










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.46 n.3 San José Sep. 1998
Abstract
The distribution of continental reefs is presented along 83 km of shore within Babia Almirante and 125 km within the ChIriquí Lagoon, Bocas del Toro, Panamá. Reef community structure (diversity, cover, zonation) is described on 15 coral reefs for both regions, representing tlie diversity of hydrologicat and orographical condiiions affecting the coastal zone. The coral reefs found in Bahía Almirante are well developed down to a maximum depth of 23 m, and a diversity of hard corais of 33 species, which represents approximately a 53% of the total diversity known for Panamá.Coral cover observed for the studied region ranged between 20% and 50% depending on the depth, and a total average of 35% is estimated, nearly twice the cover informed for other regions of Panamá (San Blas), where it is presumed that the reefs are better protected from human disturbances. Vertical distribution and composition of species are similar across the area; Porites furcata dominates shallow reefs down to 2 m depth, with a cover estimated at 90%, followed by a zone dominated by Agaricía tenuifolia (40%) down to 6 m depth, followed by a zone composed mainly of A. tenuifolia (15%), Madracis mirabilis (7%), and Siderastrea siderea (5%) down to 15 rn deep, .and finishing in a deeper zone dorninated by coralline algae and the coral S. siderea. The existence of a semi-continuous Porites furcata fringing reef of ca. 22 km in length, borders the entire coastal zone of Ensenada Grande de Almirante, is informed for the first tirne for the region. Sponges, second sessile organisms in importance, are widely distributed in all reefs with a diversity of 31 species and average cover of 6%. Curiously, the highest algae coverage was found in one reef (10%) while the remaining reefs presented an average cover of 4%. Only one coral reef was found in the Chiriquí Lagoon with development limited to 3 m, a diversity of corals no higher than 7 species, and coral cover of 24% (Porites furcata). The coast of the Chiriquí Lagoon is affected by continuous sediment discharges coming from various rivers, mailnly the Cricamola, Manantí, Uyama, Guarumo, and Cauchero, whose plumes of freshwater and silt extend for many kilometers, limiting the distribution and development of coral reef communities. The vertical distribution, composition, and dominance of certain coral species in the reefs described here (e.g., Siderastrea siderea, Porites furcata, Agaricia tenuifolia, Madracis decactis, and M. mirabilis) indicate that these communities were formed adapting gradually to unfavorable conditions in water quality (lower salinity, low light penetration) which has limited a more vigorous reef development, and perhaps more diverse community. These species, in spite of heing quite resistant lo environmental changes and building healthy reefs in waters of low clarity, presently live near tbeir limits of ecophysiological tolerance, which makes them particularly vulnerable to puncluated changes in the environment. It is expected that any unplanned development project in the region rnight increase sedimentation toward the coastal zone, which could weaken all reef organisms, affecting ecosystem function.
Key words
Coral reef, biodiversity, Caribbean, reef assessment, ecology
Los arrecifes coralinos son ecosistemas altamente productivos y diversos que cumplen funciones importantes en el mantenimiento de la zona costera, beneficiando directamente a los asentamientos humanos adyacentes y a las economías de muchas naciones (Hatcher l988, 1990 Spurgeon1992) la producción de alimentos, la utilización de corales para medicamentos, y el turismo recreativo, entre otros. Valorar cada uno de estos beneficios es una labor muy difícil, pero se acepta como regla que una protección adecuada de este ecosistema repercutirá en una economía más saludable en provecho de los pobladores de la zona costera (Spurgeon 1992).Recientemente la destrucción global de arrecifes coralinos ha sido vista con preocupación por científicos y autoridades responsables del manejo de recursos naturales. Entre las causas principales de deterioro que se mencionan están la sobrepesca, la sedimentación, y el enriquecimiento (cutroficación) por nutrientes (Roberts 1993). Las consecuencias de estas actividades, además de afectar la estructura del ecosistema arrecifal, tienen un impacto económico y social en muchos países del Caribe.
Actualmente, la falta de protección de estos ecosistemas es la causa principal de deterioro indirecto en Panamá. El manejo inadecuado de la zona litoral y la deforestación de áreas adyacentes han incrementado la destrucción de todos los arrecifes a nivel local, mientras el país se prepara para un desarrollo que contempla entre otros planes, el uso recreativo de la zona costera, particularmente los arrecifes de coral. La Provincia de Bocas del Toro es considerada como una prioridad de desarrollo a nivel nacional y se esperaría que se considere un ordenamiento territorial adecuado a las necesidades locales que permita el manejo de estos recursos a largo plazo.
Los recursos marinos de la zona costera de esta región del país no han sido evaluados apropiadamente hasta el momento, siendo una necesidad imperante la implementación de estudios que describan la distribución y estado actual de estos ecosistemas. Una revisión de la literatura científica sobre los arrecifes coralinos de Bocas del Toro revela la existencia de poca información donde se describan estas comunidades.
La información más completa proviene de diversos experimentos que se realizaran para estudiar la dispersión de petróleo y sus efectos en comunidades marinas, como parte de los planes de contingencia ante el posible escenario de un derrame de petróleo en la Laguna de Chiriquí. Gundlach et al. (1985) evaluaron las comunidades costeras de esta región y dentro de estas no mencionan la existencia de arrecifes coralinos, a pesar que se observa en este trabajo que el área de posible influencia afectaría el sector norte de la Laguna de Chiriquí, que incluye el Cayo Agua y las islas Popa y Bastimentos. Posteriormente, los únicos recuentos cuantitativos sobre arrecifes, manglares y pastos marinos se dan al realizarse experimentos donde se involucran directamente a los organismos que forman estas comunidades, y para eso se seleccionaron dos cayos ubicados entre las islas Cristóbal, Popa y Bastimentos (Ballou et al. 1989a, 1989b, Dodge y Knap 1993). En estos trabajos se describen los arrecifes coralinos en tres parcelas experimentales de 900 m2 a una profundidad inferior a 1.3 m, que fueron sometidas a diversas dosis de petróleo. Se menciona que los principales corales observados fueron Porites fucata y Agaricia tenuifolia, encontrándose inicialmente una cobertura total de organismos sésiles de ca. 50% y de corales en ca.35%. Estos resultados, aunque no sirven para describir las comunidades coralinas de Bocas del Toro por lo limitado en área estudiada y profundidad, demuestran una cobertura de coral bastante alta para esos momentos.
Otros estudios mencionan, de forma general, la existencia de arrecifes a lo largo de la zona costera de Bocas del Toro (Jackson y Coates 1993) y en el Parque Nacional Marino Isla Bastimento (Mou-Sue 1993). Además, la región de Bocas del Toro se incluyó en un estudio sobre contaminación por metales realizado para centroamérica (Guzmán y Jiménez 1992). Los autores utilizaron corales y sedimentos arrecifales como indicadores de contaminación en tres arrecifes de Bocas del Toro, infdormándose de concentraciones elevadas para varios metales (Cu, Ni, Pb, Cr, Cd) acarreados por los ríos provenientes de la actividad bananera y de las actividades del oleoducto. Otros elementos (Al, Fe, Mn) parecieran indicar un elevado aporte de material terrígeno hacia la zona costera del archipiélago. Recientemente, se publicó el estudio más completo hasta la fecha para la región de Bocas del Toro, en este se contemplan aspectos de sedimentología, cartografía submarina, y se describe inicialmente las comunidades bentonicas de la Bahía Almirante (Greb et al. 1996).
Debido a lo extenso del área geográfica del archipiélago de Bocas del Toro (ca. 3 500 km2), el presente estudio es parte de una serie de cuatro publicaciones que contemplan la descripción total de los arrecifes coralinos e inventario de organismos sésiles asociados a estas comunidades en cuatro unidades geográficas arbitrariamente definidas. Este primer estudio incluye unicamente los arrecifes que bordean la zona litoral continental o de tierra firme desde el Río Changuinola hasta Punta Valiente; los arrecifes de la región insular serán descritos posteriormente, además, de la región continental entre Punta Valiente-Río Calovévora y el Escudo de Veraguas . Una última publicación analizará todos los arrecifes del área en conjunto, incorporando Sistema de Información Geográfica en el análisis y presentación de la información. El estudio comprende tres aspectos básicos para la evaluación de los arrecifes de coral: 1) distribución de los arrecifes coralinos; 2) inventario sobre la diversidad (número de especies) de corales y algunos organismos principales asociados al arrecife (plantas y animales); y 3) zonación, cobertura y abundancia relativa de corales y principales organismos sésiles en arrecifes de la región. Además, se describe cualitativamente el estado de conservación de los arrecifes y otras comunidades litorales, incluyendo las praderas de hierbas marinas, los bosques de manglar y el bosque lluvioso tropical. Esto permitirá evaluar el estado actual de las comunidades de la zona costera, con particular énfasis en los arrecifes coralinos, y proyectar así las áreas naturales vulnerables a programas de desarrollo presentes y futuros en la región. El presente estudio también servirá de línea de base a futuras investigaciones que se realizarán en la región.
