










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.46 n.2 San José Jun. 1998
Abstract
A reproductive experiment shows that Passiflora foetida is autocompatible. Observations of floral characteristics and visitors during anthesis, plus the analysis of pollen allowed identification of floral syndrome (melittophily) and functions for each visitor. Anthesis occurs from 6 to 11 AM. Three floral phases were identified: 1) stigmas above anthers, 2) stigmas at anther level, 3) stigmas above anthers; radii, petals and sepals become incurved. The stigmas are receptive during the whole anthesis. The concentration of sugars in nectar is 34 %. The predominant color in the visible spectrum is white. The stamens and the gynoecium contrast against the limen and androgynophore in the UV spectrum, maybe a nectar guide. Three species of hymenopterans are the most frequent and constant visitors: Ptiloglossa tarsata (Colletidae) always contact the anthers and the stigmas while sucking nectar, carry a high percentage of P. foetida pollen and visit flowers in phases 1 and 2; they can be considered the main pollinators. Pseudaugochloropsis sp. (Halictidae) rarely contact anthers and stigmas while puncturing the limen to access nectar, and visit flowers in phases 2 and 3; they are nectar robbers that rarely pollinate. Augochlorella sp. (Halictidae) collect pollen without touching the stigmas and visit flowers in phases 2 and 3; they are pollen thieves.
Key words
Reproductive system, floral syndrome, flower visitors, pollinators, Passiflora.
Passiflora foetida L. posee una amplia distribución en América Central y Sudamérica (Killip 1938). En la Argentina, si bien ha sido recolectada en 15 provincias del centro-norte en los últimos dos siglos, según la colección del herbario SI (Holmgren et al. 1990), parece haber desaparecido de la provincia de Buenos Aires, donde se encontró, aunque escasa, en 1726 (Burkart 1963); esto se debió, probablemente, a la alteración del ambiente natural, convertido en campos agrícolas. A pesar de poseer propiedades medicinales (Killip 1938), comestibles (Martin & Nakasone 1970) y ornamentales, es considerada maleza en varios lugares (Engstrand 1980). Janzen (1968) y Gottsberger et al. (1988) observaron los visitantes florales y el comportamiento reproductivo de esta especie en América Central y Brasil, respectivamente, pero se desconocen los visitantes en el extremo austral de su distribución. Tampoco se conoce su grado de autocompatibilidad (Smiley 1991), aunque Gottsberger et al. (1988) aseguran que es autoincompatible, pero no presentan los resultados de los experimentos que realizaron. Esta especie aún no se cultiva; sería importante hacerlo, tanto para su aprovechamiento como para la preservación del recurso. Para diseñar los cultivos es necesario esclarecer su biología reproductiva. El presente trabajo tiene como objetivo contribuir a ello.
Materiales y Métodos
Area de estudio: la floración ocurre entre noviembre y abril. Las observaciones se llevaron a cabo en Charata (Chaco, Argentina, 27013'S, 61012'O) del 3 al 8 de enero de 1995 y del 6 al 30 de enero de 1996, en una parcela de 4 ha. Las plantas de P. foetida, rastreras-semitrepadoras, muchas veces se hallaban semiocultas por gramíneas de mayor porte; crecían en los bordes de los campos sembrados, a una distancia de 102.2 m ![]() 142.5 (desde 4 a 500 m) (n=11) entre sí. Algunas de las plantas observadas durante 1996 eran las mismas del año anterior. Un espécimen de referencia (Amela García, 138) se depositó en el herbario BAFC (Holmgren et al. 1990).
142.5 (desde 4 a 500 m) (n=11) entre sí. Algunas de las plantas observadas durante 1996 eran las mismas del año anterior. Un espécimen de referencia (Amela García, 138) se depositó en el herbario BAFC (Holmgren et al. 1990).
Sistema reproductivo: se calculó el índice reproductivo P/O (polen/óvulos) según Cruden (1977). Se embolsaron pimpollos próximos a la antesis con bolsas de tul, cuando abrieron, en algunos se indujo la autogamia y en otros la xenogamia. Otros no recibieron tratamiento, a fin de comprobar autogamia espontánea. Las flores que recibieron estos 3 tratamientos se embolsaron hasta la maduración de los frutos, para evitar daños por frugívoros. Por último, se marcaron flores expuestas a las visitas de los polinizadores, que se embolsaron al finalizar la antesis.
