinó el peso y la longitud promedio de cada gónada. La talla de madurez del 50% de la población se determinó para cada sexo, de acuerdo al criterio de Pitt (1966).
El ciclo reproductivo y tipo de desove se estudió mediante el Indice Gonadosomático (IG), según el criterio de Kaiser (1973) y la variación mensual por un análisis de varianza y una prueba a posteriori. El IG se relacionó con el peso total del pez por estadio de madurez, para constatar la no dependencia entre ambas variables por estadio y así darle una mayor validez al índice (De Vlaming et al. 1982, Isaac-Nahum y Vazzoler 1987).Además, se analizó la variación mensual de las frecuencias relativas de los diferentes estadios de maduración, tanto en hembras como en machos, los promedios mensuales del peso y longitud de los ovarios; y el diámetro de los óvulos por mes.
La fecundidad fue estudiada en 30 hembras maduras (estadio IV y V), de tallas comprendidas entre 194 y 279 mm de LT y entre 141.2 y 361.9 g de peso, seleccionadas de cada uno de los meses de muestreo, tomando una porción de la parte media de uno de los lóbulos de las gónadas; para estimar el número de óvulos, se utilizó el procedimiento descrito por Joseph (1963). El número de óvulos y el promedio de los diámetros de aproximadamente 100 óvulos por ejemplar, se utilizaron para calcular el índice de fecundidad (IF), respecto a la longitud total (IF/LT) y al peso del cuerpo (IF/PC). Las relaciones entre fecundidad y longitud y peso total del pez, así como de la fecundidad con el peso y longitud de la gónada, se analizaron según los criterios de Holden y Raitt (1975).
Resultado
Se analizaron 306 ejemplares de Lactophrys quadricornis. De éstos 167 eran hembras con tallas comprendidas entre 168 y 363 mm de L.T y peso entre 101.1 y 624.5 g; 139 machos con tallas entre 166 y 376 mm de L.T. y peso entre 102.8 a 684.2 g.
Proporción sexual: La proporción sexual total fue 1.20: 1.00, lo que indica que no se aleja de lo esperado 1: 1 (P>0.05). No obstante la proporción se mostró a favor de las hembras en los meses de julio, septiembre, enero y febrero; y a favor de los machos solo en el mes de agosto (Cuadro 1).
| Meses | | | |
| Mayo 90 | | | |
| Junio | | | |
| Julio | | | |
| Agosto | | | |
| Septiembre | | | |
| Octubre | | | |
| Noviembre | | | |
| Diciembre | | | |
| Enero 91 | | | |
| Febrero | | | |
| Marzo | | | |
| Abril | | | |
| Total | | | |
X2 = Ji- cuadrado; ns = P> 0.05; * = P< 0.05 y ** = P < 0.01. X2 = chi-square
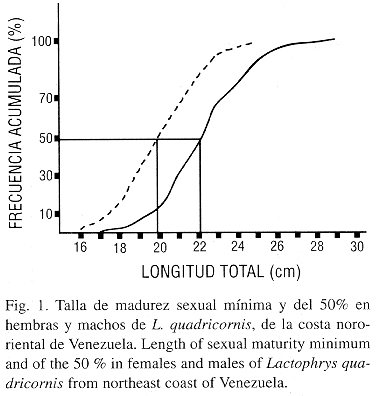
Talla mínima de madurez y del 50% de la población: La talla a la cual el 50 % de la población de L. quadricornis se encontró sexualmente madura fue 198 mm de LT, para los machos y 222 mm de LT, para las hembras. Las tallas mínimas de madurez encontradas fueron 160 y 170 mm de LT para machos y hembras, respectivamente (Fig. 1).
Indice Gonadosomático (IG): Los valores mensuales del IG en las hembras variaron considerablemente durante el año (1.29 a 8.67), lo que se puso de manifiesto en el análisis de varianza (Fs = 5.7063; P < 0.001), siendo más elevados en los meses de junio-septiembre de 1990, con una ligera disminución en julio y agosto, y enero- febrero de 1991. Los valores más bajos se presentaron de octubre a diciembre de 1990 y de marzo hasta abril de 1991 y en mayo de 1990 (Fig. 2). La prueba a posteriori de Duncan indicó la presencia de dos grupos (Cuadro 2); uno que incluye los meses de mayor actividad gonadal que va de junio a septiembre y de enero a febrero, el segundo grupo incluye los meses de octubre a diciembre, marzo y mayo, donde la especie probablemente haya incurrido en períodos de desove o en épocas de descanso.