Material y Métodos
Area de estudio: La Provincia de Bocas del Toro se encuentra localizada al occidente de la República de Panamá entre los 8°30' y 9°40' de latitud norte, y entre 82°56' y los 8l°8' de longitud oeste; limita por el norte con el Mar Caribe, al sur con la Provincia de Chiriquí, al este con la Provincia de Veraguas, y al oeste con la República de Costa Rica. La provincia tiene una superficie de ca. 8 917 km2 (Rodríguez et al. 1993), estimándose que el 68% de su superficie terrestre está cubierta actualmente por bosque tropical lluvioso (INRENARE 1995). El clima es altamente lluvioso, ca.287 cm, con una mayor precipitación hacía el lado sureste de la Laguna de Chiriquí (Gordon 1982), aunque dependiendo del tipo de vegetación o zona de vida la precipitación puede ser superior a los 700 cm (Rodríguez et al. 1993). Las estaciones seca y lluviosa no están claramente definidas, pero existen dos períodos de poca lluviosidad (marzo y septiembre-octubre) y dos períodos de alta Julio y diciembre). Por lo general, ocurren lluvias acompañadas de torrnentas en la costa durante las mañanas y noches en respuesta al enfriamiento de masas de aire relacionadas con la orografía local.
Geología y orografía: La Cuenca de Bocas del Toro contiene extensas secuencias del Neogeno que abarcan desde el Mioceno inferior hasta el Pleistoceno inferior, con la secuencia más antigua representada alrededor de los 20 Ma, identificada como lecho marino profundo tropical existente antes de la formación del istmo (Coates y Jackson 1998). Rocas ígneas y sedimentarias, que actualmente forman el basamento rocoso en muchas partes del archipiélago, indican actividad volcánica comprendidad entre 16 y 10 Ma. Al detenerse la actividad volcanica, transgresiones marinas subsecuentes permitieron la deposición de una secuencia sedimentaria, entre el Mioceno tardío y el Pleistoceno cercana a la costa y hacia la plataforma externa, la cual permitiría el asentamiento de una fauna marina diversa y abundante que incluiría formaciones arrecifales con edad Mioceno, Plioceno, y Pleistoceno (Coates y Jackson 1998). Depósitos del Mioceno medio encontrados en el area sugieren que durante ese período ocurrió un cambio faunístico en toda la región del Caribe, el cual incrementaría la especiación de corales formadores de arrecifes y foraminiferos bénticos (Collins el al. 1996). Cambios en el nivel del mar ocurridos durante los ultimos 9 500 años modificaron notablemente la batimetffa y topografía del archipiélago de Bocas del Toro, resultando en episódios de aislamiento y unión de las islas entre sí y con tierra firme. Se estima que el archipiélago actual se formó alrededor de 6 000 años B.P. (Summers et al.1997).
La orografía cercana a la zona litoral la conforma una cadena montañosa con altitud variable de entre 50 m y 400 m que corre paralela a la costa a una distancia inferior a 3.5 km en el sector de la Laguna de Chiriquí y a 1 km en la Bahía Almirante (IGNTG 1988). Numerosos ríos y quebradas drenan hacía ambas áreas. Debido a lo corto de estos y a la distancia angosta que existe entre la costa y las montañas, ocurre un aumento del caudal de ríos y quebradas en respuesta inmediata a las lluvias, lo cual produce una escorrentía constante hacía la zona costera. El agua proveniente de estos cauces forma una lente de agua superficial de ca.0.5 m de espesor y rica en material orgánico en suspensión afectando la bahía y la laguna, que posteriormente se mezcla a estratos inferiores que se encuentran bastante superficiales. La escorrentía media anual para la región de la Bahía Almirante podría estar alrededor de 1 600 mm y la precipitación en 3 000 mm, mientras que para la Laguna de Chirichí podría ser de 2 400 mm la escorrentía y similar la precipitación (IGNTG 1988).
Hidrografía: En la región de Bocas del Toro la plataforma continental es bastante angosta y cercana a la costa. La zona costera presenta profundidades máximas de entre 20-50 m, y esta formada por dos grandes cuerpos de agua, la Bahía Almirante y la Laguna de Chiriquí, y un archipiélago conformado por siete islas principales y decenas de cayos cubiertos de manglar (Rodríguez et al. 1993, ver recuadro en Fig. 1). El oleaje y las mareas tienen un efecto mayor afuera del archipiélago, donde las condiciones son más oceánicas. Las mareas del Caribe de Panamá pueden ser semidiurnas o diurnas (mixtas), no claramente predecibles y con una amplitud inferior a 0.5 m, que son afectadas durante la época seca cuando se producen mareas más altas que lo usual bajo la influencia de los vientos del nordeste (Glynn 1972). La principal corriente costanera que afecta a Panamá corre en dirección. Este todo el año, proveniente del Norte de Nicaragua y Costa Rica, y posiblemente tiene una mayor influencia entre los meses de junio y agosto cuando está más cercana a la costa de Bocas del Toro (DMA 1988, Greb el al. 1996). Los vientos de mayor influencia y dominantes a lo largo del año provienen del Norte y Nordeste (DMA 1988).
Sin embargo, la influencia de vientos, mareas y oleaje dentro de la Bahía Almirante y la Laguna de Chiriquí es notablemente reducida por el sistemas insular y barreras arrecifales situadas al norte del archipiélago, permitiéndose así un sistema semi-lagunar con corrientes marinas de dirección bastante variable y gobernadas por los vientos también variables durante el día. Las mareas oscilan entre 2 y 15 cm y las corrientes tienen una velocidad inferior a 40 cm/seg (Ballou et al. 1985, Gundlach el al. 1985, Greb et al. 1996)
Reconocimiento de arrecifes y muestreo:
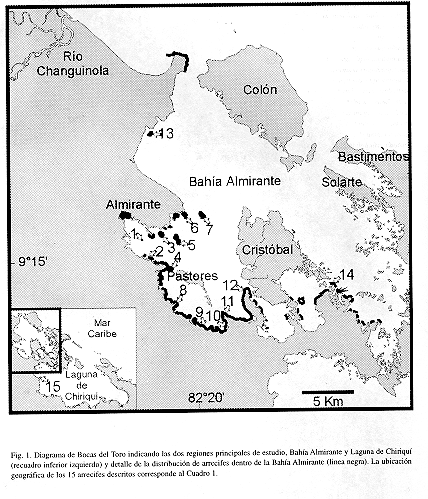
El área de estudio se dividió en dos regiones principales de tierra firme: Bahía Almirante y Laguna de Chiriquí (Fig. 1). La primera región comprende toda la zona costera desde la desembocadura del Río Changuinola hasta la punta sureste del Cerro Bruja, limitando con la lsla Loma Partida. La segunda región comprende la Laguna de Chiriquí, desde la Península de Valiente (Punta Valiente) hasta la Isla Loma Partida (lado suroeste de Isla Popa). Se hizo un reconocimiento detallado (mayo-octubre 1997) en bote de la zona costera de ambas regiones -ca.208 km- describiéndose la distribución y estado actual de los arrecifes coralinos, y a su vez, se observó las condiciones actuales de desembocaduras de ríos y quebradas, y de áreas de bosques deforestadas cercanas a la costa. Durante este recorrido se identificaron los arrecifes que serían usados posteriormente para la descripción detallada. Todos los nombres que se emplean en el presente estudio para caracterizar la localización de cada arrecife, se obtuvieron principalmente de mapas (escala 1:50 000) publicados por el Instituto Geográfico Nacional 'Tommy Guardia" (1991). La presentación en la Fig. 1 de la distribución de arrecifes coralinos se considera aproximada, debido a que no existen fotografías aéreas de la zona costera que sirvan para cartografiar adecuadamente este ecosistema.
Para cada arrecife estudiado se realizaron dos tipos de evaluaciones que permitiría la caracterización adecuada de la estructura de la comunidad arrecifal (adaptado de Sullivan y Chiappone 1992, Sullivan et al. 1994):
(1) Durante 80 min se hizo un reconocimiento completo del arrecife desde la parte más profunda hasta lo somero, anotando la composición de especies y diversidad de los organismos sésiles de los taxónes más comunes, incluidos los corales escleractínidos, octocoralarios, esponjas, algas, y otros. Además, se hizo una descripción general de la zonación de las especies de coral en el arrecife para determinar su distribución vertical. Las especies de organismos sésiles fueron clasificados usando referencias selectas para algas (Littler et al. 1989), esponjas (Zea 1987, Sefton y Webster 1986, Humann 1994a), anémonas (Sefton y Webster 1986, Humann 1994a), octocorales (Bayer 1961, Muzik 1982, Guzmán y Cortés 1985, Humann 1994b), y corales escleractinidos (Cairns 1982, Cortés y Guzmán 1985, Humann 1994b).