Reservas y viabilidad polínicas: se analizó la presencia de almidón, lípidos y proteínas en el polen siguiendo a Johansen (1940). La viabilidad polínica se estimó por la técnica de Greissl (1989) y la de persistencia de reservas (Amela García & Hoc en prensa), según la cual, una vez detectadas, se cuentan los granos teñidos.
Receptividad estigmática: se comprobó siguiendo el método de Osborn et al. (1988), que consiste en colocar peróxido de hidrógeno sobre los estigmas y observar si se producen burbujas.
Olor: se separaron los distintos ciclos florales en sendos frasquitos de vidrio y luego de una hora se realizó una prueba olfatoria.
Patrón de coloración: se compararon los colores de las distintas piezas florales con la carta de Kelly (1965) para su descripción en el espectro visible. Las flores fueron fotografiadas con un filtro a través del cual sólo pasa la luz UV (300-400 nm) (Jones & Little 1983), para obtener una aproximación al patrón de reflexión que presentan en esa parte del espectro, en el cual también ven los insectos.
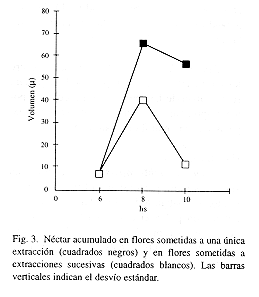
Néctar: la concentración se midió con un refractómetro de mano. El pH, con papel reactivo. En 1995, en flores no embolsadas se midió el néctar ofrecido a distintas horas. En 1996, durante 3 días, se embolsaron flores con bolsas de tul antes de que abrieran. Cada 2 hs se extrajo el néctar acumulado en distintos grupos de flores (n=17) para componer el patrón de secreción (cada flor era muestreada una vez). A otras flores (n=18) se les removió el néctar cada 2 hs para conocer el efecto que provocan las extracciones sucesivas. Las remociones se realizaron con capilares de diámetro conocido, en los que se midió la altura alcanzada por el néctar para calcular el volumen según la fórmula del cilindro.
Visitantes: se observó su actividad en las flores constantemente durante toda la antesis. Un ejemplar de cada especie fue depositado en el Museo Argentino de Ciencias Naturales "B. Rivadavia". Se emplean los vocablos "robo" y "hurto" como traducción de "robbing" y "thieving", respectivamente, usados por Inouye (l980), ya que, según la Real Academia Española (Anónimo 1989), el primero significa "tomar lo ajeno con violencia", lo que corresponde a "robbing" (búsqueda de néctar o polen causando daño a las flores). "Hurto", en cambio, no implica intimidación (Anónimo 1989), corresponde por tanto a "thieving" (obtención de recompensa sin causar daño a las flores). Los sustantivos respectivos que se usan son "ladrón" y "hurtador". Para la denominación de la actividad de los visitantes sigue a Amela García & Hoc (en prensa) y para la denominación de las piezas de la flor, a Puri (1948).
Factores meteorológicos: se registró la temperatura ambiente y la humedad relativa ambiente mediante un termohigrómetro digital.
Resultados
Sistema reproductivo: había 34 ![]() 1 óvulos/ovario (n=5). El índice P/O fue 430
1 óvulos/ovario (n=5). El índice P/O fue 430 ![]() 29 (n=2). Se produjeron frutos con semilla en todos los tratamientos (CUADRO 1). El número de semillas por placenta fue semejante en cada tratamiento y entre tratamientos. En algunos frutos, se observaron unas pocas semillas inmaduras: eran más pequeñas, con menos endosperma (más opaco) y el embrión y la pared seminal no estaban desarrollados completamente. En general, en la base de las placentas se encontraba la mayoría de los óvulos que no fueron fecundados.
29 (n=2). Se produjeron frutos con semilla en todos los tratamientos (CUADRO 1). El número de semillas por placenta fue semejante en cada tratamiento y entre tratamientos. En algunos frutos, se observaron unas pocas semillas inmaduras: eran más pequeñas, con menos endosperma (más opaco) y el embrión y la pared seminal no estaban desarrollados completamente. En general, en la base de las placentas se encontraba la mayoría de los óvulos que no fueron fecundados.