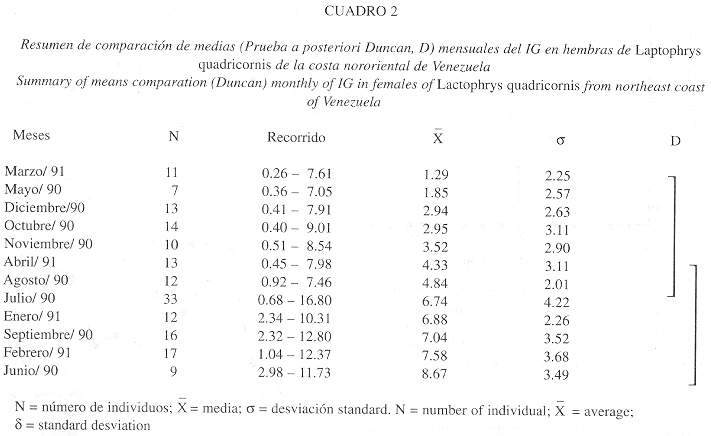
En los machos los valores del IG fueron considerablemente más bajos (0.15 – 0.32) que los obtenidos en las hembras (Fig. 2); sin embargo, se observó la misma tendencia en ambos sexos. Los picos de máxima actividad reproductiva de los machos coinciden con los de las hembras en los meses de junio y febrero, de igual manera la época de menor actividad en diciembre, marzo y abril.
Peso y longitud de los ovarios: Los valores promedios mensuales del peso de los ovarios de L. quadricornis oscilaron entre 1.96 y 26.34 g; los más altos se presentaron en los meses de junio, julio, septiembre, enero y febrero, y los más bajos en agosto, octubre-diciembre y en marzo-abril y mayo. La misma tendencia se presentó con la longitud de los ovarios (Fig. 2), observándose una relación directa entre las dos variables con el IG (P<0.001; r = 0.968 y P<0.001; r = 0.889, respectivamente).
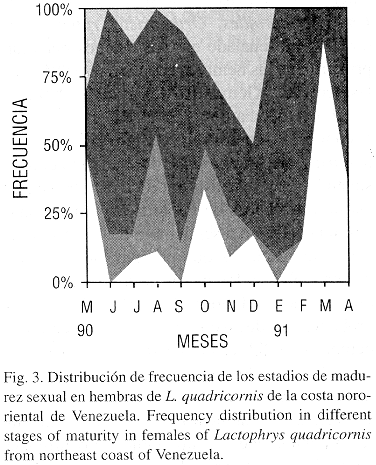
Estadios de madurez sexual: En L. quadricornis se establecieron seis estadios de madurez sexual para las hembras (Cuadro 3) y tres para los machos (Cuadro 4). El análisis de los estadios de madurez sexual reveló la existencia de pocas hembras juveniles, solo tres durante el muestreo. Las hembras en estadio II fueron abundantes en los meses de mayo, octubre y marzo; en estadio III predominaron en agosto; en estadio IV y V de madurez sexual se encontraron en mayor porcentaje en los meses de junio, julio, septiembre, enero y febrero; las hembras desovadas (VI), predominaron en mayo, noviembre, y diciembre (Fig. 3).
CUADRO 3
Características macroscópicas de las gónadas, asociadas a los estadios de madurez sexual, en hembras de Lactophrys quadricornis de la costa nororiental de Venezuela. Macroscopic appearance of gonads, associated to stages of sexual maturity in females of Lactophrys quadricornis from northeast coast of Venezuela.
| | |
| I: Inmaduro | Ovarios cilíndricos, incoloros a grises. |
| II: En vías de maduración | Ovarios de color rosado pálido, trenslúcidos, sin vasos sanguíneos ni óvulos visibles a simple vista. |
| III: Madurando | Ovarios de color amarillo pálido con vasos sanguíneos y óvulos visibles a simple vista. |
| IV: Maduro | Ovarios de color amarillo intenso, con numerosos vasos sanguíneos y óvulos maduros y transparentes. |
| V: Grávido | Ovarios de color anaranjado intenso, con fuerte irrigación, ocupan casi toda la cavidad abdominal. |
| VI: Desovado | Ovarios de color marrón, sanguinolentos, flácidos y con algunos óvulos opacos. |
Características macroscópicas de las gónadas, asociadas a los estadios de madurez sexual, en machos de Lactophrys quadricornis de la costa nororiental de Venezuela.