(2) Se escogieron al azar tres transectos de 5 metros de largo cada uno al azar y separados entre si por la misma distancia, paralelos a la costa y en cada una de las tres profundidades seleccionadas para describir cada arrecife (1 m, 3 m y 10 m), para un total de 45 m< por arrecife. La distancia de cada transecto o eficiencia de muestreo fue determinada al comparar el número de especies de coral versus área de muestreo (sensu Loya 1972), encontrándose que a la distancia de 5 m se alcanzaba un nivel de diversidad constante. En cada transecto, se estimó visualmente la cobertura de cada organismo sésil presente empleando una cuadrícula de PVC de 1 m< subdividida en 100 celdas de 100 cm < cada una.
Resultados
Bahía Almirante: Esta región se caracterizó por tener la mayor formación de arrecifes a lo largo de la costa, bajo la influencia de numerosos ríos y quebradas (Fig. 1). La influencia de la Ciudad de Almirante y sus alrededores se puede observar en los arrecifes cercanos a este sector. Las quebradas Nigua y Limón, y el Río Oeste descargan altas cantidades enormes de sedimentos en este sector, observándose el efecto de estos a varios kilómetros al sur y sureste de la Ciudad de Almirante. Los arrecifes localizados alrededor del Cayo Gerechow, al sureste de la ciudad, están completamente muertos y cubiertos por algas. Este cayo al igual que los restantes, se encontraba originalmente rodeado por un arrecife poco profundo construido por el coral ramificado Porites furcata. Actualmente el substrato del arrecife está cubierto principalmente por el alga verde Cladophora prolifera, entremezclada con parches de la anémona Stichodactyla helianthus y el gusano plumoso Sabellastarte magnifica. El coral marfil Oculina diffusa forma conjuntamente con la hierba marina Syringodium filiforme parches pequeños. Hacia el lado sureste del cayo las condiciones mejoran y se comienza a encontrar arrecifes en mejores condiciones. Es importante notar que la composición de los sedimentos en las áreas lagunares de hierbas marinas y los taludes arrecifales de la mayor parte del área de estudio, indica una alta contribución de carbonatos de origen biogénico y poco sedimento fino silicoclástico (ver Greb et al. 1996).
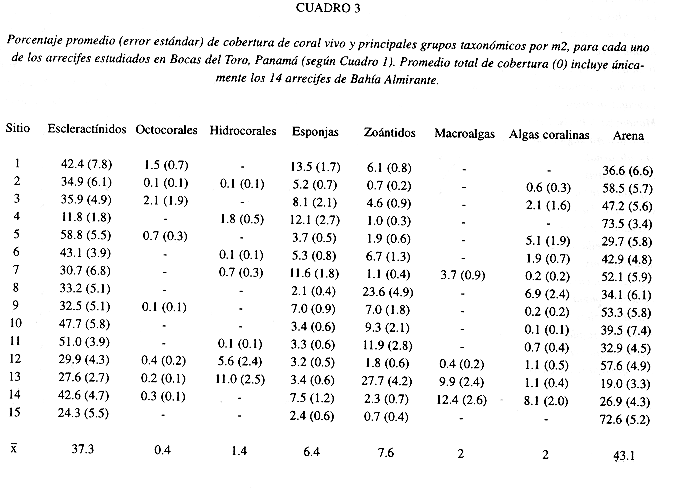
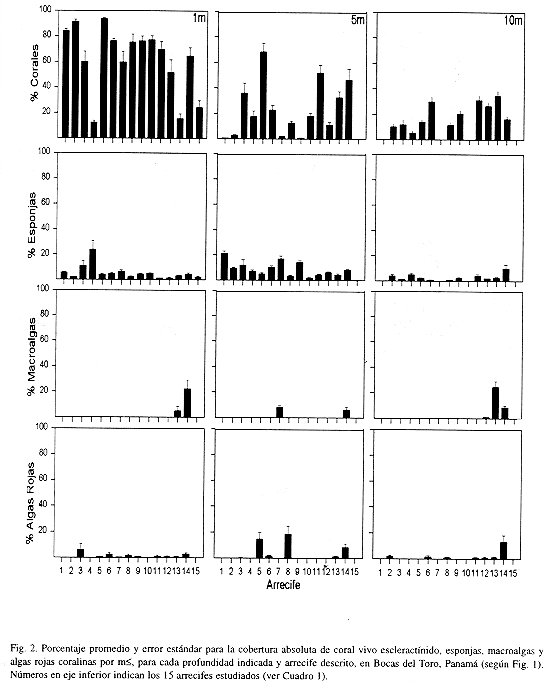
Se estudiaron en detalle 14 arrecifes coralinos distribuidos a lo largo de la costa y representativos de esta sub-región (Fig. 1, Cuadro 1). La diversidad y cobertura de coral viviente varia de acuerdo al arrecife y su profundidad de desarrollo. La diversidad de animales sésiles no presenta un patrón geográfico definido, aunque se observa una tendencia a menor diversidad en las áreas internas (suroeste) de la bahía. Se encontró una mayor diversidad de corales escleractínidos en los arrecifes 6, 13, 11 y 12 , con un número superior a 20 especies, de un total de 33 encontradas en la región de estudio (Cuadro 2). La menor diversidad se da en arrecifes poco desarrollados en cuanto a profundidad e influencia de los manglares (arrecifes 1, 4, 7, y 8). Acropora cervicornis, una importante especie constructora de arrecifes, se encontró casualmente en solo dos arrecifes (6, 1 3), mientras que A. palmata no se observó en los arrecifes. Siderastrea siderea, Porites furcata, Agaricia tenuifolia, Madracis decactis y M. mirabilis son comúnes en casi todos los arrecifes (Cuadro 2). Por lo general, las áreas arrecifales poco profundas (<3 m) presentaban coberturas de coral superiores al 20% y en su mayoría cercanas al 50% (Fig. 2). Los arrecifes 4 y 13 fueron los de menor cobertura, debido a la existencia de una barra de arena en el primero y un alto crecimiento de macroalgas en el segundo (Cuadro 3). Se encontró que la cobertura de corales disminuyó con la profundidad en la mayoría de los casos, excepto en los arrecifes 6, 9, y 11-14, donde la cobertura se mantuvo superior al 20% hasta los 10 m de profundidad (Fig. 2). Se podría decir que la cobertura de coral escleractínidos promedio fue bastante alta, oscilando entre 11% y 59% (Cuadro 3).
