











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
Los incendios forestales se presentan en casi todos los tipos de ecosistemas, sin embargo, la capacidad de propagación y extensión está relacionada con la cantidad de humedad y combustible de la vegetación (Mondal y Skumar, 2016; Vargas-Sanabria y Campos-Vargas, 2018). Estos fuegos influyen en procesos y patrones de los ecosistemas como la distribución y la estructura de la vegetación, el ciclo de carbono y el clima (Lasslop et al., 2019; Armenteras et al., 2020).
El cambio climático, incluido las olas de calor, sequías, patrones meteorológicos, la variación de los vientos y las temperaturas de la superficie del mar conocidas como el fenómeno del Niño, pueden aumentar la probabilidad de prevalencia de los incendios forestales (Skinner et al., 2022). Esta variabilidad climática incrementa la frecuencia e intensidad del fuego en algunas partes del planeta, como los incendios ocurridos durante el periodo 2023 al 2024 en Grecia, Canadá, Amazonía y Chile, con importantes repercusiones para la sociedad y el medio ambiente (Jones et al., 2024). Bajo este contexto, es importante generar acciones de prevención y mitigación de los incendios forestales al utilizar como base el conocimiento sobre el comportamiento histórico del fuego, esto permite saber dónde, cuándo y por qué se propaga el fuego en ciertos escenarios (Ramos et al., 2021).
La mayoría de los incendios forestales se originan por actividades antropogénicas (Mora y Esquivel, 2023) y por cambios en el uso del suelo, lo que contribuye al riesgo de fuegos rápidos o intensos, especialmente en los bosques tropicales donde las personas son la principal causa de degradación forestal (Jones et al., 2024; Quesada-Román, y Vargas-Sanabria, 2022). Una de las problemáticas más graves de los incendios es la recurrencia año tras año en un mismo sitio (Vargas-Sanabria y Campos Vargas, 2020), lo que ocasiona una pérdida en la resiliencia de los ecosistemas y provoca alteraciones en los procesos biológicos y de servicios ecosistémicos (Ramos et al., 2021; Vargas-Sanabria y Quesada-Román, 2018).
La distribución espacial de los incendios dificulta el acceso al estudio de áreas afectadas, lo que genera vacíos de información sobre la dinámica del fuego, especialmente en sitios con restricciones especiales como áreas protegidas, territorios indígenas o áreas remotas inaccesibles (Wang et al., 2020). Dicho esto, la teledetección ofrece una valiosa capacidad para recopilar información sobre áreas quemadas de manera rápida, lo cual facilita la creación de cartografía esencial para el registro y seguimiento de los incendios a lo largo del tiempo (Villar- Hernández et al., 2022).
La teledetección es una técnica de observación de la tierra a distancia por medio de imágenes satelitales o radares que permiten obtener datos en tiempo real de la superficie terrestre y se emplea con diversos fines de investigación y monitoreo ambiental (León-Fernández y Garavito-Rincón, 2021). Sabins y Ellis (2020) por su parte, definen la teledetección como la ciencia dedicada a adquirir, procesar e interpretar imágenes y datos obtenidos principalmente desde aviones y satélites, por medio de sensores que capturan digitalmente la interacción entre la energía electromagnética y la materia, permitiendo un análisis detallado de las características de la superficie terrestre. Esta tecnología satelital se ha convertido en una aliada para la generación de información sobre áreas quemadas, sensores como MODIS y VIIRS son utilizados para el monitoreo de incendios forestales a escalas regionales y globales con información casi a tiempo real (Katagis y Gotas, 2022).
El Parque Internacional La Amistad (PILA) sector Costa Rica se ubica en la Cordillera de Talamanca, (Quiñonez-Guzmán et al., 2019). Es el área protegida terrestre más grande del país (Mejía-Agüero y Quesada-Román, 2024) y se extiende a lo largo de 199 147 hectáreas de bosque tropical (SINAC, 2019). Por otro lado, el territorio panameño cuenta con 207 000 hectáreas (Ministerio de Ambiente, 2023) y juntos comparten alrededor de 406 174 hectáreas lo que constituye un puente biológico entre América del Norte y América del Sur, que alberga nueve zonas de vida presentes en Costa Rica (Sánchez-Ocampo, 2024).