Macroscopic appearance of gonads, associated to stages of sexual maturity in males
of Lactophrys quadricornis from northeast coast of Venezuela.
| Estadio | |
| I: Inmaduro | Testículos transparentes y filiformes. |
| II: Maduro | Testículos de color blanco lechoso a crema, más gruesos y algo arrollados, con irrigación sanguínea. |
| III: Desovado | Testículos de color crema con zonas pardo-marrones, algo flácidos y con mucha irrigación sanguínea. |
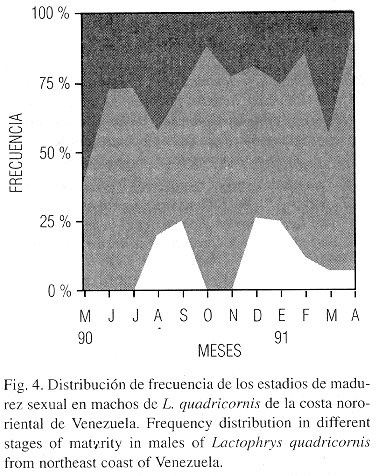
Los machos juveniles (estadio I) abundaron en mayo, agosto y marzo; los maduros (estadio II) fueron abundantes en junio - julio, octubre – noviembre, febrero y abril; los ejemplares desovados (III) se presentaron con mayor frecuencia en mayo, agosto y marzo (Fig. 4). Los meses donde los machos maduros presentaron mayor frecuencia coincidieron con los valores más altos del IG (Fig. 3).
Diámetro de los óvulos: El análisis de la distribución de frecuencia porcentual del diámetro de los óvulos, indicó valores entre 0.399 a 0.932 mm, y se encontraron en un mismo ovario de 5 a 8 clases de diámetro de óvulos. La frecuencia más elevada correspondió al grupo de óvulos con diámetro igual o mayor a 0.666 mm. Este grupo se tomó como próximo a desovar. El diámetro promedio de los óvulos de los ovarios analizados por mes osciló entre 0.630 y 0.782 mm, con valores promedios más elevados en los meses de junio, septiembre y febrero.
Fecundidad: El número de óvulos en gónadas maduras osciló entre 30 597 y 256 429, con un promedio de 112 536 ± 57 047.97. El Indice de fecundidad (IF) con relación al peso del pez varió de 194.39 a 902.60 óvulos/g con un valor promedio de 462.23 ± 177.96. El IF respecto a la longitud total varió entre 160.19 y 986.27 óvulos/mm con un promedio de 478.84 ± 214.72. Las relaciones del número de óvulos con la longitud y peso del pez fueron positivas y significativas (P<0.01), al igual que las relaciones de la fecundidad con el peso y longitud de los ovarios, y el diámetro de los óvulos (Cuadro 5). Esto indica que la fecundidad depende del la longitud y el peso del cuerpo del pez, como también del peso, longitud de los ovarios y del diámetro de los óvulos.
Relación entre la fecundidad (F) y la longitud total (LT), peso (P), peso y longitud promedio
de las gónadas (Pg y Lg) y diámetro promedio de los óvulos (DO), en ejemplares hembras de
Laptophrys quadricornis de la costa nororiental de Venezuela.
Relationships between fecundity (F) and total length (LT), weigth (P), weight and mean length of gonads
(Ps and Lg), and mean diameter eggs in females of Lactophrys quadricornis from northeast coast of
Venezuela.
| | | |
| Log F = -3.125 + 3.438 Log LT | | |
| Log F = -2.026 + 1.256 Log P | | |
| Log F = -3.986 + 0.856 Log Pg | | |
| Log F = 0.506 + 2.496 Log Lg | | |
| F = -594 389.17 + 966 DO | | |
N = número de ejemplares; r = coeficiente de correlación; ** = P < 0.01).
N = number of individual; r = correlation coefficient.
Discusión
Se estudia por primera vez el ciclo reproductivo de una especie de Ostraciidae del Atlántico, Lactophrys quadricornis. La proporción sexual total fue de 1:1; sin embargo, en los meses de julio, septiembre, enero y febrero, el número de hembras fue superior al de los machos, coincidiendo con los meses de mayor actividad reproductiva, representada por los valores más altos del IG. Esto puede estar relacionado con el hecho de que en muchas especies las hembras se congregan con fines reproductivos. Por otra parte, la proporción sexual en peces puede variar de especie a especie, entre poblaciones y de un año a otro, dentro de una misma especie (Nikolsky 1963); además, puede variar con el período reproductivo y con los grupos de tallas analizadas (Cousseau y Christiansen 1971).