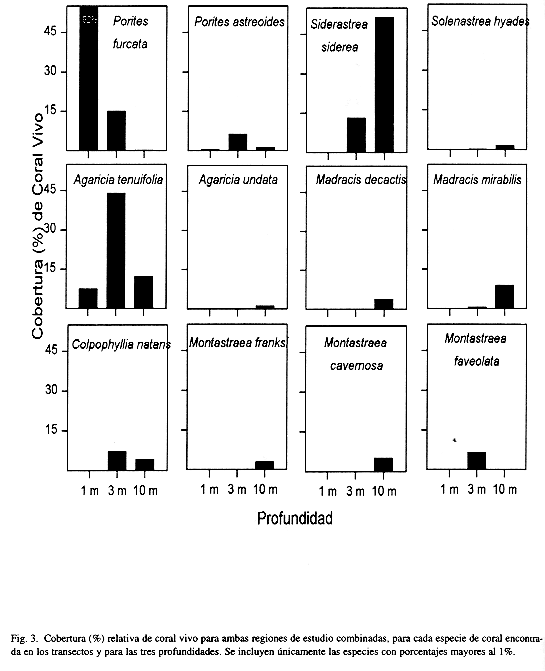
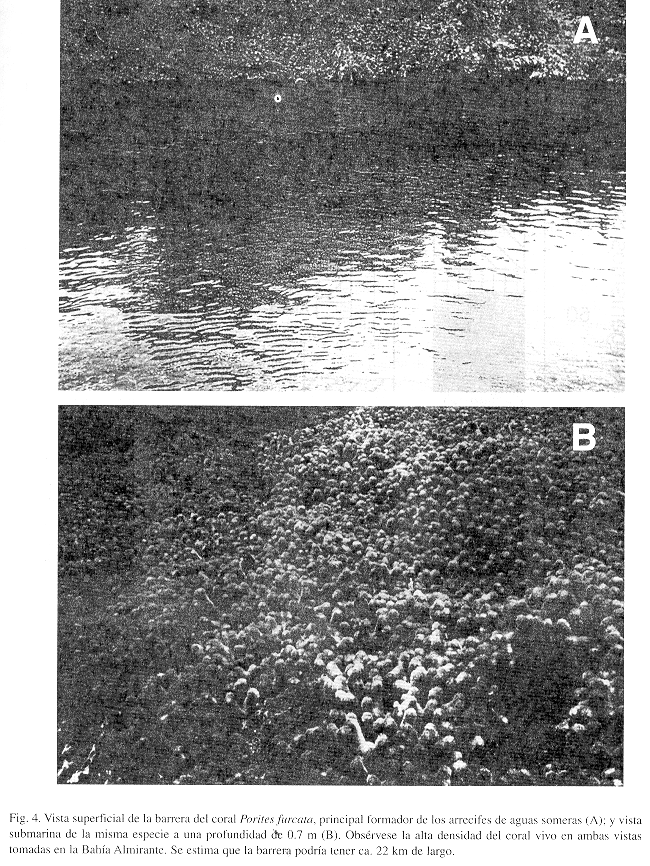
La Figura 3 ilustra la zonación de especies de coralpara todos los arrecifes en conjunto, y basado en la cobertura relativa de las especies. El coral ramificado o ''dedos" Porites furcata es la especie constructora de arrecifes más importante de aguas someras, siendo responsable de ca. 90% de la cobertura. Algo curioso de notar es la alta densidad (ca. 40 individuos/m2) de las poblaciones de la estrella frágil Ophiothrix suensonii encontradas en estadios juveniles y adultos cubriendo los arrecifes de P furcata. Además, es importante destacar que se encontró al coral P furcata formando una barrera arrecifal casi continua, hasta 2 m de profundidad, de ancho variable entre 6 y 25 m y una longitud de ca. 21.5 km, desde la primera punta de manglar que sobresale al sur del Río Oeste hasta la Punta Bella Vista en el extremo sureste de la Ensenada Grande (ver Figs. 1 y 4). La barrera es interrumpida en sectores donde desembocan ríos y quebradas y donde se forman pequeñas ensenadas de mangle, por lo general a distancias inferiores a los 200 m. Otras barreras similares se observaron alrededor de la Isla Pastores (ca. 11 km) y el lado Oeste de Isla Cristóbal, desde Punta Juan hasta Punta Rodríguez (ca. 8 km), pero estas serán descritas en un trabajo consecuente. La segunda especie constructora de arrecifes que domina de 3 a 6 m de profundidad es el coral "lechuga" Agaricia tenuifolia (>40%), donde se mezcla con especies de crecimiento masivo como Calpophyllia natans, Montastraea cavernosa, M. franksi y Siderastrea siderea, entre otras (Fig. 3). Hacia aguas más profundas, de 10 m en adelante, domina el coral S. siderea (ca. 50%) conjuntamente con A. tenuifolia (ca. 15%) y parches extensos de Madracis mirabilis (ca. 7%), terminando la base del arrecife compuesta de colonias en forma de platos de S. siderea y el alga roja calcárea Amphiroa rigida var. antillana y posiblemente las algas incrustantes Hydrolithion boergesenii y Sporolithon episporum. La especie de coral Agaricia undata, comúnmente encontrada en San Blas a profundidades mayores a los 25 m, se encontró creciendo saludable y abundante (ca. 3%) después de los 10 m de profundidad (Fig. 3).
Otros organismos sésiles que contribuyen notablemente al desarrollo arrecifal son las esponjas. Este grupo resultó bastante diverso, con 31 especies de variadas formas (Cuadro 4), y una cobertura de hasta 13% en algunos arrecifes (Cuadro 3). Por lo general, las esponjas se encontraron más abundantes en profundidades intermedias de hasta 5 m (Fig. 2). Se observa un patrón aparente en la distribución geográfica de la diversidad y abundancia de esponjas; los arrecifes del sector este de la Ensenada Grande, los más alejados del continente (12 y 14), tuvieron la mayor diversidad (Cuadro 4) pero no así la cobertura, que se encontró más alta en los arrecifes cercanos a tierra firme (Fig. 2).
Las macroalgas, que por lo general se encuentran muy abundantes en todos los arrecifes del Caribe, parecieran no ser un problema en esta región interna de la Bahía de Almirante. La ausencia de algas en muchos arrecifes contrasta con la alta cobertura encontrada en un arrecife (14), el único donde se observó un promedio superior al 10% (Cuadro 3), o algo mayor dependiendo de la profundidad (Fig. 2). El alga parda Dyctiota ciliolata es responsable de la cobertura observada en aguas someras, y el alga roja Gelidium pussilum en profundidades mayores a 10 m (Fig. 2). Similarmente, el alga roja coralina Arnphiroa rigida var. antillana domina amplias extensiones en los arrecifes, particularmente el de Punta Bella Vista (12), con coberturas superiores al 15% dependiendo de la profundidad (Fig. 2). Otras algas rojas coralinas incrustantes se distribuyen en todos los arrecifes; Hydrolithon boergesenii y Sporolithon episporum en aguas más profundas y Titanoderma bermudense en aguas someras, siempre creciendo sobre coral muerto.
El grupo de los octocorales se encontró poco diverso, con alrededor de ocho especies, formando por lo general parches monoespecíficos dentro del arrecife, principalmente compuestos por Pseudopterogorgia spp., Erythropodium caribaeorum, o Briareum asbestinurn (Cuadro 5). Además, se encontró un extenso parche de octocorales (ca. 3 800 m2) y a una profundidad de 2-3.5 m, formada principalmente por Pseudopterogorgia americana y P. acerosa en altas densidades (ca. 3-7 colonias/m2), y Gorgonia flabellurn hacia las márgenes del parche, ubicado entre Punta Pondsock y el arrecife de este nombre (7). La mayor cobertura de octocorales (2%) se encontró en el arrecife insular del Cayo Ambrosia (3) (Cuadro 3).
Laguna de Chiriquí: Se encontró únicamente un arrecife coralino dentro de la Laguna de Chiriquí (Fig. 1). Este arrecife (15) presentó un desarrollo arrecifal limitado hasta los 3 m de profundidad, una diversidad de corales no mayor a las siete especies, y una cobertura de coral vivo de 24% (Porites furcata), que podría considerarse alta. La mayor parte de este único arrecife esta cubierto por substrato desnudo o arena (73%) y un porcentaje reducido de esponjas (2.4%) (Cuadro 3, Fig. 2). Se encuentran parches arrecifales de pequeño tamaño distribuidos dentro de la laguna al sur de la Isla Loma Partida, y al Oeste de la Península Valiente (alrededores del Cayo Paterson), y a 3-6 m de profundidad. Estos parches aunque conspicuos en cobertura de organismos sésiles, principalmente corales agarícidos y esponjas, no fueron descritos por considerarse comunidades coralinas, y encontrarse en avanzado estado de deterioro posiblemente debido a la sedimentación en la laguna.
Las praderas de hierbas marinas y los bosques de mangle dentro de la Laguna de Chiriquí están bien desarrollados, y en cierto modo su estructura y composición son similares a la descrita para la otra región. La distribución de bosques de manglar es casi continua a lo largo de toda la costa. Las áreas deforestadas de manglar y bosque coinciden por lo general con áreas de ganadería y agricultura cercanas a Punta Róbalo y los alrededores de las quebradas Las Cañas, La Tronosa, Grande y Cauchero, donde se observan amplios asentamientos de campesinos, o áreas alteradas por los poblados indígenas al lado Oeste de la Península Valiente. Se observa deforestación avanzada en la Isla Loma Partida. El bosque de manglar está bastante desarrollado entre las quebradas Los Chiricanos y Molejón, donde el bosque puede alcanzar alturas superiores a los 25 m, y conformado en su mayoría por especies de Avicennia germinans y faguncularia racemosa, entremezclado con el mangle Pelliciera rhizophorae. El sector noroeste del área de estudio está conformado por numerosas islas de mangle rojo Rhizophora mangle bien conservado y de bajo porte. Las praderas de hierbas presentan una composición de especies de organismos asociados similar a Bahía Almirante, aunque pareciera que la densidad de Thalassia y la longitud de sus hojas es mayor, creciendo favorablemente en sedimentos lodosos.