Reservas y viabilidad polínicas: las reservas del polen son lípidos y proteínas. La viabilidad de los granos por la técnica de Greissl (1989) fue del 90 %, y por la de persistencia de reservas, del 98.5 %.
Antesis: todas las flores abrían después que comenzaba una franja de claridad en el horizonte (5.15-5.30 hs), entre las 5.45 y 6.10 hs (n= 42). Raramente, el día anterior a la antesis, ca. 20 hs, algunos pimpollos presentaban los pétalos separados en el ápice, dejando ver los estigmas, pero las anteras aún estaban indehiscentes. Cada flor demoraba en abrir aproximadamente 1 min. Diariamente abría una flor por nudo y por rama, en ocasiones había dos en un mismo nudo. Cada planta producía 2 ![]() 1 flores por día (n=25) (1-6), dependiendo de la cantidad de ramas floríferas.
1 flores por día (n=25) (1-6), dependiendo de la cantidad de ramas floríferas.
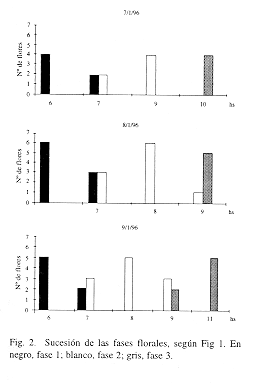
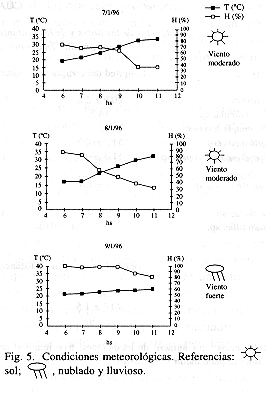
Fases florales: se distinguieron 3 fases (Fig. 1): 1) estigmas por encima de las anteras; éstas, la corona, la corola y el cáliz, perpendiculares al eje floral; 2) estigmas a la altura de las anteras o por debajo de éstas; 3) el ápice de los estilos se recurva, quedando los estigmas por encima de las anteras; los radii, los pétalos y los sépalos se incurvan. La sucesión de las fases florales se muestra en la Fig. 2. Duraban, aproximadamente: la fase 1, hasta las 7 hs; la fase 2, hasta las 9 hs; la fase 3, hasta las 11 hs, al final de la cual la flor se cerraba. Cuando las flores se encontraban a la sombra tardaban más en cambiar de fase. El 9/1/96 llovía al amanecer, las anteras y los estigmas estuvieron más bajos que otros días. Si había ocurrido fecundación, al día siguiente de la antesis ya se detectaba al ovario con mayor tamaño.

Receptividad: los estigmas estaban receptivos en todas las fases florales; el burbujeo con peróxido de hidrógeno era más intenso cuando se hallaban por debajo de las anteras en la fase 2 (n=14).
Olor: el operculum, los pali y radii presentaban un olor tenue, dulce, en las fases 1 y 2. Durante todo el día se percibía olor en las brácteas, igual al de las hojas pero más leve.
Patrón de coloración: en el espectro visible, los colores de las distintas piezas florales no variaban durante la antesis: pistilo, filamentos estaminales y sépalos, distintas tonalidades de verde amarillento; conectivo y borde de las anteras, amarillo claro; tecas, amarillo vívido; operculum, limen, androginóforo y borde de la cara abaxial de los sépalos y los pétalos, blanco; ápice y base de los radii, los pali y máculas del limen, del androginóforo y de los filamentos estaminales, distintas tonalidades de púrpura rojizo. Una misma planta producía, además, flores con los pali y el extremo de los radii púrpura claro o blanco. En la Fig. 1 (A-F) se aprecia que, en el espectro UV, el gineceo y los estambres reflejaban esa luz, contrastando con el resto de las piezas, que la absorbían, principalmente el limen y androginóforo; y también contrastaban con las partes vegetativas de la planta (Fig. 1, E-F).
