










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.46 n.4 San José Dec. 1998
Abstract
Age and growth of Oreochromis niloticus from Lagoon of Coatetelco, Morelos State, Mexico were studied from January through December, 1993. Scales of 318 specimens were collected. Modal length at capture was 10.5-11.5 cm standard length. Scales rings were formed during December. Back-calculated lengths-at-age showed no significant differences by sex. Four check marks were recorded. According to the growth curve parameters for population, the fish grow at a low rate (k=0.07) until they achieve a size (L*) of 29.19 cm. Length-frequency analysis (Bhattacharya's Gaussian component determination procedure) do not differ significantly (t-student, p<0.05), from the scale reading.
Key words
Oreochromis niloticus, age, growth, length-frequency, scales, Mexico.
The family Cichlidae, with about 700 species, is the predominant perch-like fish occuring naturally in Africa and Madagascar, and also in the Central and South America Tropical Zone ( Fryer & Iles 1972, Morales 1991). They have been introduced into a large number of tropical and sub-tropical countries around the world in the last four or five decades, either accidentally or deliberately. The tilapias are important food fish in aquaculture and they are normally reared in open ponds in the tropics (Soderberg 1990). The Nile Tilapia Oreochromis niloticus (Linnaeus, 1757), is considered a suitable species for rural fish farming, taking into account its fast growth rate, adaptability to a wide range of culture conditions and its high capacity to transform farm waste into protein. The tilapias has proven to bring both social and economic benefits to the rural people living close to aquatic systems under, which show exploitation and management fisheries (Arredondo 1983).
The annual production of tilapias in Mexico was 92 891 t in 1994 (Anonymous 1995). This figure represent about 7.9 % of total production from fish caught in lakes and from fish farming. In 1976 O. niloticus was introduced to Mexico from Panama (Arredondo-Figueroa and Tejeda-Salinas 1989), including releases into Coatetelco Lagoon in 1990. Studies in this aquatic system are very important, because they are the basis to determine the rate at which a rational exploitation should be carry out (Oduleye 1982).
Various aspects of the biology of Cichlids of commercial importance have been studied by Garrod (1959), Fryer and Iles (1972), Fagade (1974), Bernal (1984), Guerra and Peña (1985), Moreau (1985), Arredondo-Figueroa and Guzmán-Arroyo (1986), Hernández (1987), Alejo et al. (1989), Yamaguchi et al. (1990), Jaramillo and Sánchez (1991), Legendre and Albaret (1991), Seyoum and Kornfield (1992), Morales (1991), Morales (1992), Zhang and Runham (1992), Guzmán (1994) and Abdulrahaman (1996-1997). There is however no published account of the biology of these species from Coatetelco Lagoon, Mexico.
The present study deals with the determination of age and growth rate in O. niloticus from Coatetelco Lagoon, Morelos State, Mexico, by means of scales reading and length frequency distribution analysis.
Materials and Methods
Samples from the commercial catch at Coatetelco Lagoon, located in Morelos State (18°45' N, 99°20' W, altitude 1 100 m), were taken approximately at monthly periods from January to December 1993. Fish were caught by static gill-net with mesh size 5.08 cm. The specimens were collected by the same fisherman in the same place and day. A total of 318 fish were analyzed: 157 females and 161 males. Fish were measured for total length (TL) and standard length (SL) to the nearest mm and weighed (total weight) to the nearest g. The sex was determined by examination of internal organs. Ten to fifteen scales were removed from the left side of the body in the region between the middle of the pectoral fin and the lateral line (Tesch 1968, Ruiz-Dura et al. 1970). They were washed, mounted on glass slides and examined by a slide projector. The rings that continued around the entire circumference of the scale were counted and the total number of these rings was used to estimate the age of each specimen. Counts for each specimen were performed twice, approximately at periods two months apart, without knowledge of the length of the specimen. The scale radius (R) was defined as the maximum distance from the focus to the distal edge of the scale. The distance from the focus to each annulus was measured along this axis. The relationship between SL and R was linear but not directly proportional, having a positive intercept on the ordinate. SL-R regressions were tested for differences in slopes between sexes using analysis of covariance (ANCOVA). Back-calculated length-at-age of individual fish were estimated from the standard length-scale radius regressions using the Fraser-Lee method (Tesch 1968). Mean weighted back-calculated lengths-at-age were tested for differences between sexes using Student's t-test. Also the length frequency distribution analysis was employed through the modification of Bhattacharya´s procedure (Pauly and Caddy 1985).