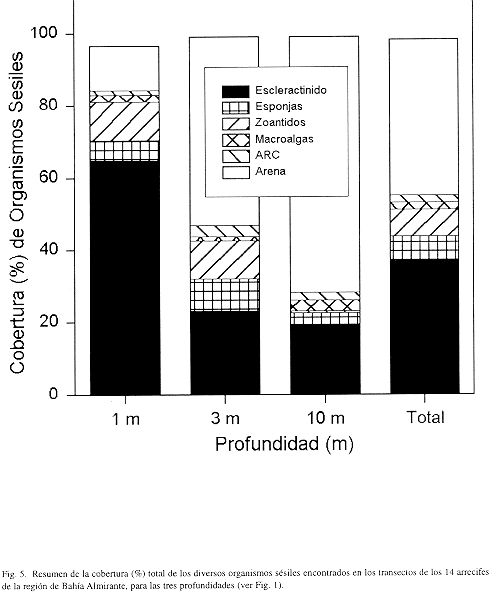
En resumen, si se analiza la cobertura global (todos los 15 arrecifes estudiados) de los principales organismos sésiles, incluyendo los corales, encontramos para los arrecifes continentales una cobertura promedio superior a 35% de corales (incluye 0.8% de Millepora spp.), 2% macroalgas y algas coralinas, 6% esponjas, y 7% zoántidos (Fig. 5). Aproximadamente el 43% del substrato arrecifal está ocupado por arena. Estas coberturas varían considerablemente con la profundidad, encontrándose un 66% de corales en aguas someras y un promedio de 20% en aguas más profundas (Fig. 5), y diferencias significativas entre la cobertura a 1 m y las otras dos profundidades (Anova Kruskal-Wallis H= 21.8, P<0.0001; y SNK). De igual forma se encontró diferencias significativas para las esponjas de mayor profundidad (H= 10.4, P= 0.005), pero con coberturas similares entre 1 y 3 m. Las cobertura de macroalgas y algas rojas coralinas no variaron entre profundidades (H= 1.58, P= 0.452, H= 0.12, P= 0.942; respectivamente). Los zoántidos mostraron diferencias significativas en las profundidades de l m y 3m con respecto a la más profunda (H= 22.1, P< 0.0001).
Los arrecifes continentales estudiados en Bocas del Toro están bien desarrollados y bastante saludables si se les compara con otros arrecifes de Panamá. La cobertura promedio de 40% para coral vivo encontrada en la región de la Ensenada Grande, Bahía Almirante, y la cobertura de algas inferior al 3% sitúa a esta región como la mejor preservada el país. Los arrecifes del sector central del Caribe panameño presentaban para 1991 coberturas de coral de 4% en la zona industrial de Bahía Las Minas y de 9% en el sector Portobelo-Isla Grande (Guzmán et al. 1993), y una cobertura de algas superior al 65% en mbas regiones. Además, Shulman y Robertson (1996) informan de coberturas de 28% y 26% para algas y corales, respectivamente para el sector del Porvenir (San Blas) en 1990. Sin embargo, para 1992 se informa de una cobertura de algas de entre 55% y 82% para la región de San Blas y una cobertura de coal vivo inferior al 10% en la zona de Porvenir y de 20% en los Cayos Limones y Holandeses (Guzmán en prep.).
Los corales encontrados dentro del área que comprende el presente estudio representan un 53% de la diversidad total informada para el Caribe de Panamá (Holst y Guzmán 1993). De igual forma se estima que la diversidad de octocorales en el área de estudio podría ser inferior al 20% de las especies conocidas para el resto del país (Guzmán en prep.). En general, las principales especies formadoras de arrecifes del Caribe están presentes dentro del área de estudio. La ausencia de las especies restantes (48%) se podría relacionar con el grado de desarrollo de los arrecifes, el cual no se extiende a profundidades mayores de 20 m, mientras que en otros lugares de Panamá (i.e., San Blas) estos llegan hasta más de 40 m de profundidad. Esta limitación está relacionada con la profundidad de penetración de la luz, controlada por factores físico-químicos locales.
Entre estos factores el más importante observado para la región de la Bahía Almirante, donde ocurre el mayor desarrollo arrecifal en este estudio, es la presencia casi permanente de una capa superficial de agua dulce rica en taninos, mientras que para la región de la Laguna de Chiriquí el factor más importante es la presencia permanente de sedimentos en suspensión asociados a la alta sedimentación de los ríos. Para ambos casos, la penetración de la luz es limitada. Además, la ausencia de ciertas especies puede ocurrir al utilizar hábitats muy específicos que no se encuentran en ninguna de las dos áreas evaluadas (e.g. ambientes de alta energía y oleaje). Por ejemplo, la especie de coral Acropora palmata no se encontró dentro del área de estudio.
La eutroficación, que por lo general conlleva a un incremento notable en la turbidez, es el resultado de múltiples actividades antropogénicas asociadas a la deforestación y erosión de suelos, que incluye la agricultura, descargas de aguas domésticas sin tratamiento y contaminación industrial. La calidad del agua es importante para el desarrollo arrecifal y solamente un número reducido de especies son tolerantes a cambios de salinidad, temperatura y turbidez (Bak 1978, Rogers 1983, Muthiga y Szmant 1987, Acevedo et al. 1989, Rice y llunter 1992). Por lo general, la distribución y abundancia de los corales escleractínidos es determinada por el efecto neto de múltiples factores ambienfales interrelacionados entre si y que pueden actuar de forma aditiva o sinérgica (Tomascik y Sanders 1987). Particularmente, la profundidad de penetración de la luz limita este desarrollo y la dorninancia y diversidad de especies en el arrecife, siendo la turbidez un factor importante en la fisiología del coral que normalmente es alterado con el aumento en la sedimentación de la zona litoral. Se ha demostrado que la turbibez no solo disminuye la lasa fotosintética en el coral, sino que causa un incremento inmediato en la utilización de moléculas orgánicas que sirven de reserva para mantener procesos metabólicos de alto costo energético como son la producción de mucus y la limpieza de sedimentos (Kendall et al. 1985).
La composición y textura de los sedimentos a lo largo de la costa de la Laguna de Chiriquí, formados por restos de moluscos, grano muy fino y alto contenido de materia orgánica (M. Best com. per.). y una alta concentración de silicatos y nitratos en la columna de agua (L. D'Croz com. per.), indican una alta influencia de sedimentos silicoclásticos provenientes de ríos y quebradas. La costa de la laguna está afectada por descargas continuas de sedimentos provenientes de varios ríos, principalmente los Cricamola, Manantí, Uyama, Guarumo y Cauchero, cuyas plumas de agua dulce y sedimentos se extienden muchos kilómetros desde sus desembocaduras, posiblemente limitando el asentamiento y desarrollo de comunidades coralinas dentro de la Laguna de Chiriquí. No obstante, ocurre lo contrario dentro de la Bahía Almirante, donde a excepción del Río Oeste cerca de la Ciudad de Almirante, las descargas de los ríos son bajas todavía, y la turbidez no está directamente asociada a sedimentación como se explicó anteriormente.
Existen varios mecanismos de limpieza de sedimentos en corales, siendo la producción de mucus una de ellas. El movimiento de limpieza de sedimentos sobre la colonia de coral es más efectivo si existen corrientes de agua fuertes, de lo contrario al no ser removido el mucus con el sedimento atrapado en este podría observarse efectos letales sobre el organismo (Bak y Elg ershuizen 1976). Ambas regiones estudiadas de Bocas del Toro, no están sujetas a grandes corrientes o oleaje que permitan a los corales limpiarse el sedimento y ayudar en la remoción de mucus. Actualmente, no se observa producción generalizada de mucus en las colonias de coral, lo que sugiere que los corales están bastante adaptados a las condiciones ambientales actuales, con algunas excepciones. Pareciera que las especies de corales que dominan estos arrecifes, Porites furcata, Agaricia tenuifolia, Madracis mirabilis, M. decactis y Siderastrea siderea, presentan características peculiares que les permiten vivir muy cercanas a sus límites de tolerancia, siendo algunas de estas cualidades la foto-adaptación, las cuales les permite vivir completamente autótrofos en ambientes difíciles (sensu Edmunds y Spencer-Davies 1989), la eficiencia en la remoción de sedimentos (Bak 1978, Rogers 1983, Tomascik y Sanders 1987) y la alimentación de material particulado en suspensión capturado por medio de redes de mucus (Lewis y Price 1975). Además, la mayoría de las colonias masivas (Siderastrea, Montastraea, Stephanocoenia) se encuentran creciendo en forma horizontal (forma de plato) en aguas después de 5 m de profundidad, lo que indica que se han adaptado a ambientes de poca luminosidad como ocurre en otros hábitats a profundidades de más de 25 m. En Costa Rica se observa este mismo fenómeno de adaptación y desarrollo arrecifal limitado en respuesta a altas tasas de sedimentación (Cortés y Risk1985).