Néctar: el porcentaje de azúcares totales fue 34 % ![]() 5 (n=9). El pH, 5. La oferta de néctar fue 22.03
5 (n=9). El pH, 5. La oferta de néctar fue 22.03 ![]() l
l ![]() l2 .31 (n=13) en todo momento de la antesis. El patrón de secreción y el efecto de las extracciones se muestran en la Fig. 3. La secreción es continua, en la fase 2 (8 hs) el volumen es considerablemente mayor que en las flores recién abiertas, mientras que en la fase 3 es muy escaso. El volumen menor a las 10 hs que a las 8 hs se debe a la gran variabilidad entre flores respecto a este parámetro. Las flores sometidas a extracciones sucesivas repusieron semejante volumen (aunque menor) que las que no recibieron este tratamiento.
l2 .31 (n=13) en todo momento de la antesis. El patrón de secreción y el efecto de las extracciones se muestran en la Fig. 3. La secreción es continua, en la fase 2 (8 hs) el volumen es considerablemente mayor que en las flores recién abiertas, mientras que en la fase 3 es muy escaso. El volumen menor a las 10 hs que a las 8 hs se debe a la gran variabilidad entre flores respecto a este parámetro. Las flores sometidas a extracciones sucesivas repusieron semejante volumen (aunque menor) que las que no recibieron este tratamiento.
Visitantes: en 1995, casi no acudieron insectos a las flores de P. foetida. En 1996, la sequía temporaria causada por las
elevadas temperaturas, a pesar de las recientes lluvias (Gonzáles, com. pers.), aparentemente provocó una disminución en la abundancia de insectos; sin embargo, P. foetida recibió visitas. A continuación, se detalla el comportamiento de los visitantes en las flores:
Hymenoptera: generalmente recorrían 1 ![]() 1 flores/planta en cada visita (n= 34) (1-4).
1 flores/planta en cada visita (n= 34) (1-4).
Colletidae
Ptiloglossa tarsata (Friese 1900) (machos y hembras): las visitas eran muy rápidas. Se posaban sobre los radii con el cuerpo paralelo a éstos y libaban separando el operculum del limen. Contactaban las anteras, y los estigmas en fase 2. El polen se depositaba sobre el dorso del tórax. Las hembras a veces se lo pasaban a las patas III. Los machos volaban hasta 15 min. alrededor de las plantas, probablemente a la espera de hembras; no siempre libaban.
Caupolicana sp.(machos): se posaban con el cuerpo paralelo a los radii y separaban el operculum del limen para libar, contactando las anteras con el dorso del tórax.
Apidae
Peponapis fervens (Smith 1879) (machos): se posaban sobre los radii y libaban separando el operculum del limen, contactando las anteras con el dorso del tórax.
Apis mellifera L. 1758: se posaban sobre los radii e intentaban libar pero no lo lograban; se colgaban de las anteras para recolectar polen, impregnándose con éste el dorso del tórax, patas y zona ventral del abdomen.
Halictidae
Pseudaugochloropsis sp.: se posaban sobre los radii, perforaban el limen en varios puntos y libaban todo el néctar. Se embadurnaban con polen el dorso del tórax y alas; a veces se lo removían después de la visita, sobre plantas adyacentes. Contactaban los estigmas ocasionalmente.
Augochlorella sp.: recolectaban polen colgándose de las anteras, quedando éste en las patas y la zona ventral del abdomen; a veces perforaban el limen o intentaban libar, extendiendo el aparato bucal junto a la base del androginóforo o entre limen y operculum.
Dialictus sp., Halictillus sp.: intentaban libar extendiendo el aparato bucal entre limen y operculum, pero no podían introducirlo; en la mayoría de las visitas recolectaban polen colgándose de las anteras, quedando éste en las patas y zona ventral del abdomen.
Vespidae: inspeccionaban entre los radii.
Lepidoptera, Sphingidae: un ejemplar libó una flor el 18/I/96 a las 6.10 hs.
Coleoptera, Chrysomelidae, Diabrotica speciosa: comían polen y también el limen y el operculum.
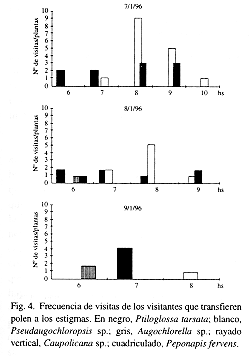
En la Fig. 4 se registran las frecuencias de trabajo en las flores de los visitantes que transfieren polen a los estigmas. Los más frecuentes y constantes fueron Ptiloglossa tarsata, Pseudaugochloropsis sp. y Augochlorella sp. En el Cuadro 2 se detalla la carga polínica de los visitantes en los que se encontró polen.