La talla de primera madurez de L. quadricornis fue mayor en hembras que en machos, lo que indica que los machos alcanzan la madurez sexual primero que las hembras. Este parámetro puede variar dentro de una misma población pues en él intervienen factores hereditarios y la misma fisiología del organismo con relación a sus características genéticas y factores ambientales (Nikolsky 1963). Así, en otras especies de peces en las costas de Venezuela, tales como Haemulon sciurus, Cynoscion jamaicensis, Umbrina coroides, Orthopristis ruber, entre otras, se ha señalado que los machos maduran primero que las hembras (Urreiztieta 1985, Colassaco 1987, Gómez 1990, Guzmán 1990).
La presencia de hembras en estadios IV y V en todos los muestreos permite inferir que L. quadricornis se reproduce durante todo el año; sin embargo, el análisis del IG indica la presencia de dos épocas de máxima actividad gonadal; una de junio a septiembre de 1990, con ligeras fluctuaciones que podrían indicar desoves parciales en los meses de julio y agosto, y otra de enero a febrero de 1991; y dos épocas probables de desove que van de septiembre a octubre y de febrero a marzo. Este índice es a menudo utilizado para establecer el ciclo reproductivo en peces, así los incrementos progresivos reflejan un aumento en el grado de madurez, mientras que su caída indica la ocurrencia de un desove, observandose coincidencia de la máxima actividad reproductiva con los valores altos del índice; mientras que los valores mínimos se asocian con épocas de descanso (Kaiser 1973; Htun-Han 1978). El IG es un indicador de actividad gonadal cuando no varia en función del peso del pez, pero cambia con los estadios de madurez; si se cumple esta condición, como ocurrió con la especie estudiada, el índice puede ser un buen indicador de períodos de desove (De Vlaming et al. 1982, Erickson et al. 1985, Cayre y Laloé 1986, West 1990).
Otros parámetros relacionados con las gónadas, como el peso y la longitud, diámetro de los óvulos y estadios de madurez gonadal, son considerados indicadores adecuados y confiables para determinar épocas de reproducción en peces (Isaac-Nahum y Vazzoler 1987). En L. quadricornis se registró una alta correlación entre el peso y la longitud de los ovarios con el IG (P<0.001). Así mismo si relacionamos la frecuencia de los estadios de madurez IV y V, en los meses donde se presentaron en mayor porcentaje, con el IG, peso y longitud de los ovarios, se puede inferir que los períodos de junio a septiembre de 1990 y de enero a febrero de 1991, son épocas de máxima actividad reproductiva; la mayor frecuencia de hembras desovadas (VI) coincide con las épocas de menor actividad reproductiva o descanso de la especie. Vazzoler y Agostinho (1991a en Vazzoler 1996) analizaron el comportamiento de los valores del IG por estadios de madurez de 62 especies de teleosteos y constataron que el índice presenta un intervalo de variación definido para cada estadio, en donde los valores más altos del IG coinciden con la época de máxima actividad reproductiva (estadios IV y V), este mismo comportamiento se observó en L. quadricornis.
La presencia de óvulos de diferentes diámetros, indica desarrollo asincrónico de los óvulos, lo que hace suponer que esta especie efectúa más de un desove al año. Desoves parciales se han señalado para especies marinas del oriente de Venezuela, como Haemulon aurolineatum, H. steindachneris, Cynoscion jamaicensis, Nicholsina usta, entre otras (Kossowski 1985, Rodríguez 1985, Colassaco 1987, Márquez 1987). Desoves fraccionados y períodos prolongados de desove son característicos de especies de peces tropicales y subtropicales, no sólo como una adaptación a las fluctuaciones en la disponibilidad de alimento, sino para la preservación de las especies bajo condiciones desfavorables al desove, por lo que la tasa reproductiva se vería menos afectada que en las especies con desove total y donde la abundancia de individuos podría disminuir con el tiempo (Nikolsky 1963).