The validation of ageing and the periodicity of the annulus formation were verified through marginal growth examination and focus-to-annulus distance (mean and range). Marginal growth was examined in all scale aged during the whole sampling period. The percentages of fish having edge growth were compared by month of capture (Yamaguchi et al. 1990).
The mean values resulted from length frequency distribution analyses and those obtained by hard-part readings, they were used to estimate the growth parameters of the von Bertalanffy growth function (vBGF) which was fitted to mean weighted back-calculated lengths-at-age using a non-linear regression program available in StataCorp (1997), to obtain the nominal value for L* and k. However, the vBGF was used according to Barlow (1992). Boxplot scatter plots (Salgado-Ugarte 1992), residual analyses (Curts 1984) and Student t-test were used.
Results
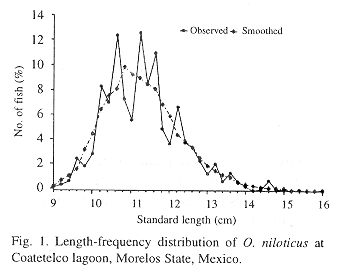
The total sample size was 318 fishes (Table1). Mean standard body length of males was 8.9 to 14.8 cm (females 9.0 to 15.8 cm) and their weight 25.7 to 106.5 g (females 25.8 to 100.6 g). The standard length-frequency distribution of tilapias in the lagoon is shown in figure 1. Modal length at capture was 10.5-11.5 cm standard length and it observed a bias to values more than 12.0 cm. The total length was highly correlated with standard length:
SL = -0.229 + 0.794 TL, r2 = 94.6%, n = 318.
| | | ||||||
| n | Range | Mean | S.E | Range | Mean | S.E. | |
| Males | 161 | 8.9-14.8 | 11.12 | 0.08 | 25.7 -106.5 | 57.9 | 0.57 |
| Females | 157 | 9.0-15.8 | 11.36 | 0.09 | 25.8 -100.6 | 52.6 | 0.51 |
| All fish | 318 | 8.9-15.8 | 11.24 | 0.06 | 25.7 - 106.5 | 55.2 | 0.41 |
Age determination by length-frequency distribution analyses: The length-frequency distribution analyses for pooled data were smoothed by running average over five (Pauly 1987) to clarify the distribution and to obtain more efficiently the component means. Component characterization obtained through the Bhattacharya's procedure were carried out using sexes combined data, because the mean values for males and females did not differ significantly (t-student, p<0.05). Seven size groups were estimated.
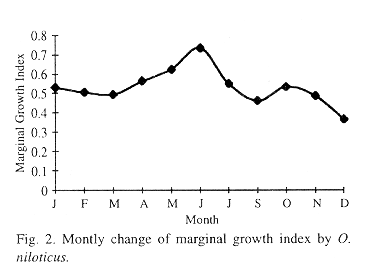
Age determination by scales: Scales were collected from 318 fish. Seventy-three percent (110 for females, 122 for males) of the scales were readable and used for study of age and growth. Check marks were distintic in all scales fields. Data indicated that two annulus were formed per year. Observation of the position of the last complete ring scales of 232 specimens, showed that most of the fish caught during December-January (71.8%) and June (63.3%), had a complete ring close to the edge. However, the marginal growth index (Fig. 2) suggested that annuli are formed between December and January. The mean focus-to-annulus distance estimated for each individual, was consistent for each annulus (Table 2). Modal age at capture was represented mainly by age two (57.32%), and a lowest percentage (1.72%) by age zero.
| | | | | | | |
| 0 | | | | | | |
| | | | | | | |
| 1 | | | | | | |
| | | | | | | |
| 2 | | | | | | |
| | | | | | | |
| 3 | | | | | | |
| | | | | | | |
| 4 | | | | | | |
| | | | | | | |
The regressions of SL (cm) on scales radius (R) were estimated separately for separated and combined sexes. The slopes of the SL-R regressions did not differ significantly between sexes (ANCOVA, P<0.05). Therefore, the body-scale relationship used for back-calculate length-at-age for combined data was:
SL = 6.0931 + 1.2617 R, n=232 fish, r2=65.2 %
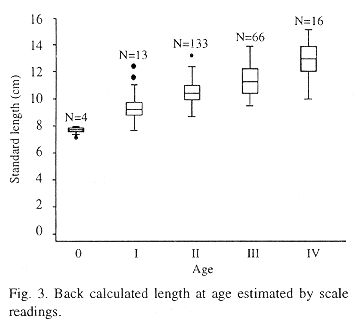
Intercepts from this regressions were used to back-calculate length-at-age which are summarized in Table 3. The greatest growth in length occurred in the first year of life, and growth increment declined steadily after the first year reaching a low figure around 9.81 mm SL.