El PILA está integrado por diversos ecosistemas como bosque húmedo, sabanas, turberas catalogadas como sitio Ramsar y páramos, con una topografía de difícil acceso, y condiciones climáticas variadas (Artavia-Rodríguez et al., 2024; Mejía-Agüero y Quesada-Román, 2024; Monro et al., 2017; Quiñonez-Guzmán et al., 2019; Roche y Flores, 2017).
En la zona de estudio correspondiente a la zona de amortiguamiento del Parque Internacional La Amistad se han identificado el mayor número de incendios forestales atendidos por las autoridades del SINAC (SINAC, 2017). Aquí destacan comunidades de los pueblos originarios Bribri y Cabécar, distribuidos entre los Territorios Indígenas (T.I. de ahora en adelante) de Ujarrás, Salitre y Cabagra (González-Brenes y Elizondo-Sánchez, 2023), además de población campesina no indígena. En dichas comunidades se desarrollan diversas actividades sociales, económicas, productivas y recreativas, vinculadas con los diversos servicios ecosistémicos derivados de los recursos naturales del entorno inmediato, que podrían influir directa o indirectamente en los recursos del Parque Internacional La Amistad (SINAC, 2019).
El presente trabajo tiene el objetivo de identificar la presencia de incendios forestales en la zona de amortiguamiento del PILA y su relación con variables geográficas que influyen en la ocurrencia de dichos eventos, los cuales, además de actuar directa o indirectamente en los recursos del parque, pueden tener un impacto en las comunidades aledañas, donde se desarrollan actividades sociales, económicas, productivas y recreativas. Por esta razón, es importante identificar zonas prioritarias para implementar estrategias de mitigación y prevención de incendios forestales.
Materiales y métodos
Generación de datos de incendios forestales:
Primero, descargamos los datos en formato vectorial de ESRI tipo Shapefile (SHP) de áreas quemadas mensuales correspondientes al periodo de estudio 2012-2020, desde la plataforma GWIS (Global Wildfire Information System (GWIS), 2023) que provienen del sensor MODIS MCD 64A1 con una resolución global de 500 metros. Utilizamos los datos obtenidos para los meses de enero a abril relacionados con la época seca y la temporada donde ocurren la mayoría de incendios forestales en Costa Rica (Campos- Vargas y Vargas-Sanabria, 2021).
Posteriormente, realizamos una validación de los datos de áreas quemadas proporcionados por el sensor MODIS MCD 64A1 en dos etapas: la primera fue la validación con datos de puntos de calor de los satélites Aqua y Terra del sensor MODIS y VIIRS, los cuales fueron descargados del sitio FIRMS (Fire Information for Resources Management System (FIRMS), 2024) de la NASA. La segunda etapa consistió en validar dichos datos mediante el criterio experto de los funcionarios del PILA, quienes realizaron una interpretación visual de las áreas indicadas como quemadas utilizando el software QGIS versión 3.10.7 y el visor Google Earth Pro.
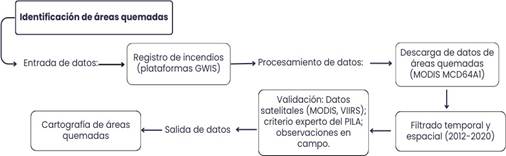
Para identificar las áreas de recurrencia de incendios forestales en la zona de amortiguamiento del PILA, procedimos a convertir los datos de áreas quemadas de formato vector a raster y luego realizamos una sumatoria con las ocho capas correspondientes para cada año (2012 al 2020) utilizando la herramienta estadística de celda del programa ArcGIS 10.4.1. Por último, realizamos una clasificación basada en la metodología empleada por Quesada-Román y Vargas-Sanabria (2022) . Presentamos un diagrama del flujo de los procesos empleados en la metodología para la identificación de las áreas quemadas. (Fig. 1)
Variables geográficas que influyen en la ocurrencia de incendios forestales:
Para identificar las variables geográficas presentes, tomamos como base elementos del entorno propuestos por Moraga (2017) y Galiana et al. (2009) los cuales se adaptaban al área de estudio y consisten en datos de presencia de red vial, elevaciones del área de estudio, presencia de poblados, tipo de vegetación (como elemento de combustible) y pendientes. Adicionalmente, utilizamos la red hídrica como elemento fundamental en el auxilio de las labores de extinción de incendios (Cordeiro et al., 2021). Los datos de cada variable los obtuvimos mediante la descarga en formato vectorial a través del Sistema Nacional de Información Territorial de Costa Rica, por medio de los servicios OGC (Consorcio Geoespacial Abierto) y posteriormente los procesamos en el programa QGIS versión 3.10.7 mediante el servicio de descarga archivo vectorial WFS (Web Feature Server).