La fecundidad en L. quadricornis puede considerarse alta al igual que las relaciones de este parámetro con el peso y la longitud del pez, si se les compara con lo indicado para otras especies de peces tropicales cuyas tallas promedios son semejantes (Kossowski 1985, Rodríguez 1985, Colassaco 1987). La fecundidad es un factor importante en la explotación racional de una población íctica, y se emplea para calcular el potencial reproductivo y la supervivencia (Holden y Raitt 1975). Por otra parte, el número de óvulos y el diámetro de los mismos varía según la especie; además, puede estar relacionado con la mortalidad debida a fluctuaciones ambientales, y al cuidado parental de los huevos y alevines (Margalef 1977). Vazzoler (1996) señaló que tanto la fecundidad como el diámetro de los ovocitos maduros pueden presentar variaciones intraespecíficas, entre períodos reproductivos sucesivos, individuos de la misma talla en un mismo período y entre localidades. Micropogonioas furnieri, en las costas de Brasil, presentó variaciones latitudinales en la fecundidad, siendos más alta en la costa norte, donde la presión ambiental es más elevada (Vazzoler 1991).
Concluimos que Lactophrys quadricornis se reproduce durante todo el año, con dos períodos de máxima actividad reproductiva que coinciden con épocas de surgencias en la región nororiental de Venezuela, y que el índice gonadosomático, la longitud y peso de las gónadas constituyen buenos indicadores de períodos de madurez y desove de la especie.
Resumen
Se estudió la biología reproductiva en Lactophrys quadricornis, de muestras colectadas mensualmente desde mayo 1990 a abril 1991, en el oriente de Venezuela. La proporción sexual no se alejó de la esperada 1:1. La mitad de la población de machos y hembras maduraron a 200 y 222 mm de longitud total (LT), respectivamente. Las tallas mínimas de madurez fueron 160 y 170 mm de L.T en machos y hembras, respectivamente. De acuerdo a la observación macroscópica de las gónadas se establecieron seis estadios de madurez en hembras y tres en machos. El Indice Gonadosomático (IG) y los estadios de madurez gonadal sugieren que la especie se reproduce durante todo el año con dos picos de máxima actividad reproductiva: de junio a septiembre, y de enero a febrero. El peso, la longitud de los ovarios y la frecuencia porcentual de hembras en estadios IV y V son evidencias de esa actividad. El desarrollo asincrónico de los ovocitos con diámetros entre 0.399 a 0.930 mm, indica que los desoves son fraccionados. Los valores de la fecundidad oscilaron entre 30 597 a 256 492 óvulos con un promedio de 112 536 ± 57 047.97, observandose un incremento de la fecundidad con la talla y el peso del pez y con el peso y la longitud de los ovarios.
Referencias
Cayre, P. & F. Laloé. 1986. Review of the gonad index (GI) and an introduction to the concept of it "critical value" Aplication to the spipjack tuna Katsuwonus pelamis in the Atlantic Ocean. Mar. Biol. 90: 345-351. [ Links ]
Cervigón, F. 1966. Los Peces Marinos de Venezuela. Tomo II. Fundación La Salle Cienc. Nat., Monogr. 12, p. 847-850. [ Links ]
Cervigón, F. 1996. Los Peces Marinos de Venezuela Vol. IV. Ex Libris, Caracas p. 107-113. [ Links ]
Colassaco, A. 1987. Ciclo reproductivo de la Tonquicha, Cynoscion jamaicensis (Vaillant y Bocourt, 1883), Pisces: Sciaenidae, en la plataforma Unare - Píritu, Edo. Anzoátegui. Tesis de Licenciatura, Universidad de Oriente, Cumaná, Venezuela. [ Links ]
Cousseau, M. & H. Christiansen. 1971. La reproducción de la merluza y su relación con otros aspectos biológicos de la especie. Bol. Inst. Biol. Mar. Argentina 20: 44-74. [ Links ]
De Vlaming, V., G. Grossman & F. Chapman. 1982 On the use of the Gonosomatic Index. Comp. Bioch. Physiol. 73 A: 31- 39. [ Links ]