| | No of fish | | ||||
| | | | | | | |
| | | | | | | |
| | | | | | | |
| | | | | | | |
| | | | | | | |
| | | | | | | |
| | | | | | | |
| | | | | | | |
| | | | | | | |
| | | | | | | |
Length at age estimated hard part readings by means of boxplot (Fig. 3) allowed to establish at the median as the central tendency value for being resistent. In this case, both the mean and median were similar value and mean value was used. Therefore, the mean values age from I to IV of the hard part readings were similar with the mean values age from I to IV of the length frequency analysis (t-student, p<0.05) through Bhattacharya´s procedure.
| I | II | III | IV | V | VI | VII | |
| Total | 9.54 | 10.88 | 11.88 | 12.77 | 13.55 | 14.67 | 15.56 |
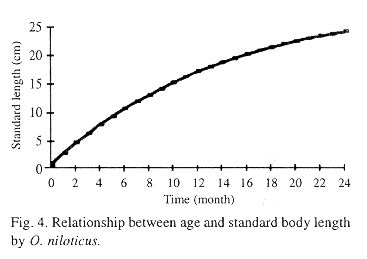
L(Li,t) = 29.19 - (28.19 * exp(-0.07*t))
Discussion
The length-frequency distribution of O. niloticus shows a young looking population, because they were caught with static gill-net size 5.08 cm. We obtained small specimens (from 8.8 to 16.0 cm standard length), in comparison with Oreochromis aureus reported at others reservoirs in Mexico, caught with gill-net size from 7.62 to 10.16 cm, which ranging from 15.0 to 35.0 cm total length (Bernal 1984, Morales 1992, Guzmán 1994), because fishermen did not allow to use other gill-net size. We are convinced that specimens with a larger average total body length could be obtained in the Lagoon of Coatetelco, because this aquatic system has a high nutritional potential and it is considered as eutrophic (Granados 1990). On the other hand, if fishermen do not change their attitude respect the use of other fishing gear, regardless the lagoon is characterized as an eutrophic system, the fishing effort and the capture could decreasing in time. Therefore, it is necessary to realize research with different fishing gear to determine which one requires less energy per unit of effort, selectivity of catch as to the size and age, and increased survival of the catch.
The formation of rings in the scales of tilapia were observed in December-January (winter), which corresponds with the minimum value of marginal growth.This event may be attributed to lower temperatures (21 °C) prevailing in the lagoon during this season. However, the percentage of fish caught during December-January and June had a complete ring close to the edge, and therefore the growth rings found (mainly June) on the scales were probably due to growth interruption by the gonad maturation and the spawning activity (although spawn all the year around), because O. niloticus is a mouth brooder and it is possible that in the females, this breeding habit would prevent feeding and thus lead to growth ring formation. Yamaguchi et al. (1990) found that the formation of rings occurred every January when they used the minimum value of marginal growth.
Respect to the formation of marks in the scales, Fagade (1974) in Tilapia malanotheron of Lagos Lagoon found that the complete growth rings on the opercular bone are formed probably by the spawning activity during the height of the rainy season and incomplete growth rings formed during the dry season. Garrod (1959) has shown that in the mouth breeding T. esculenta caught in L. Victoria, the growth rings are formed in the scales as a result of spawning, and the same pattern was reported by Alejo et al. (1989) in O. mossambicus, and Guzmán (1994) in O. aureus from Mexico. It is probably that the frequency of spawning may be influenced by the abundance and season availability of food and by other environmental factors in different localities (Babiker and Ibrahim 1979, Oduleye 1982).
Our data show that two growth band are formed, and this together with the high correlation found between standard length and scales radius, demonstrate the validity of using scales for estimating the age and past growth history of tilapia, as presented by Yamaguchi et al. (1990).
Age determination by the two methods used in this work, allowed to compare the average values attained to estimate the growth rate of tilapia, because the scale size could be different in fishes of the same length and also within the specific place where they are taken (Carlander 1987). Therefore, the length-frequency distribution for this species during the study was based on mainly by the number of fishermen that realized this activity (40 individuals), and the gill-net mesh-size that fishermen society used.