Relación entre las fases florales, frecuencia de visitas y condiciones meteorológicas (cfr. Figs. 2, 4 y 5): entre las especies más frecuentes y constantes, los individuos de P. tarsata visitaban las flores durante las fases 1 y 2 principalmente, mientras que los de Pseudaugochloropsis sp. y Augochlorella sp. lo hacían durante las fases 2 y 3. Las condiciones meteorológicas del día 9 provocaron una disminución en la diversidad de visitantes, sin la concurrencia de los insectos más pequeños.
Relación entre las medidas de las flores y de los visitantes (Cuadro 3): la superficie que la flor ofrece para posarse (radii) es suficiente para cualquiera de los visitantes. P. tarsata, Caupolicana sp., P. fervens y A. mellifera pueden contactar las anteras con el dorso del tórax. A veces, debido a sus movimientos, Pseudaugochloropsis sp. también lo hace. Si bien todos los visitantes medidos alcanzan el fondo de la cámara nectarífera, es necesaria determinada fuerza para abrirla. P. tarsata, Caupolicana sp. y P. fervens accedían legítimamente al néctar; también Pseudaugochloropsis sp., cuando no perforaba el limen.
| | ||||
| Especie | | | ||
| Ptiloglossa tarsata | Dorso de cabeza, tórax y abdomen, patas II y III | 100 P. foetida | ||
| Caupolicana sp. | Abdomen (V) y patas III | 100 P. foetida | ||
| Peponapis fervens | Patas III | 100 P. foetida | ||
| Pseudaugochloropsis sp.
| Abdomen (V) Patas III Patas II | 87 Solanum spp. 13 P. foetida 99.8 Solanum spp. 02. P. foetida 100 P. foetida | ||
| Augochlorella sp.
| Abdomen (V) Patas III y costados tórax | 91 P. foetida 9 Solanum spp. + Lythraceae 100 P. foetida | ||
| Dialictus sp. | Patas III | 100 Solanum sp. | ||
| Halictillus sp. | Abdomen (V) y patas III | 100 Solanum sp. | ||
*=no se pudo medir.
Discusión
Sistema reproductivo: según el índice P/O, P. foetida sería autógama facultativa-xenógama facultativa. La obtención de semillas en todos los tratamientos indica que existe un importante grado de autogamia (principalmente cuando se induce) y de xenogamia. El porcentaje de semillas producidas por autogamia espontánea fue menor que en el resto de los tratamientos. En condiciones naturales, el escaso polen propio en el borde de los estigmas (depositado al girar las anteras cuando las flores abren o cuando los estigmas alcanzan la altura de aquéllas) no interfiere espacialmente con el polen transferido por los polinizadores. Por lo tanto, los frutos tendrán semillas producto de autogamia espontánea y xenogamia. Si una flor no recibe visitas, al menos algunas semillas serían producidas por autogamia. Por lo tanto, P. foetida sería xenógama facultativa. El porcentaje de frutos obtenido por xenogamia inducida fue menor que por polinización libre, probablemente debido al desconocimiento de la relación óptima cantidad de granos de polen/óvulos. No parece haber competencia por los tubos polínicos entre placentas ya que el número de semillas por placenta es similar en cada fruto, pero sí entre óvulos de la misma placenta, pues generalmente son fecundados los apicales, como ocurre en P. mooreana (Amela García & Hoc en prensa).
Reservas y viabilidad polínicas: los visitantes que recolectan el polen de P. foetida (en forma directa o removiéndolo de las partes del cuerpo donde es depositado "accidentalmente" y pasándolo a las patas III) son recompensados por sus reservas, de gran valor nutritivo para sus larvas. La alta viabilidad del polen indica que no existiría esterilidad masculina y que es posible implementar su uso en cruzamientos.