Las capas correspondientes a red vial, hidrografía, cobertura de pastos y presencia de poblados se descargaron a escala 1:5000. Las pendientes de la zona de amortiguamiento se generaron mediante la capa de curvas de nivel a escala 1:200.000, y se utilizó la herramienta Topo to Raster del programa ArcGIS 10.4.1. con el cual generamos el modelo de elevación digital (DEM).
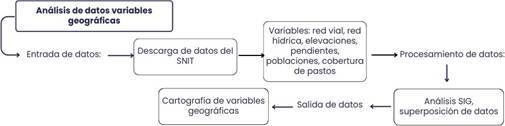
Una vez generada la capa de elevaciones, procedimos a desarrollar en el programa ArcGIS 10.4.1 un modelo de pendientes. Desde la herramienta Pendiente (Slope), se definieron y clasificaron las pendientes en diferentes rangos, seguidamente, se reclasificaron estos rangos mediante la herramienta Reclasificación (Reclassify), en categorías establecidas en la “Metodología para la Determinación de Uso de las Tierras de Costa Rica” la cual es oficial y fue elaborada por el Ministerio de Agricultura y Ganadería en 1991 bajo el Decreto N° 23214-MAG- MIRENEM. Mostramos el flujo de procesos empleados para la identificación de variables geográficas del entorno. (Fig. 2)
Resultados
Identificación de áreas quemadas por incendios forestales:
Se logró identificar en la zona de amortiguamiento, perímetros de áreas quemadas en los sectores Territorio Indígena Ujarrás, Territorio Indígena Salitre y Territorio Indígena Cabagra (Fig. 3). En los sectores Altamira, Biolley y Las Tablas no se detectaron incendios durante el periodo de estudio. La utilización de puntos de calor MODIS y VIIRS permitió validar que las áreas identificadas como quemadas mediante el sensor MODIS MCD 64A1 sí están relacionadas a actividad de incendios, lo cual también fue corroborado por el personal del parque nacional (Fig. 4). Se registraron un total de 30 370.59 hectáreas afectadas durante el periodo de estudio, siendo el sector de T.I Ujarrás el más afectado con un total de 24 617.52 hectáreas; mientras que T.I Salitre, con 302.73 hectáreas quemadas, presenta el menor grado de afectación.
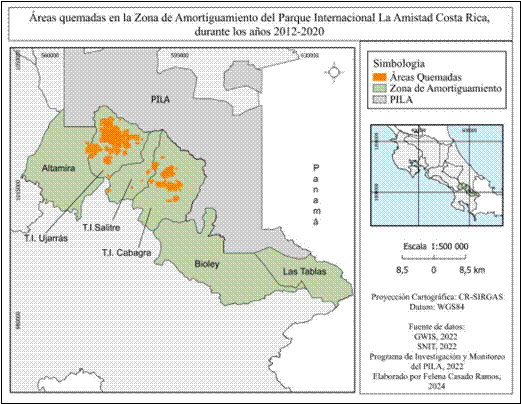
Fig. 3 Áreas quemadas detectadas durante los años 2012 - 2020 en la Zona de Amortiguamiento del Parque Internacional La Amistad.