El escenario anteriormente descrito para estos arrecifes de Bocas del Toro -alta cobertura de corales y baja cobertura de algas- es difícil de explicar en estos momentos debido a la falta de información general para la zona. La mayor parte de los arrecifes del Gran Caribe se encuentran parecidos a las áreas ya descritas para el resto de Panamá (baja cobertura de coral vivo y alta de macroalgas), y han sufrido en forma similar las consecuencias asociadas a disturbios naturales y antropogénicos (ver Brown 1987, Goenaga 1991); el calentamiento de las aguas y blanqueamiento de corales (Brown y Ogden 1993, Goreau y Hayes 1994), falta de herbívoros producto de la mortandad masiva del erizo Diadema antillarum y la sobrepesca generalizada (Lessios et al. 1984, Hughes 1994), y enfermedades de corales (Gladfelter 1982), entre otros. Eventualmente, el área de estudio no ha escapado a algunos de estos eventos, y podría explicarse la ausencia de macroalgas y por ende la alta cobertura de coral vivo debido a: (i) la composición de corales tiende a especies más resistentes. La tolerancia de las principales especies constructores de arrecifes a ambientes de alta turbidez las hace a su vez más resistentes a disturbios ambientales; (u) la alta densidad (no estimada) del erizo Echinometra viridis encontrada en todos los hábitats del arrecife e inclusive en las praderas de pastos marinos. Estas altas densidades del erizo posiblemente han mantenido el nivel de herbivoria requerido por estas comunidades bajo condiciones saludables. Se conoce que en San Blas y Galeta aumentaron considerablemente las poblaciones de E. viridis al ocurrir la mortalidad masiva de D antillarum en 1983 (Lessios et al. 1984).
Sin embargo, en los arrecifes de estas dos últimas regiones no se observa el mismo efecto de herbivoría que posiblemente ocurre en Bocas del Toro, y la alta cobertura de algas persiste. Además, otros herbívoros importantes (peces) han sido eliminados de la región de la Bahía Almirante, donde se observan claros efectos de sobrepesca en toda el área. Se observa un esfuerzo de pesca alto por parte de los indígenas, quienes emplean continuamente redes sobre los arrecifes, resultando ya en una escasez de peces a todos los niveles tróficos. Los efectos negativos de la sobrepesca en la estructura del arrecifes coralinos son ampliamente entendidos (ver Hughes 1994, Roberts 1995); y (iii) una combinación de ambos procesos, corales resistentes y alta herbivoría.
Finalmente, la información de este estudio permite valorar adecuadamente el estado actual de los arrecifes en este sector de Bocas del Toro. Los arrecifes del área en consideración, aunque no presenta un atractivo turístico explotable directamente, bien cumplen las funciones principales a nivel de ecosistema y sirven de apoyo a poblaciones indígenas locales (Jaén 1993). El proyectado desarrollo de esta región traerá como consecuencia inmediata la sobrepoblación en la zona costera, que se réflejará en la destrucción de manglares, mayor demanda por recursos del mar ya sobreexplotados a nivel artesanal, cambios culturales en las poblaciones indígenas quienes tendrán que competir más por espacio y recursos naturales, y una disminución de la calidad del agua al aumentar la eutroficación y sedimentación. Actualmente la situación es difícil para las comunidades indígenas locales, quienes ya han comenzado a explotar los recursos del bosque y mar en forma comercial, y no a nivel de subsistencia (Jaén 1993). Es notable, que lo anteriormente descrito afectará la productividad y estructura de las comunidades arrecifales, manifestándose en una mortalidad mayor de corales y una bajas tasas de reproducción y asentamiento de nuevos corales, efectos que ya han sido documentados ampliamente para otras regiones (Loya 1976, Bak 1978, Cortés y Risk 1985, Tomascick y Sander 1987, Baines y Morrison 1990, Rogers 1990, Goenaga 1991, Tomascik 1991, Wittenberg y Hunte 1992, Guzmán et al. 1993, Guzmán y Holst 1993). Es probable que los arrecifes del área estudiada ya estén viviendo cercanos al límite de tolerancia a cambios ambientales; su composición y zonación actual son características que así lo indican.
Conclusiones y recomendaciones
El presente estudio demuestra claramente la presencia de arrecifes bien desarrollados en la zona costera continental (no insular), zona esta que podría ser la más propensa a la influencia terrígena asociada a la modificación de bosques y cuencas hidrográficas. Los arrecifes coralinos aquí descritos definitivamente pueden ser considerados como los de mayor cobertura de coral vivo descritos para el Caribe panameño, si se les compara con otros arrecifes del resto del país (Guzmán et al. 1993, Guzmán in prep.). Por lo tanto considerarnos importante se protejan estas comunidades litorales de la sedimentación y deforestación, y podemos señalar la imperante necesidad de encontrar soluciones adecuadas que ayuden a mitigar en lo posible el inminente impacto negativo que ocasionará el desarrollo no planificado de la Provincia de Bocas del Toro sobre las comunidades marítimo-costeras. Las recomendaciones que se presentan a continuación, aunque algunas de aparente carácter científico, podrían proporcionar un punto de partida para entidades gubernamentales encargadas del manejo de los recursos naturales y del desarrollo de la provincia.
(1) Estudiar la dinámica y patrones de corrientes marinas dentro de las dos regiones, lo que permitiría comprender con precisión la distribución de los sedimentos acarreados por los ríos y quebradas a lo largo de la costa. A su vez esto permitiría predecir cuales arrecifes y comunidades costeras estarían más susceptibles a las altas tasas de sedimentación a períodos de mediano y largo plazo.
(2)Establecer a la brevedad leyes y regulaciones que controlen y eviten la deforestación de los bosques de manglar y los bosques lluviosos en montañas adyacentes a la costa. Hasta el momento la mayor parte del area presenta una cobertura boscosa saludable y las bocas de los ríos y quebradas no están muy alteradas. Sin embargo, es de esperarse que los planes de desarrollo para la Provincia de Bocas del Toro promueva la invasión de tierras, la deforestación, la ganadería, y la sobrepoblación a lo largo de la costa, con consecuencias ambientales bastantes conocidas (Myers 1997).
(3)Sugerimos se diseñe e implemente una campaña de información ambiental que permita a los indígenas y latinos locales conocer más sobre la presencia e importancia de los arrecifes, lo susceptible de estas comunidades naturales a cambios físicos en la costa, y la necesidad de que los mismos pobladores los protejan. Actualmente, se observan señales de sobrepesca; la abundancia y diversidad de peces asociados al arrecife son muy bajas en ambas regiones estudiadas. De igual forma, ya se observan señales de deforestación y ganadería cercanas al litoral.
Agradecimientos
Agradecemos a Juan Maté por su ayuda durante el trabajo inicial en el campo. A 1. Valdespino y O. Vallarinos de la Asociación para la Conservación de la Naturaleza (ANCON), y 1. Rubinoff, H. Barnes, L. Motta y I. Ivancic del Smithonian Tropical Research Institute (STRI) por apoyo brindado durante esta investigación. Agradecemos a M.E. Tupper y a la tripulación del Y/V Starfire, P. Miller y K. Simpson, por su apoyo logístico incondicional. Agradecemos a X. Guerra por el diseño del mapa y a K. Kauffman por preparar las estadísticas descriptivas. Este estudio fue financiado parcialmente por M.E. Tupper, ANCON y STRI.
Resumen
Se presenta la distribución de azrecifes coralinos a lo lar-go de la costa continental (tierra firrne) de la Bahía Almirante (ca. 83km) y de la Laguna de Chiriquí (ca 125km), Provincia de Bocas del Toro, Panarná. Se describe la estructura de 15 arrecifes coralinos, los cuales representan la diversidad de condiciones hidrológicas y orográficas de la zona costera con mayor inítuencia terrígena de Bocas del Toro. Los arrecifes encontrados en la Bahía Almirante se encuentran bien desarrollados hasta una profundidad máxima de 23 m y una diversidad de corales madrepóricos de 33 especies, lo cual representa aproximadamente un 53 % de la diversidad conocida para Panamá. las coberturas de coral vivo observadas para la región de estudio oscilan entre 20 % y 50 % depediendo de la profundidady se estima un promedio total de 35 % casi el doble de la cobertura informada para otras regiones de Panamá (San Blas) donde se presume que los arrecifes estan mejor protegidos.