Antesis: Janzen (1968) y Gottsberger et al. (1988) observaron que la antesis comenzaba cuando aún estaba oscuro, a diferencia de lo que ocurría en Charata; es probable, entonces, que esté más influenciada por la temperatura y la humedad ambientales que por las condiciones de luz. La duración de la antesis coincide con la observada por Janzen (1968) pero no con la observada por Gottsberger et al. (1988), que empieza y termina antes. Es posible que hayan trabajado en días con más variabilidad de los factores metereológicos, dado que se prolongó en días fríos y nublados; en la localidad chaqueña, en cambio, sólo la lluvia intensa influyó, al empujar las anteras y los estigmas hacia abajo. La sincronización en la apertura concuerda con las observaciones de ambos trabajos.
Fases florales: se reconocen las mismas que para P. mooreana (Amela García & Hoc en prensa): 1 y 3, dadoras de polen; 2, dadora y receptora de polen. Sin embargo, la duración de las fases (sobre todo de la 1) es menor (debido a que la antesis es más breve) y,en la fase 3, los estilos quedan recurvados y la pérdida de turgencia es más notoria (probablemente por la consistencia más tenue de los ciclos).
Atractivos: P. foetida no presenta gran cantidad de flores (igual que P. lutea, Neff & Rozen 1995) como atractivo a larga distancia, a diferencia de otras especies más conspicuas de Passiflora (Faegri & van der Pijl 1979). El limen y el androginóforo (en contraste con los estambres y el gineceo) constituirían una guía de néctar a corta distancia en el espectro UV. El olor de la corona es también un atractivo. El olor de las brácteas, probablemente producido por las numerosas glándulas que poseen, podría ser disuasivo de florívoros; Durkee et al. (1984) detectaron una sustancia resinosa secretada por ellas.
Néctar: registra un máximo durante la fase 2 (dadora y receptora de polen), que es la más visitada. Las extracciones sucesivas provocan menos reposición que en P. mooreana (Amela García & Hoc en prensa), probablemente en relación con la antesis más breve.
Síndrome floral: por la antesis diurna, color en el espectro visible, guías de néctar en el especto UV, olor fresco, superficie de aterrizaje, acceso al néctar a través de una entrada que debe reconocerse y forzarse, recompensa oculta no muy profundamente y néctar con alta concentración de azúcares, se encuadra en el síndrome melitófilo descripto por Faegri & van der Pijl (1979), aunque es actinomorfa, los órganos sexuales están expuestos y el néctar es abundante. Por sus dimensiones está adaptada a ser polinizada por himenópteros medianos.
Visitantes: para el lapso de floración estudiado, puede interpretarse el papel de cada taxon visitante como sigue: P. tarsata: por su constancia, sus visitas a flores en fase 1 y 2, su contacto de las anteras y los estigmas, y el gran porcentaje de polen de P. foetida que transportan en partes corporales que contactan los estigmas, los individuos de esta especie son muy importantes polinizadores. Caupolicana sp. y P. fervens: por su baja frecuencia, la libación de flores en fase 1, su contacto con las anteras en dicha fase y el gran porcentaje de polen de P. foetida que transportan en partes del cuerpo que no contactan los estigmas, hurtarían néctar y polen. A. mellifera: por su baja frecuencia, su recolecta de polen en flores en fase 1 y 2, su incapacidad para acceder al néctar y su falta de contacto con los estigmas, hurtarían polen, como también lo observaron Gottsberger et al. (1988). Pseudaugochloropsis sp.: por su alta frecuencia y constancia, sus visitas a flores en fase 2 y 3, las perforaciones que realizan en el limen, su contacto con el polen y transferencia ocasional a los estigmas, serían principalmente ladrones primarios de néctar, hurtadores de polen, y raramente polinizarían; la especie observada por Gottsberger et al. (1988), en cambio, no contactaba anteras ni estigmas. Augochlorella sp.: por su moderada frecuencia y constancia, sus visitas a flores en fase 2 y 3, su recolecta abundante de polen, la perforación eventual del limen y su falta de contacto de los estigmas, actuarían principalmente como hurtadores de polen y, ocasionalmente, como ladrones primarios de néctar. Dialictus sp. y Halictillus sp.: por su baja frecuencia, la escasa recolecta de polen (colectaban en el campo pero a los ejemplares capturados y examinados no se les encontró polen de P. foetida), sus intentos fallidos en busca de néctar y su falta de contacto con los estigmas, serían hurtadores de polen poco relevantes. Vespidae: probablemente buscaban insectos entre los radii. Sphingidae: probablemente hurten néctar. Diabrotica speciosa: serían ladrones primarios de polen y florívoros.