The mean values from I to IV age group by the two methods used were similar, and most of the total sample (133 specimens) was represented by fish with two growth checks. In another fraction the fish of age 0 (4 specimens) and IV (16 specimens) were less represented, due to gill-net mesh size, in the range of 8.0-16.0 cm. Making thus, the market price in Coatetelco is low.
With reference to growth parameters, Morales (1991) for O. aureus and Yamaguchi et al. (1990) for O. niloticus reported similar values of body length (25.0 cm) at two years old. Moreau (1985) who worked with T. rendalli found 28.25 cm body length with 0.06 growth coefficient. It is probably that the similarity with our results could be due to the temperature of water (24-30 °C), even though the eutrophic grade and catch of fish are very different. Studies in several other fish species of Oreochromis (Guerra and Peña 1985, Hernández 1987, Jaramillo and Sánchez 1991) from temporary ponds in Morelos State during one year, had been carried out and similar results in standard body length were obtained. This rapid linear growth in the first year of life may represent an evolved adaptation to reduce its vulnerability to the environment.
Flores (1994) found that O. niloticus in ponds at Mexico City, attains 20.7 cm asymptotic length standard (L*) with 0.2038 growth coefficient. Although this asymptotic value is smaller than the one obtained in this study, the mean values used by him were similar to those used to estimate the growth parameters of the von Bertalanffy growth function (VBGF).
Compared to the standard body length obtained in this study, Fryer and Iles (1972) mentioned that T. nilotica in Lake Albert reaches a length of 50 cm, and does not breed until it has attained a length of about 28 cm. However, this same species in the lagoon called Buhuka (600 m and 2 m deep), adjacent lake Albert, did not exceed a length of 17 cm, and were breeding at a length of 10 cm. This population was considered with "dwarf" or pygmy Tilapia. Also, cited that under natural conditions the growth achieved in the first year of life by several species of Tilapia is from about 9 to 12 cm. This represents adaptations to adverse conditions for growth as a result of selection for low growth rates and relatively early maturation.
This last observation becomes important because the Coatetelco lagoon presented a collapse in 1986 by a crack at ground, and the water was absorbed. In 1990, after solving the problem, the system was flooded and O. niloticus were introduced. Therefore, factors such as temperature, pH, total hardness, depth level, nutrients and productivity could be very different and the growth rate of the fish would be affected.
It is important to mention that the information presented in this work require additional analyses, besides the use of other bone structures (opercular bones, otolith, vertebrae) to compare results and to obtain better estimates of growth parameters.
Resumen
Se realizaron estudios de enero a diciembre de 1993 para conocer la edad y crecimiento de Oreochromis niloticus obtenida de las capturas comerciales de la laguna de Coatetelco, Morelos, Mexico. Se colectaron escamas de 318 peces. La moda de longitud patrón que se obtuvo en la captura fue de 10.5-11.5 cm. Se encontró que la formación de los anillos se realiza en Diciembre. Asimismo, no se detectaron diferencias significativas entre las hembras y los machos para las longitudes retrocalculadas para cada edad. En las escamas se registraron cuatro marcas. Se encontró que de acuerdo a los parametros de la ecuación de crecimiento, los peces tienen baja tasa de crecimiento (k=0.07) y alcanzan un tamaño adecuado (L* =29.19 cm). Los resultados obtenidos por medio del análisis de distribución de frecuencias no difieren significativamente (t-student, p<0.05) de los obtenidos por medio de la lectura de la estructura ósea (escamas).
Acknowledgements
I am indebted to Mercedes Garduño Paredes, Juan Avelar Esquivel and Bertha Peña Mendoza for collecting, processing and providing measurements from O. niloticus. Reviews and suggestions of an early version of this manuscript by I.H. Salgado-Ugarte, Manuel Arroyo Guzmán and Marco Antonio Barrera Camacho were very helpful. The author acknowledges Benny Weiss Staider for his help with the manuscript. This work was supported by Facultad de Estudios Superiores-Zaragoza, U.N.A.M. and Secretaría de Desarrollo Rural of Morelos State.