La zonación y composición de especies de corales son similares en toda el área de estudio; Porites furcata domina los arrecifes someros hasta 2 m de profundidad, con una cobertura de ca. 90% seguido por una zona dominada por Agaricia tenuifolia, (40%) hasta los 6 m. seguido de una zona principalmente compuesta de A. tenuifolia (15%) Madracis mirabilis (7%) y Siderastrea siderea (50%) hasta los 15 m de profundidad, terminando en una zona profundad dominada por algas rojas calcareas Amphiroa rigida, Hydrolithon boergesenii, Sporolithon episporum y el coral S. siderea. El coral Agariciaundata, común en habitats profundos se encontró relativamente abundante (3%) y creciendo después de los 10 m. Se informa por primera vez para la región la existencia de una barrera coralina construida por Porites furcata, semi-continua de ca. 22 km de largo y que bordea la zona costera de la Bahía Almirante. El grupo de las esponjas, segundo en importancia, está ampliamente distribuido en todos los arrecifes con una diversidad de 31 especcies y cobertura promedio de 6%. Curiosamente, se encontró un solo arrecife de los estudiados con cobertura de macroalgas del 10 %, mientras que los rstantes arrecifes presentan cobertura promedio de macroalgas inferiores al 4%. Solamente se encontró un arrecife coralino dentro de la Laguna de Chiriquí, con desarrollo limitado hasta 3 m de profundidad, una diversidad de corales no mayor a 7 especies y cobertura de coral de 24% (Porites furcata). La costa de la Laguna Chiriquí está afectada por descargas continuas de sedimentos provenientes de varios ríos, principalmente Cricamola, Manantí, Uyama, Guarumo y Cauchero, cuyas plumas de agua dulce y sedimentos se extienden mucchos kilómetros desde sus desembocaduras, limitado el asentamiento y desarrollo de comunidades coralinas. La distribución vertical y dominancia de ciertas especies de corales en los arrecifes aquí descritos (e.g. Siderastrea siderea, Porites furcata, Agaricia tenuifolia, Madracis decactis, y M. mirabilis) indican que estas comunidades se formaron adaptándose gradualmente a condiciones variables de calidad de agua (salinidad, temperatura y penetración de la luz) relativamente no óptimas para un desarrollo arrecifal más diversos y de aguas más profundas. Esto sugiere que estas especies pesar de ser bastante resistentes a cambios ambientales, viven actualmernte muy cercanas a sus límites de tolerancia ecofisiológica, lo cual las hace particularmente vulnerables a cambios puntuados. Cuaquier proyecto de desarrollo no planificado incrementará la sedimentación hacia la zona costera debilitando a los organismos arrecifales en su totalidad, afectándose así la funcionalidad del ecosistema.
Referencias
Acevedo, R., J. Morelock & R.A. Olivien. 1989. Modification of coral reef zonation by terrigenous sediment stress. Palaios 4: 92-100.
Baines, G. B.K. & R.J. Morrison. 1990. Marine pollution from landuse in the Pacilic Islands region. Mar. Pollut. BuIl. 21:506-515.
Ballou, T.G., M.S. Brown, R.E. Dodge, C.D. Getter, A.H. Knap, & T.D. Sleeter. 1985. Tropical oil pollution investigations in coastal Systems (TROPICS): a technical status report. Research Planning Institute. Columbia. 128 p. [ Links ]
Ballou, T.G., S.C.Hess, R.E.Dodge, A. H Knap & T.D. Sleeter.1989a. Effects of untreated and chemically dispersed oil on tropical marine communities: A long-term field experiment. Proceedings of the 1989 Oil Spill Conference, American Petroleum Institute, Washington, D.C. 447-454 [ Links ]p.
Ballou, T.G., R.E. Dodge, S.C. Ress & A.H. Knap. 1989b. Tropical oil pollution investigations in coastal systems (Tropics): The effects of untreated and chemically dispersed Prudhoe Bay crude oil on mangroves, seagrasses, and corals in Panama, p. 229-256. In L.M. FIaherty (ed.). Oil dispersants: new ecological approaches. American Societyfor Testing and Materlais, Philadelphia.
Bak, R.P.M. & J.H.B.W. Flgershuizen, 1976. Patterns of oil-sediment rejection in corals. Mar. Biol. 37:105-113.
Bak, R.P.M. 1978. Lethal and sublethal effects of dredging on reef corals. Mar. Pollut. Bull. 9:14-16. [ Links ]
Bayer, F.M. 1961. The shallow-water octocorallia of the West Indian region. Martinus Nijhoff, The Hague. 373 p.
Brown, B.E. 1987. Worldwide death of corals-natural cyclical events or man-made pollution? Mar. Pollut Bulí. 18: 9-13.
Brown, E.E. & J.C. Ogden.1993. Coral bleaching. Scientific America 268: 64-70. [ Links ]
Cairns, S.D. 1982. Stony corals (Cnidaria: Hydrozoa, Scleractinia) of Carrie Bow Cay, Belize. Smithsonian Contrib. Mar. Sci. 12: 271-302. [ Links ]
Coates, A.G. & J.B.C. Jackson. 1998. Neogene sedimentary basins of Costa Rica arid Panama. 1 lth Conference ínter. Bryozoology Association, Panama. 39 p.
Collins, LS., A.F Budd & A.G. Coates. 1996. Earliest evolution associated with closure of the Tropical American Seaway. Nat. Acad. Sci. Proc. 93: 6069-6072. [ Links ]
Cortés, J. & H.M. Guzman. 1985. Organismos de los arrecifes coralinos de Costa Rica III. Descripción y distribución geográfica de los corales pétreos (Anthozoa: Scleractinia) en la costa Caribe. Brenesia 24: 63-123. [ Links ]
Cortés, J. & M.J. Risk. 1985. A reef under siltalion stress: Cahuita, Costa Rica. Buil. Mar. Sci. 36: 339-356.
DMA (Delfense Mapping Agency). 1988. Atlas of Pilot Charts for the North Atlantic Ocean. DMP, 37 p.
Dodge. R.E. & A.H. Knap. 1993. Long-term monitoring (2.5 years) of effects of short-term fleid exposures of stony corals to dispersed and undispersed crude oil, p. 87-93.In R.N. Ginsburg (ed.). Global aspects of coral reefs: health, hazards and history. University of Miami, Florida.
Edmunds, P.J. & P. Spencer-Davis. 1989. An energy budget for Porites porites (Scleractinia), growing in a stressed environrnent. Cora] Reefs 8: 37-44.
Gladfelter, W.B. 1982. White-band disease in Acropora palmata: implications for the structure and growth of shallow reefs. Bull. Mar. Sci. 32: 639-643. [ Links ]
Greb, L., B. Saric, H. Seyfried, T. Broszonn. S. Brauch, G. Gugau, C. Wiltschko & R. Leinfelder. 1996. Ókologie ind sedimentologie eines rezenten rampensystem an der Karibikkuste von Panama. Profil, Band 10, Universitat Stuttgart, 168p. [ Links ]
Glynn, PW. 1972. Observations on the ecology of the Caribbean and Pacific coasts of Panarna. Bull. Biol. Soc. Wash. 2:13-30.
Goenaga, C. 1991. The state of coral reefs in the wider Caribbean. Interciencia 16:12-20. [ Links ]
Gordon, B.L. 1982. A Panarna forest and shore: natural histoty and Amerindian culture in Bocas del Toro. The Boxproof Press, California. 178 p.
Goreau, T.J. & R.L. Hayes. 1994. Coral bleaching and ocean 'hot spots". Ambio 23:176-180. [ Links ]
Gundlach, E..R., G. Moss, F. de Vincenti & J. Janssen. 1985. Resource mapping and contingency planning, PTP Pipeline Facilities, Panama. Proceedings 1985 Oil Spill Conference, Los Angeles, California. 229-234. [ Links ]
Guzmán, H.M. & J. Cortés. 1985. Organismos de los arrecifes coralinos de Costa Rica. IV. Descripción y distribución geográfica de octocorales (Anthozoa: Octocorallia) en la costa Caribe. Brenesia 24: 125-173. [ Links ]
Guzmán, H. M. & C.E. Jiménez. 1992. Contamination of reef corals by heavy rnetals along the Caribbean coast of Central America (Costa Rica and Panama). Mar. Pollut. Bull. 24: 554-561. [ Links ]
Guzmán, H.M. & 1. Holst. 1993. Effects of chronic oil-sediment pollution on the reproduction of the Caribbean rref coral SideTasirea siderea. Mar. Pollut. Bulí. 26:276-282.
Guzmán, H.M., I.B.C. Jackson & 1. Holst. 1993. Changes aad recovery of subtidal reef corals, p. 361-446. In B.D. Keller and J.B.C. Jackson (eds.). Long-term assessment of the oil spill at Bahía Las Minas, Panamá, synthesis Report. u.S. Department of the Interior, New Orleans.
Hatcher; B.G. 1988. Coral reef primary productivity: a beggar's banquet. TREE 3:106-111. [ Links ]
Hatcher; B.G. 1990. Coral reefs primary productivity: a hierarchy of pattern and process. TRIPE 5:149-155.