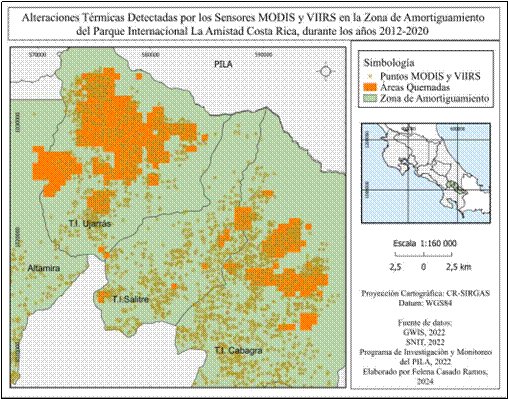
Fig. 4 Puntos de Calor detectados por los sensores MODIS y VIIRS durante los años 2012-2020 en las áreas quemadas de la Zona de Amortiguamiento del Parque Internacional La Amistad.
Por otro lado, definimos tres categorías de recurrencia de incendios forestales: un incendio, dos incendios y tres incendios. Durante el periodo 2012-2020, las zonas con recurrencia de al menos tres incendios se localizan en el territorio T.I Ujarrás, mientras que las zonas con presencia de al menos dos incendios se encuentran tanto en T.I Ujarrás y T.I Cabagra. Por último, las zonas donde ha habido al menos un incendio en los últimos ocho años corresponden a T.I Ujarrás, T.I Salitre y T.I Cabagra (Fig. 5)
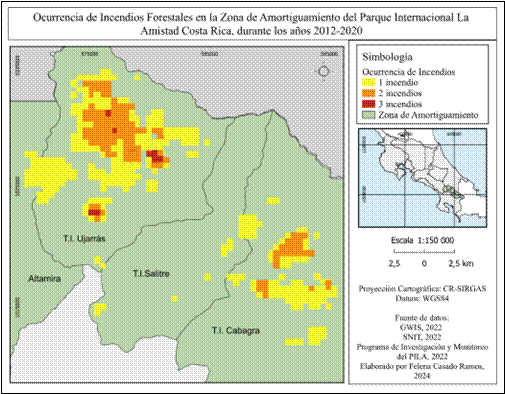
Áreas quemadas y variables geográficas presentes:
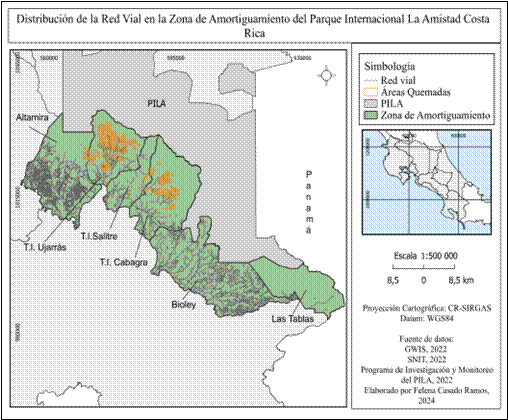
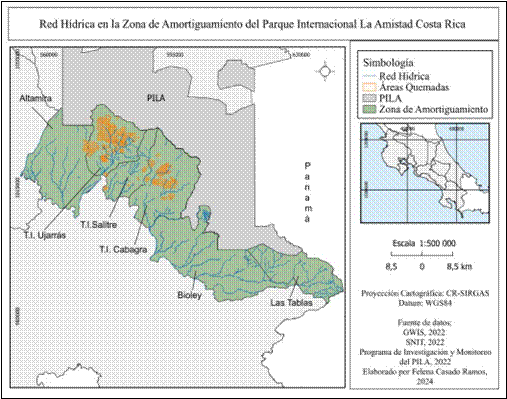
Los datos geoespaciales muestran mayor presencia de red vial (Fig. 6) en zonas de altitudes bajas y lejanas a los límites del parque, el sector de Altamira aglomera la mayor cantidad de vías, contrario a T.I Salitre con menor presencia de red vial. La red hídrica (Fig. 7), por su parte, se encuentra distribuida en toda la zona de amortiguamiento, con alta concentración de cuerpos de agua en la parte alta de Biolley y en la zona de T.I Ujarrás y menor presencia de agua en las partes altas y más próximas al Parque Internacional La Amistad, correspondientes a los sectores de Altamira, T.I Ujarrás, T.I Salitre y T.I Cabagra.