References
Abdulrahaman, A.K. 1996-1997. Food, size and condition of Oreochromis niloticus in Nigeria (Pisces: Cichlidae). Rev. Biol. Trop. 44/45: 655-658. [ Links ]
Alejo, P.M.C., M.E.M. Laguna & P.T. Ramírez. 1989. Estudio de algunos aspectos Biológicos de Oreochromis mossambicus (Osteichthyes: Cichlidae) en la Laguna "El Rodeo", Estado de Morelos. Tesis de Licenciatura, Escuela Nacional de Estudios Profesionales Zaragoza, Universidad Nacional Autónoma de México, México. 129 p. [ Links ]
Anonymous. 1995. Anuario Estadístico de Pesca. Dirección General de Informatica, Estadística y Documentación. Secretaría de Pesca, México. 320 p. [ Links ]
Arredondo, F.J.L. 1983. Especies Animales Acuáticas de Importancia Nutricional Introducidos en México. Inst. Nal. de Invest. sobre Recur. Bióticos, Xalapa, Veracruz, México, 8. 25 p. [ Links ]
Arredondo-Figueroa, J.L. & M. Guzmán-Arroyo. 1986. Actual situación taxonómica de las especies de la Tribu Tilapiini (Pisces: Cichlidae) introducidas en México. An. Inst. Biol. Univ. Nal. Autón. Méx. Ser. Zool. 56: 555-572. [ Links ]
Arredondo-Figueroa, J.L. & M. Tejeda-Salinas. 1989. El hueso faríngeo, una estructura útil para la identificación de especies de la tribu Tilapiini (Pisces: Cichlidae) introducidas en México. An Inst Cienc. del Mar y Limnol. Univ. Nal. Autón. Méx. 16: 59-68. [ Links ]
Babiker, M.M. & H. Ibrahim. 1979. Studies on the biology of reproduction in the cichlid Tilapia nilotica (L.): gonadal maduration and fecundity. J. Fish Biol. 14: 437-448. [ Links ]
Barlow, J. 1992. Nonlinear and logist growth in experimental populations of guppies. Ecology 73: 941-950. [ Links ]
Bernal, B.F.W. 1984. Análisis de los factores relacionados con la producción pesquera de Tilapia nilotica en la presa Adolfo López Mateos (Infiernillo). Mich-Gro. Tesis de Licenciatura, Facultad de Ciencias, Universidad Nacional Autónoma de México, México, 44 p. [ Links ]
Carlander, D.K. 1987. A history of scale age and growth studies of North American freshwater fish; p. 3-14. In C.R. Summerfelt & H.E. Gordon (eds.). Age and Growth of fish. Iowa State University, Ames, Iowa. [ Links ]
Curts, J.B. 1984. Introducción al análisis de residuos en Biología. Biótica 9: 271-278. [ Links ]
Fagade, S.O. 1974. Age determination in Tilapia melanotheron (Ruppell) in the Lagos Lagoon, Lagos, Nigeria. with a discussion on the enviromental and physiological basis of growth markings in the tropics; p. 71-77. In T.B. Bagenal (ed.). The Ageing of Fish. Unwin, Old Woking, Surrey, England. [ Links ]
Flores, M.O. 1994. Crecimiento de Oreochromis niloticus en estanques con diferente fertilización, en un clima templado. Tesis de Licenciatura, Facultad de Estudios Superiores Zaragoza, Universidad Nacional Autónoma de México, México, 56 p. [ Links ]
Fryer, G. & T.D. Iles. 1972. The Cichlid Fishes of the Great Lakes of Africa: Their Biology and Evolution. Oliver & Boyd, Edinburgh, 641 p. [ Links ]
Garrod, D.J. 1959. The growth of Tilapia esculenta Graham in Lake Victoria. Hydrobiologia 12: 268-298. [ Links ]
Granados, R.J.G. 1990. El comportamiento del zooplancton en tres ambientes acuáticos epicontinentales del estado de Morelos, México. Tesis de Maestría en Ciencias (Biología), Facultad de Ciencias, Universidad Nacional Autónoma de México, México. 64 p. [ Links ]
Guerra, H.E.A. & M.B. Peña. 1985. Estudio del crecimiento de Oreochromis hornorum (Trewavas, 1980) en relación a las condiciones Limnológicas del embalse permanente de Michapa, Edo. de Morelos. Tesis de Licenciatura, Escuela Nacional de Estudios Profesionales Zaragoza, Universidad Nacional Autónoma de México, México, 70 p. [ Links ]
Guzmán, U.A. 1994. Parámetros biológicos de tilapia (Oreochromis aureus Steindachner, 1864) de la presa Adolfo López Mateos "El Infiernillo", Michoacan Guerrero, México. Tesis de Licenciatura, Facultad de Ciencias, Universidad Nacional Autónoma de México, México. 33 p. [ Links ]