Holst, I & H.M. Guzmán. 1993. Lista de corales hermatípicos (Anthozoa: Scleractinia; Hydrozoa: Milleporina) a ambos lados del istmo de Panamá. Rev. Biol. Trop. 41: 535-540. [ Links ]
Hughes, T.P. 1994, Catastrophes, phase shifts, and large-scale degradation of a Caribbean coral reef. Science 265:1547-1551 [ Links ]
Humann, P. 1994a. Reef Creatures Identification: Florida, Caribbean and Bahamas. Paramount Miller Graphics, Florida. 320 p. [ Links ]
Humann, P. 1994b. Reef Coral Identification: Florida, Caribbean and Bahamas. Paramount Miller Gruphics, Florida. 240 p.
IGNTG (Instituto Geográfico Nacional "Tommy Guardia"). 1988.Atlas Nacional de la República de Panama. IGNTG, Panamá. 222 p. [ Links ]
INRENARE (Instituto Nacional de Recursos Naturales Renovables). 1995. Informe de cobertura boscosa a 1992. Dirección de Cuencas Hidrográficas. Sistema de Información Geográfica. INRENARE, Panamá.27 p, [ Links ]
Jackson, J.B.C. & A.G. Coates. 1993. Biología y geología del Istmo de Panamá. p. 35-41. In S. Heckadon-Moreno (ed.). Agenda Ecológica y Social para Bocas del Toro. Impresora Continental, S.A., Panamá. [ Links ]
Jaén, B.C. 1993. Los pueblos indigenas y el uso de los recursos naturales en Bocas del Toro, p. 119-135.In S. Heckadon-Moreno (ed.). Agenda Ecológica y Social para Bocas del Toro. Impresora Continental, S.A., Panamá. [ Links ]
Kendall, Jr., J.J., E.N. Powell, S.J. Connor, TJ. Btight & CE. Zastrow.1985. Effects of turbidity on calcification rate, protein concentration and the free amino acid pool of the coral Acropora cervicornis. Mar. Biol. 87: 33-46.
Lessios, H.A., J.D. Cubit, D.R. Roberison, M.J. Shulman, M.R. Parker, S.D. Garrity & S.C. Levings. 1984. Mass mortality of Diadema antillarurn on the Caribbean coast of Panama. Coral Reefs 3: 173-182.
Lewis, J.B. & W.S. Price. 1975. Feeding mechanisms and feeding strategies of Atlantic reef corals. J. ZooI., 176: 527-544.
Littler, D.S., M.M. Littler, K.E. Bucher & J.M. Norris. 1989. Marine plants of the Caribbean: a field guide from Florida to Brazil. Smithsonian Institution Press, Washington, D.C. 263 p. [ Links ]
Loya Y. 1972. Comnunity structure and species diversity of hermatypic corals at Eilat, Red Sea. Mar. Biol. 13:100-123. [ Links ]
Loya, Y. 1976. Effects of water turbidíty and sedimentation on the community structure of Puerto Rican corals. Bull. Mar. Sci. 26: 450-466. [ Links ]
Mou-Sue, L. 1993. Parque Nacional Marino Isla Bastimentos, p. 163-173. In S. Heckadon-Moreno (ed.).Agenda Ecológica y Social para Bocas del Toro. Impresora Continental, S. A., Panamá.
Muthiga, A.N. & A.M. Szmant. 1987. The effects of salinity stress on the rates of aerobic respiration and phosysnthesis in the hermatypic coral Siderastrea siderea. Biol. Bull. 173: 539-551 [ Links ]
Muzik, K. 1982. Octocorallia (Cnidaria) from Carrie Bow Cay, Belize. Smithsonian Contrib. Mar. Sci. 12:303-310. [ Links ]
Myers, J. 1997. Mitigating the effects of land-use change on tropical aquatie systems. Biodiversity Network News, lO: 3-4.
Rice, S. A. & C. L. Hunter. 1992. Effects of suspended sediment and burial on scleractinian corals from West Central Florida patch reefs. Bull. Mar. Sci. 51: 429-442. [ Links ]
Roberts, C.M. 1993. Coral reefs: health, hazards and history. TREE 8: 425-427. [ Links ]
Roberts C.M. 1995. Effects of fishing on the ecosystem structure of coral reefs. Conserv. Biol. 9:815-826. [ Links ]
Rodriguez, E., R. Almanza & R.Alvarado. 1993. Situación biofísica y arnbiental de la Provincia de Bocas del Toro, p.55-72. In S. Heckadon-Moreno (ed.). Agenda Ecológica y Social para Bocas del Toro. Impresora Continental, S.A. Panamá. [ Links ]
Rogers, C.S. l983.Sublethal and lethal effects of sediments applied to common Caribbean reef corals in the field. Mar. Pollut. Bull. 14: 378-382.
Rogers, C.S.l990. Responses of coral reefs and reef organisms lo sedimentation. Mar. Ecol. Prog.Ser. 62: 185-202. [ Links ]
Sefton, N. & S.K. Webster. 1986. A field guide to Caribbean Reef lnvertebrates. Sea Challengers, Monterey, California. 112 p
Shulman, M.J. & D.R. Robertson. 1996. Changes in the coral reefs of San Blas, Caribbean Panama: 1983-1990. Coral Reefs 15: 231-236. [ Links ]
Spurgeon, J.P.G. 1992. The economic valuation of coral reefs. Mar. Pollut. Bull. 11:529-536. [ Links ]
Sullivan, K.M. & M. Chiappone. 1992. A comparison of belt quadral and species presence/absence sampling of stony corals (scleractinia and Milleporina) and sponges for evaluating species patteming on patch reefs of the central Bahamas. Bull. Mar. Sci. 50: 464-488. [ Links ]
Sullivan, K.M., M. Chiappone & C. Lott. 1994. Abundance patterns of stony corals on platforrn margin reefs of the Caicos Bank. Bahamas J Sci. 1: 2-12 [ Links ]
Summers, K., E. Berrningham, L. Weigt, S. McCafferty & L. Dahlstrom. 1997. Phenotypic and genetic divergence in three species of dart-poison frogs with contrasting parental behavior J. Heredity 88: 8-13. [ Links ]
Tomascik, T. & F. Sanders. 1987. Effects of eutrophication un reef-building corals II. Structure of scleractinians coral communities on fringing reefs, Barbados, West Indies. Mar. Biol. 94: 53-75.
Tomascik, T. 1991. Settlement patterns of Caribbean scleractinian coralson artificial substrata along a eutrophication gradient, Barbados, West Indies. Mar. Ecol. Prog. Ser., 77: 261-269.
Wittenberg, M. & W. Hunter. 1992. Effects of eutrophication and sedimentation on juvenile corals. Mar.. Biol. 112:131-138.
Zea, S.1987. Esponjas del Caribe Colombiano. Editorial Catálogo Científico, Bogotá. 286 p [ Links ]
N° Latitudid/Longitud Localidad Metros
Especies 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
Acropora cervicornis x
Agaricia agarícites x x x x x x x x x x x
Agaricia lamarcki x x x x x x x x x x x x x x
Agaricia tenuifolia x x x x x x x x x x x x x x x
Agaricia undata x
Colpophyllia breviserialis x x x x x
Diploria strigosa x x x
Eusmilia fastigiata x x x
Favia fragum x x x x x x x x x x x x x x x
Leptoseris cucullata x x x x x x x x x
Madracis decacris x x x x x x x x x x x x
Madracismirabilis
Millepora alcinornis x x x x x x x x
Montastraea faveolata x x x
Mycetophyllia a!iciae x x
Mycetophyllia lamarckiana x
Oculina diffusa x x
Porites astreoídes x x x x x x x x x x x x x x x
Porites divaricata x
Porites furcata x x x x x x x x x x x x x x x
Scolymia cubensis x x x x x x x x x x x
Siderastrea radians x x x x x x x x x x x x x x
Especies 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
Anthosigmella varians x x x x
Aplysina lacunosa x x
Chondrilla nucula x x x x x
Cliona delitrix x x x x x x x x x
Cribrochalina vasculum x
Ectyoplasia ferox x x x x
Ircinia campana x x
Ircinia felix x
Ircinia strobilina x x x x x
Iotrochota birotulata x x x x x x x x x
Mycale leavis x x x
Verongula rigida x x
Especies 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
Briareum asbestinum x
Erythropodium caribaeorum x x x x x x x x
Eunicea calyculata x
Eunicea sp. x
Pseudoplexaura sp x x x x
Condylactis gjgantea* x x x x x x x
Sichodacyla helianthus x x
Zoanthus pulchellus** x x x x x x x x x x x
Totales 6 1 4 5 3 2 2 5 4 3 5 6 3 9 1
1Smithisonian Tropical Research Institute. Unit 0948, APO AA 34002-0948, U.S.A.