En nuestra área de estudio, cuando las flores empezaban a marchitarse (fase 3), todavía recibían visitas (aunque menos frecuentes y sólo de Pseudaugochloropsis sp. y Augochlorella sp.), a diferencia de lo observado por Gottsberger et al. (1988).
Eficiencia en el transporte de polen : como la antesis es breve, la transferencia del polen entre las plantas de P. foetida debe ser rápida. Los individuos de P. tarsata parecen ser eficientes; a pesar de su baja frecuencia, probablemente debida a la fumigación indiscriminada que se realiza en esa región agrícola desde 1994 (González, com. pers.), se obtuvo un alto porcentaje de frutos mediante polinización libre. Por esto, un cultivo de P. foetida donde habite P. tarsata no necesitaría de polinización manual. Según el manejo que se desee realizar, se podrá inducir la autogamia o no.
Grado de adaptación entre la planta y sus visitantes: el síndrome melitófilo para abejas medianas concuerda con que los polinizadores sean los individuos de P. tarsata. Janzen (1968) cita individuos de varias especies de Ptiloglossa como polinizadores específicos de P. foetida en América Central, coincidiendo con el presente estudio el uso de las flores como fuente de néctar por ambos sexos, su período de actividad y comportamiento en ellas, el contacto de anteras y estigmas, y las medidas de las flores y de la altura del tórax de estos insectos. Aunque también a pleno día, este colétido se encuentra activo en horas de poca luz (Roig Alsina, com. pers.), lo que coincide con la temprana apertura de las flores. Por todo esto, existiría cierto grado de adaptación entre P. foetida y estas especies de Ptiloglossa. Los individuos de este género fueron los únicos polinizadores registrados en Charata y en las localidades donde trabajó Janzen (1968); Gottsberger et al. (1988), en cambio, observaron especies de Centris y Xylocopa polinizando. El resto de los visitantes que se vieron durante este estudio, serían oportunistas; si bien la actividad principal de la mayoría de ellos es el hurto o el robo, no causarían importantes efectos negativos a la planta, ya que el porcentaje de frutos con semilla obtenido por polinización libre fue elevado.
Agradecimientos
Agradecemos a José Luis Telma por su ayuda en el campo. A Oscar González por el hospedaje y apoyo brindado. A Arturo Roig Alsina por la determinación de los himenópteros y los útiles comentarios sobre su biología. Al CONICET, por el subsidio N° 548, con parte del cual se financió este estudio.
Resumen
Un experimento reproductivo muestra que Passiflora foetida es autocompatible. Observaciones de las características florales y de los visitantes durante la antesis, más el análisis del polen transportado, permitieron identificar el síndrome floral (melitofilia) y las funciones de cada visitante. La antesis ocurre desde las 6 hasta las 11 hs. Se identificaron 3 fases florales: 1) estigmas por encima de las anteras, 2) estigmas a la altura de las anteras, 3) estigmas por encima de las anteras; los radii, los pétalos y los sépalos se incurvan. Los estigmas están receptivos durante toda la antesis. La concentración de azúcares del néctar es 34 %. El color predominante en el espectro visible es el blanco. En el espectro UV, los estambres y el gineceo contrastan con el limen y el androginóforo; pueden ser una guía de néctar. Tres especies de himenópteros fueron los visitantes más frecuentes y constantes: Ptiloglossa tarsata (Colletidae) siempre contactan las anteras y los estigmas cuando liban, transportan un alto porcentaje de polen de P. foetida y visitan flores en fase 1 y 2; pueden ser considerados los principales polinizadores. Pseudaugochloropsis sp. (Halictidae) raramente contactan las anteras o los estigmas cuando perforan el limen para acceder al néctar y visitan flores en fase 2 y 3; son ladrones de néctar que raramente polinizan. Augochlorella sp. (Halictidae) recolectan polen sin tocar los estigmas y visitan flores en fase 2 y 3; son hurtadores de polen.