Erickson, D. L., J. Hightower & G. Grossman. 1985. The relative Gonadal Index: An Alternative index for quantification of reproductive condition. Comp.Biochem. Physiol. 81a : 117 –120. [ Links ]
Fisher, W. (Ed.). 1978. Species identification sheets for fishery purposes. Western Central Atlantic. (Fishing Area 31) Vol. I. Roma, F.A.O. 459 p. [ Links ]
Gómez, G. 1990. Algunos aspectos biológicos y pesqueros de la petota, Umbrinacoroides (Cuvier 1830). Pisces: Sciaenidae, del área norte del Estado Sucre, Venezuela. Tesis de Maestría, Universidad de Oriente, I.O.V. Cumaná, Venezuela. [ Links ]
Guzman, G. 1990. Aspectos biológicos y pesqueros del corocoro, Orthopristis ruber (Cuvier, 1830) (Pisces: Haemulidae), en áreas de la isla de Margarita y Costa Norte del Edo. Sucre, Venezuela. Tesis de Maestría, Universidad de Oriente, I.O.V. Cumaná, Venezuela. [ Links ]
Holden, M. & D. Raitt. 1975. Manual of Fishery Sciences. Part. II. Methods of resources investigation and application. F.A.O. Fishery Technical Papers 115(1): 34-43. [ Links ]
Htun – Han, M. 1978. The reproductive biology of the dab Limanda limanda(L) in the North Sea: Gonadosomatic Index, hepatosomatic index and condition factor. J. Fish. Biol. 13: 369 – 378. [ Links ]
Isaac- Nahum, V. & A. E. A. de M. Vazzoler. 1987. Biologia reproductiva de Micropogonias furnieri (Desmarest, 1823) (Teleostei: Sciaenidae), 2. Relacao Gonadossomatic, comprimento e peso dos ovarios como indicadores do periodo de desova. Bol. Inst. Oceanogr. , S. Paulo 35: 123 – 134. [ Links ]
Joseph, J. 1963. Fecundity of Tunnus albacares and Katsuwonus pelamis from the Eastern Pacific Ocean. Bull. Inter. Amer. Trop. Tuna Comm. 8: 257 – 277. [ Links ]
Kaiser, C. E. 1973. Gonadal maturation and fecundity of horse mackarel, Trachurus murphi of the coast of Chili. Trans. Amer. Fish. Soc. 102: 101 – 108. [ Links ]
Kossowski, A. 1985. Reproducción del cují, Haemulon aurolineatum (Cuvier, 1829) de la Isla de Margarita. Tesis de Licenciatura, Universidad de Oriente, Cumaná, Venezuela. [ Links ]
Laevastu, T. 1972. Manual de Métodos de Biología Pesquera. Acribia. F.A.O. Zaragoza, España 233 p. [ Links ]
Margalef, R. 1977. Ecología. Omega, Barcelona, España 951 p. [ Links ]
Marquéz, Y. 1987. Estudio sobre la actividad reproductiva de Nicholsina usta (Valenciennes, 1839) Pisces: Scaridae. Tesis de Licenciatura, Universidad de Oriente, Cumaná, Venezuela. [ Links ]
Nikolsky, G. 1963. The Ecology of Fishes. Academy, Nueva York 352 p. [ Links ]
Pitt, T. 1966. Sexual maturity and spawning of the american plaice, Hippoglossoides platessoides (Fabricius) from New Foundland and Grand Bank ares. J. Fish. Res. Board Can. 23: 651 – 672. [ Links ]
Rodríguez, I. 1985. Reproducción del chere-chere, Haemulon steindachneris (Jordan y Gilbert, 1882) (Perciformes: Pomadasyidae). Tesis de Licenciatura, Universidad de Oriente, Cumaná, Venezuela. [ Links ]
Urreiztieta, I. 1985. XXXV Convención Anual de la Asociación Venezolana para el Avance de la Ciencia. Acta Científica Venezolana Vol 36, suplemento Nº 1, Venezuela 240 p. [ Links ]
Vazzoler, A. E. A. de M. 1991. Sintase de conhocimentos sobre la biología da corvina, Micropogonias furnieri (Desmarest, 1823) da costa do Brasil. Rev. Atlantica, Rio Grande 13: 55 – 74. [ Links ]
Vazzoler, A. E. A. de M. 1996. Biologia de Reproducao de Peixes Teleosteos: Teoía e Practica. Maringá, EDUEM / SBI / CNPq / Nupelia, Sao Paulo 169 p. [ Links ]
West, G. 1990. Methods of assessing ovarian development in fishes: a review. Aust. J. Mar. Freshwater Res. 41: 199 – 222. [ Links ]
1 Departamento de Biología, Escuela de Ciencias, Universidad de Oriente. Aptdo. Postal 245. Cumaná, Venezuela. Correo electrónico: liruiz@sucre.udo.edu.ve