Hernández, A.J.S. 1987. Evaluación de la tasa de crecimiento y robustez de la mojarra: Oreochromis urolepis hornorum (Trewavas, 1980), carpa plateada: Hipophtalmichthys molitrix (Valenciennes) y carpa cabezona: Aristichthys nobilis (Richardson) en un sistema de policultivo extensivo en el estanque de temporal de Chavarría en el Edo. de Morelos. Tesis de Licenciatura, Escuela Nacional de Estudios Profesionales Zaragoza, Universidad Nacional Autónoma de México, México. 94 p. [ Links ]
Jaramillo, S.G. & A.V. Sánchez. 1991. Evaluación del crecimiento de la carpa barrigona (Cyprinus carpio rubrofuscus) y tilapia (Oreochromis urolepis hornorum) bajo condiciones de policultivo en el bordo de temporal "Chavarria", en el Mpio. de Coatlán del Río, Edo. de Morelos. de junio 1989 a Enero 1990. Tesis de Licenciatura, Escuela Nacional de Estudios Profesionales Zaragoza, Universidad Nacional Autónoma de México, México. 87 p. [ Links ]
Legendre, M & J.J. Albaret. 1991. Maximum observed length as an indicator of growth rate in tropical fishes. Aquaculture 94: 327-341. [ Links ]
Morales, D.A. 1991. La tilapia en México. A.G.T. México, D.F., 190 p. [ Links ]
Morales, B.E. 1992. Evaluación de algunos aspectos biológico-pesqueros de la tilapia Oreochromis aureus (Steindachner, 1864) en la presa Vicente Guerrero, Las Adjuntas, Tamaulipas. Tesis de Licenciatura, Escuela Nacional de Estudios Profesionales Iztacala, Universidad Nacional Autónoma de México, México. 28 p. [ Links ]
Moreau, J. 1985. The general von Bertalanffy growth function: possible use in the Beverton and Holt Model. Fishbyte 3: 7-10. [ Links ]
Oduleye, S.O. 1982. Growth and growth regulation in the Cichlids. Aquaculture, 27:301-306. [ Links ]
Pauly, D. & J. Caddy. 1985. A modification of Bhattacharya's method for the separation of normal distributions. FAO Fish. Circ. 781. 16 p. [ Links ]
Pauly, D. 1987. A Review of the ELEFAN System for Analysis of Length-Frequency Data in Fish and Aquatic Invertebrates: 7-34. In: Pauly, D. and G.R. Morgan (eds.). Length-based Methods in Fisheries Research. ICLARM Conf. Proc. 13. [ Links ]
Ruíz-Durá, M.F., A. Orijel & G. Rodríguez. 1970. Líneas de Crecimiento en Escamas de algunos Peces de México. Inst. Nal. de Invest. Biol. Pesq. Serie Investig. Pesq., Estudio 2, 100 p. [ Links ]
Salgado-Ugarte, I.H. 1992. El análisis Exploratorio de Datos Biológicos. Fundamentos y Aplicaciones. Marc Ediciones y Universidad Nacional Autónoma de México, 243 p. [ Links ]
Seyoum, S & I. Kornfield. 1992. Taxonomic notes on the Oreochromis niloticus subspecies-complex (Pisces: Cichlidae), with a description of a new subspecies. Can. J. Zool. 70: 2161-2165. [ Links ]
Soderberg, W.R. 1990. Temperature effects on the growth of the blue tilapia in intensive aquaculture. The Progressive Fish Cult. 52: 155-157. [ Links ]
Statacorp. 1997. Stata Statistical Software: Release 5.0. University Drive East, College Station, Texas, USA, 445 p. [ Links ]
Tesch, F.W. 1968. Age and Growth: 93-123. In: Ricker, W.E.: Methods for Assessment of Fish production in Fresh Waters. Blackwell Sci. Pub. IBP HANDBOOK No. 3. [ Links ]
Yamaguchi, Y., N. Hirayama, A. Koike & H.A. Adam. 1990. Age determination and growth of Oreochromis niloticus and Sarotherodon galilaeus in High Dam Lake, Egypt. Nippon Suisan Gakkaishi 56: 437-443. [ Links ]
Zhang, Z. & N.W. Runham. 1992. Effects of food ration and temperature level an the growth of Oreochromis niloticus (L.) and their otoliths. J. Fish Biol. 40: 341-349. [ Links ]
1 Laboratorio de Limnología, Facultad de Estudios Superiores Zaragoza, Campus II, UNAM. Batalla del 5 de Mayo esq Fuerte de Loreto, Col. Ejercito de Oriente, C.P. 09230, Mexico, D.F. Fax 7 73 01 51, correo electrónico: luis@puma2.zaragoza.unam.mx