Referencias
Amela García, M. T. & P. S. Hoc. 1997. Aspectos de la biología floral y el sistema reproductivo de Passiflora mooreana. Darwiniana (en prensa). [ Links ]
Anónimo. 1989. Diccionario manual e ilustrado de la lengua española. Real Academia Española. Espasa-Calpe, Barcelona. 1666 p. [ Links ]
Burkart, A. 1963. Antiguos herbarios argentinos en Gran Bretaña y su interés fitogeográfico. Darwiniana 12: 533-558. [ Links ]
Cruden, R. W. 1977. Pollen-ovule ratios: a conservative indicator of breeding systems in flowering plants. Evolution 31: 32-46. [ Links ]
Durkee, L. T., C. W. Baird & P. F. Cohen. 1984. Light and electron microscopy of the resin glands of Passiflora foetida (Passifloraceae). Amer. J. Bot. 71: 596-602. [ Links ]
Engstrand, L. 1980. Passionsfrukt (Passiflora edulis). Svensk Bot. Tidskr. 74: 5-8. [ Links ]
Faegri, K. & L. v. der Pijl. 1979. The principles of pollination ecology. Pergamon, Oxford. 244 p. [ Links ]
Greissl, R. 1989. Vitality analysis of monadic and polyadic pollen grains using optical contrast-fluorescence microscopy. Sci. Tech. Inform. 15: 180-184. [ Links ]
Gottsberger, G., J. M. F. Camargo & I. Silberbauer-Gottsberger. 1988. A bee-pollinated tropical community: the beach dune vegetation of Ilha de Sao Luís, Maranhao, Brazil. Bot. Jahrb. Syst. 109: 469-500. [ Links ]
Holmgren, P. K., N. H. Holmgren & L. C. Barnett. 1990. Index Herbariorum. Part I: The Herbaria of the World: 1-693. Regnum Vegetabile 120. New York Botanical Garden, Nueva York. [ Links ]
Inouye, D. W. 1980. The terminology of floral larceny. Ecology 6l: 1251-1253. [ Links ]
Janzen, D. H. 1968. Reproductive behaviour in the Passifloraceae and some of its pollinators in Central America. Behaviour 32: 33-48. [ Links ]
Johansen, D. A. 1940. Plant microtechnique. McGraw Hill, Nueva York. 523 p. [ Links ]
Jones, C. E. & R. J. Little. 1983. Handbook of experimental pollination biology. S&AE, Nueva York. 558 p. [ Links ]
Kelly, K. L. 1965. Inter-Society Color Council- National Bureau of Standards color-name charts illustrated with centroid colors. Standard sample N° 2106 Suppl. to Nat. Bur. Standards Cir. 553. U. S. Government Printing Office, Washington, D. C. 26 p.
Killip, E. P. 1938. The American species of Passifloraceae. Field Mus. Nat. Hist. Bot. Series 19: 1-613. [ Links ]
Martin, F. W. & H. Y. Nakasone. 1970. Edible species of Passiflora. Economic Botany 24: 333-343. [ Links ]
Neff, J. L. & J. G. Rozen. 1995. Foraging and nesting biology of the bee Anthemurgus passiflorae (Hymenoptera: Apoidea), descriptions of its immature stages, and observations on its floral host (Passifloraceae). Amer. Mus. Novitates 3138: 1-19. [ Links ]
Osborn, N. M., P. G. Kevan & M. Lane. 1988. Pollination biology of Opuntia polyacantha and Opuntia phaeacantha (Cactaceae) in southern Colorado. Pl. Syst. Evol. 159: 85-94. [ Links ]
Puri, V. 1948. Studies in floral anatomy, V. On the structure and nature of the corona in certain species of the Passifloraceae. J. Ind. Bot. Soc. 27: 130-149. [ Links ]
Smiley, J. T. 1991. Passiflora foetida (Passifloraceae) (Bombillo, Calzoncillo, Norbo, Granadilla del Monte, Pasionaria, Passion Flower), p. 301-302. In D. H. Janzen (ed.). Historia Natural de Costa Rica. Universidad de Costa Rica, San José. [ Links ]
1 Lab. 11, Depto. Cs. Biológicas, Pab. II, Ciudad Universitaria, 1428, Bs. As. Argentina. Fax: 54-1-7820582. Correo electrónico: amela@biolo.bg.fcen.uba.ar