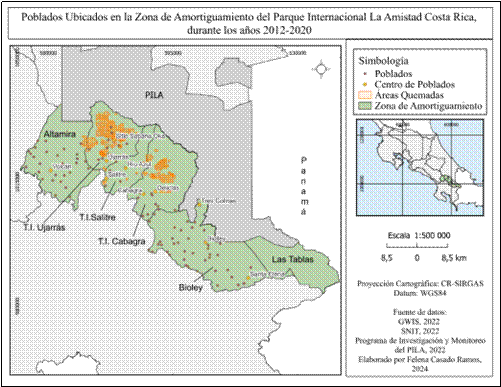
En cuanto a la variable de poblados (Fig. 8), se contabilizaron 106 poblados a lo largo del área de amortiguamiento, con excepción de la zona de Las Tablas en donde los datos no indican presencia de los mismos. Biolley es la zona más poblada con 45 asentamientos, seguidamente Altamira con 30 asentamientos, T.I Ujarrás 13, T.I Cabagra 11 y T.I Salitre presenta solamente siete poblados.
La concentración poblacional en Altamira, T.I Salitre y T.I Cabagra se ubica principalmente en zonas bajas y alejadas de los límites del Parque Internacional La Amistad. Los poblados de T.I Ujarrás se encuentran en zonas más próximas a los límites del parque y en el caso de T.I Cabagra, la población está dispersa de manera más homogénea por toda la zona. Destacan seis poblados localizados muy cerca de los límites del parque: Tres Colinas, Helechales, Sitio Sabanas, Tierras Negras, La Puna y Altamira.
Con respecto a la vegetación, la cobertura de pastos (Fig. 9) se extiende a lo largo de toda la zona de amortiguamiento, con excepción del sector de Las Tablas donde la cobertura es mínima, mientras que en Altamira se observan parches aislados de pastos distribuidos por toda el área. En T.I Ujarrás, se observa una amplia y densa cobertura de pastos en casi la totalidad del área, excepto en las partes altas más próximas a los límites del parque y la zona limítrofe con Altamira.
En T.I Cabagra, la cobertura de pastos también es extensa y se ubica lejos del Parque Internacional La Amistad. En T.I Salitre los pastos se extienden por toda la zona, con mayor concentración en las partes bajas alejadas del Parque Internacional La Amistad, conforme el área se acerca a los límites del parque, la cobertura de pastos se mantiene en pequeños parches. El sector de Biolley también alberga una extensa cobertura de pastos en toda el área, exceptuando la parte central y algunos parches muy cercanos al límite con el Parque Internacional La Amistad.
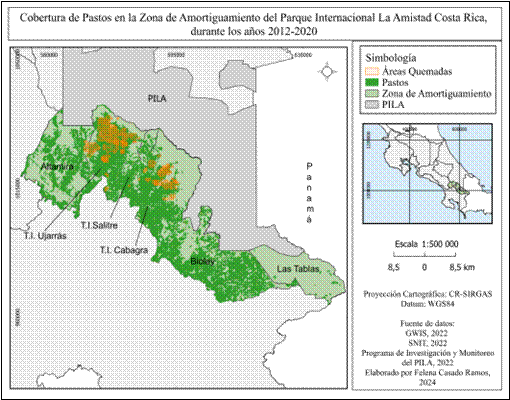
Fig. 9 Cobertura de pastos presente en la Zona de Amortiguamiento del Parque Internacional La Amistad.
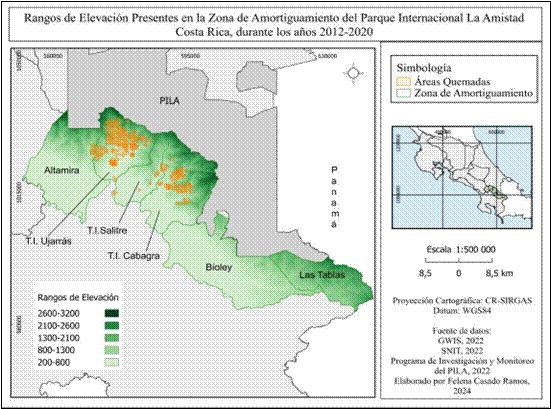
En relación a la topografía de la zona de amortiguamiento, el modelo de elevación digital elaborado (DEM) muestra diversos rangos que oscilan entre 200 m.s.n.m. hasta 3200 m.s.n.m (Fig. 10). Las mayores alturas están representadas con rangos que van desde los 1600 m.s.n.m., y se concentran en sectores cercanos a los límites del Parque Internacional La Amistad: Altamira,T.I Ujarrás, T.I Salitre y T.I Cabagra, a excepción del sector de Las Tablas, cuyas zonas altas se encuentran próximas a la frontera con Panamá. El sector con menor rango de elevación según el modelo es Biolley que va de 200 m.s.n.m a 1900 m.s.n.m.
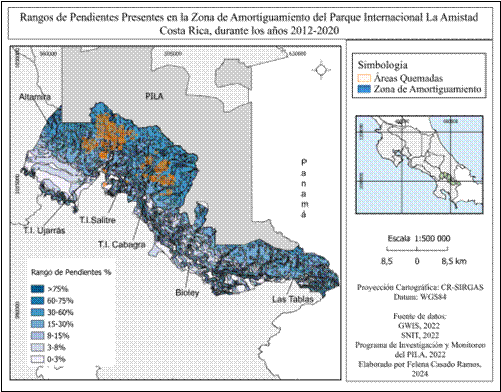
La topografía muestra áreas muy escarpadas o de mucha pendiente (Fig. 11), en la zona de Altamira, la más alejada del Parque Internacional La Amistad, predominan pendientes entre 60-75% y 8 -15%, en las zonas más próximas al parque de 30-60% y de 15-30%. En T.I Ujarrás, predominan las pendientes de 30-60% y de 15-30% las más cercanas a los límites del área protegida. En T.I Salitre y T.I Cabagra también predominan las pendientes de 30-60% y de 15- 30%, además se observan pendientes entre 0-3% y 60-75%.
En el sector de Biolley se observan mayor cantidad de pendientes entre los rangos 0-3%, 8-15% y 60-75%, y se observan pendientes de 30-60% en la parte más cercana a los límites del Parque Internacional La Amistad. Se observa una mayor cantidad de rangos de pendiente entre 30-60% en Las Tablas y pocas pendientes mayores a 75%, en las partes altas más próximas al Parque Internacional La Amistad en los sectores de Altamira, T.I Ujarrás y T.I Cabagra.
Discusión
Los incendios forestales en la zona de amortiguamiento del PILA durante el periodo 2012 al 2020, se localizaron en Ujarrás, Cabagra y Salitre, que corresponden a territorios indígenas. En esta zona predomina una alta cobertura boscosa así como áreas con presencia de pastizales (Quesada-Román et al., 2023) las cuales suelen ser quemadas para limpieza de terrenos o cambio de uso del suelo propiciando en muchos casos la ignición de incendios debido a un mal manejo del fuego (Martínez, 2022). Asimismo, Artavia-Rodríguez et al. (2024) sugieren que los fuegos en estos territorios son originados por acciones humanas como cacería, vandalismo y actividades agrícolas que suelen salirse de control.
Roche y Flores (2017) indican que el incremento de los incendios en los últimos años se debe a ganaderos no indígenas, que han utilizado la quema para expandir sus áreas de potreros. De acuerdo a Martínez (2022) los incendios en las zonas de T.I Salitre y T.I Cabagra suelen presentar una severidad alta debido a la presencia de cobertura de pastizal lo que provoca una fácil propagación del fuego.
Por lo general, el 91.5% de los incendios identificados (279 T.I Ujarrás y 130 en T.I Cabagra) ocurren cerca de asentamientos humanos, con características geográficas y topográficas complejas como acceso vial, disponibilidad de agua y restricciones de ingreso a los territorios indígenas, lo que dificulta la labor de prevención, control y manejo del fuego. Esto representa una amenaza para la biodiversidad del PILA; y también para los elementos fundamentales de la cosmovisión indígena y los recursos que utilizan para su subsistencia.
Walker et al. (2022) menciona que la relación entre la ocurrencia de incendios y las tierras indígenas puede ser compleja por temas de autonomía o legislación y que en otros estudios como el desarrollado por Fonseca et al. (2020), se han registrado dinámicas complicadas de los incendios dentro de estos territorios, lo que puede afectar áreas protegidas si no se realiza un control apropiado.
La ocurrencia de incendios en estos tres territorios indígenas, define una dinámica del fuego que se presenta a partir de los 1600 m.s.n.m. a excepción de algunos incendios registrados desde los 500 m.s.n.m. en la parte baja de T.I Salitre. La mayoría de incendios se registran entre los rangos de pendiente de 30-60% y 60-75% (ondulado y escarpado), y en las zonas bajas con rangos de pendiente de 0-30% (plano o casi plano). Galiana et al. (2009) mencionan que las áreas con una pendiente mayor a 30% presenta una alta peligrosidad en la propagación de los incendios lo que influye en la atención temprana de los brigadistas debido a las condiciones topográficas. Vargas-Sanabria y Quesada-Román (2018) sugieren que la propagación del fuego generalmente está asociada a pendientes pronunciadas y fuertes vientos, dado que existe una estrecha relación entre la geomorfología y el fuego. Asimismo, Elizondo menciona en el Plan de Manejo del Fuego del PILA (2013), que las pendientes juegan un rol fundamental en la gestión del fuego y señalan tres sitios importantes en la parte alta del sector T.I Ujarrás que son: Dúrika, Sabanas Oka y Yuavin, donde han ocurrido la mayoría de incendios forestales afectando ecosistemas de bosques, sabanas y charrales.
Durante el desarrollo de este estudio, el sector de T.I Ujarrás ha registrado la mayor cantidad de áreas quemadas y puntos recurrentes de incendios forestales entre 2012 y 2020, lo mismo fue reportado por Artavia-Rodríguez et al. (2024). Este territorio se ha identificado como una zona crítica para las brigadas de incendios, ya que representa una amenaza significativa para los recursos naturales del parque debido a su proximidad al límite con el área protegida. En 2015, se evidenció la mayor extensión de tierras afectadas por el fuego, alrededor de 13 001.73 hectáreas, y la influencia de eventos hidrometeorológicos como el Fenómeno del Niño (ENOS) (Pérez-Bertozzi et al. 2024) intensificó los incendios debido a las sequías y altas temperaturas durante ese período.
Esta recurrencia de incendios ha impulsado la organización de grupos locales para la promoción de prácticas sostenibles, educación ambiental y brigadas forestales con el fin de proteger los recursos naturales del PILA tanto del lado costarricense como panameño (Roche y Flores, 2017). De acuerdo a Mora y Esquivel, (2023) estas actividades propiciadas por personas de las comunidades han permitido fortalecer los planes de manejo del fuego e implementar acciones de prevención y control con el fin de detener la degradación forestal en la zona, producto de los incendios forestales.
Por último, es importante resaltar la importancia de las herramientas tecnológicas como los satélites para la generación de datos sobre áreas afectadas por incendios forestales. Esta tecnología permitió la identificación de alrededor de 447 incendios forestales ocurridos durante el periodo de estudio , información relevante para apoyar la planificación y toma de decisiones a las entidades responsables de la gestión integral del fuego del Parque Internacional La Amistad.
Ethics, conflict of interest, and funding statement
Declaramos, Felena Casado Ramos y Daniela Vargas Sanabria, haber cumplido con todos los requisitos éticos y legales pertinentes, tanto durante el estudio como en la preparación de este documento; que no hay conflictos de interés de ningún tipo, y que todas las fuentes financieras se detallan plena y claramente en la sección de agradecimientos. Asimismo, estamos de acuerdo con la versión editada final de esta publicación. El respectivo documento legal firmado se encuentra en los archivos de la revista.
La declaración de contribución de cada autor es la siguiente: F.C.R., D.V.S.: Diseño del estudio, recolección y análisis de datos. Todos los coautores: preparación y aprobación final del manuscrito.

