











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
Permalink
Introduction
The genus Dendrophylax Rchb.f. (Epidendroideae: Vandeae: Angraecinae), commonly known as “ghost orchids”, currently consists of 15 species distributed primarily in the Greater Antilles but with two species that have ventured north to Florida (USA) (D. lindenii (Lindl.) Benth. ex Rolfe, D. porrectus (Rchb.f.) Carlsward & Whitten), and two occur in Megamexico (D. megarhizus Molgo & Carnevali, D. porrectus). Dendrophylax and its sister genus, Campylocentrum Benth., are the only two representatives of the subtribe Angraecinae in the western hemisphere.
Dendrophylax is epiphytic yet leafless at maturity, rendering them inconspicuous. Flowers are borne singly or on laxly flowered racemes or sparsely branched panicles (Campylocentrum has distichous, densely flowered racemes). Flowers are yellow, green, or white, spurred, and produce nectar as a pollinator reward. The stubby column has a split rostellum and the anther bears two independent, yellow pollinia with separate stipes (hemipollinaria). The larger species have spectacular white, nocturnally fragrant flowers that seem to float in the understory as they are usually borne alone at the end of long, slender inflorescences; hence the name “ghost orchids” (Ackerman & Collaborators, 2014).
The center of Dendrophylax diversity is Hispaniola, which is home to 11 species, five of which are endemic. This matches the overall percent endemism for orchids on the island (44% for all orchids; 45% for Dendrophylax) (Ackerman & Collaborators, 2014). High endemism is not only characteristic of the orchid flora of Hispaniola, but also prevalent across many plant and animal groups making Hispaniola a major contributor to the Caribbean biodiversity hotspot (e.g., Powell et al., 1999; Santiago-Valentín & Olmstead 2004). Its geological diversity, physiographic and climatically fragmented landscape offers opportunities for population isolation where genetic drift and natural selection may occur without homogenizing effects of gene flow (Cano Carmona et al., 2010; Cano Ortiz et al., 2016; Hu et al., 2022; Tremblay et al., 2005).
While it is tempting to regard the Hispaniola as the evolutionary origin of the Dendrophylax, the phylogenetic evidence is equivocal. The earliest diverging clades have the most widespread species occupying multiple islands of the Greater Antilles: D. monteverdi (Rchb.f.) Ackerman & Nir (Puerto Rico, Hispaniola, Jamaica, and Cuba), D. barrettiae Fawc. & Rendle (Hispaniola, Jamaica, and Cuba), and D. porrectus (Puerto Rico, Hispaniola, Jamaica, Cuba, Florida, and Mexico) (Ackerman & Collaborators, 2014; Carlsward et al., 2006; Molgo et al., 2016). As for diversification of Dendrophylax, it likely diverged from Campylocentrum in the late Miocene (Farminhão et al., 2021). Unfortunately, diversification within Dendrophylax lacks clarity since only three morphologically disparate species of Dendrophylax have been assessed, and they diverged in the early to mid-Pliocene (Pessoa et al., 2018). Nonetheless, the phylogram depicting Dendrophylax phylogeny of Carlsward et al. (2006) showed short Fitch lengths, suggesting recent divergence of sister species occupying different islands (D. sallei of Hispaniola and D. lindenii of Cuba and Florida; D. fawcettii of the Cayman Islands and D. funalis of Jamaica).
We encountered a population of a small flowered Dendrophylax that we have concluded is an undescribed species, which we rectify here.
Taxonomic treatment
Dendrophylax pustulatus Ackerman & E.Fernández, sp. nov. (Fig. 1-2)
TYPE: República Dominicana. Cordillera Central: provincia Santiago, a 1 km en dirección este por el sendero, después de la caseta del Parque Nacional Armando Bermúdez, La Diferencia, frente a La Diferencia Eco-Retreat. A lo largo de la vegetación contigua al Río Amina. En pequeñas poblaciones a lo largo del sendero. Pico floral parece ser entre principios y mediados de Julio, 19°16”13”N, 71°03’07”W, elev. 735 m, 12 julio 2024, E. Fernández 23 (holotype: JBSD 138476; isotype: NY). Fig. 1-2.
Diagnosis: Dendrophylax pustulatus differs by a combination of characteristics from other small-flowered and short-spurred members of the genus, D. serpentilingua (Dod) Nir, and D. constanzensis (Garay) Nir. Dendrophylax pustulatus inflorescence has abundant ellipsoid pustules and an obovate retuse mid lobe of the lip, whereas the pubescence of D. serpentilingua is hispid and the mid lobe is ovate, apiculate and two-tailed, and D. contanzense is setaceous-hirsute, and the mid lobe is anchor-shaped, apiculate. Dendrophylax pustulatus also has a laterally compressed spur, a trait that is shared only with D. serpentilingua. Furthermore, D. pustulatus consistently has a 90° bend at the transition between the pedicel and ovary, not yet seen in other species of the genus.
Plant epiphytic, monopodial, leafless. Roots ca. 10 or more, green when wet, white to whitish green when dry, caespitose, 1.0-1.5 mm diam., up to 15 cm long, smooth. Rhizome largely obscured by roots, 5-10 mm long. Inflorescences erect, filiform, racemose or sparsely branched, 1-6 flowers produced in succession, usually 1 open at a time, densely provided with brown elliptical pustules; peduncle green, 17-30 mm long, 0.5 mm diam., comprised of 3 nodes, bracts brown, sheathing, acute, 1.5-2.0 mm long, progressively smaller toward the apex; rachis 5-15 mm long, somewhat fractiflex; floral bract 1.5-2.0 mm long. Flowers yellow to yellow green, in the resupinate position; pedicellate ovary geniculate; pedicel erect, 2.0-2.5 mm long, glabrous or with sparse brown pustules; ovary perpendicular to the pedicel, 1.5-2.0 mm long, ribbed, densely provided with brown pustules; sepals weakly pustulose/verrucose abaxially, entire; dorsal sepal ovate, obtuse, 1.9 × 1.2 mm, reflexed upward 0.5 mm from base; lateral sepals ovate, slightly oblique, acute, entire, 2.8-3 mm long, 1.8 mm wide; petals glabrous, flanking column, ovate, obtuse, 1.8 × 1.2 mm; lip glabrous, 3.0-3.5 mm long, excluding spur, superficially simple, obscurely trilobed, lateral lobes erect, flanking column, rounded, entire, creating a narrow entryway to spur, and meeting the mid lobe at about half the length of the lip, mid lobe obovate, rounded, retuse, ca. 2.0 mm long, 2.4 mm wide, disc callus a low elliptical bump in front of the column 0.7 mm long, spur 1.5-2.0 mm long × 1.0-1.2 mm deep, laterally compressed, truncate, parallel to ovary, abutting against the pedicel; column light green, stubby, 0.5 mm long; anther cap yellow with two brown spots, pollinia 2, globose, ca. 0.16 mm diam., attached to separate stipes and viscidia. Fruit pedicel ca. 4 mm long; capsule green with sparse, brown, elliptical pustules, fusiform, ribbed, slightly asymmetrical both basally and apically, 12-18 × 3.0-3.5 mm, dehiscence longitudinal along a single suture.
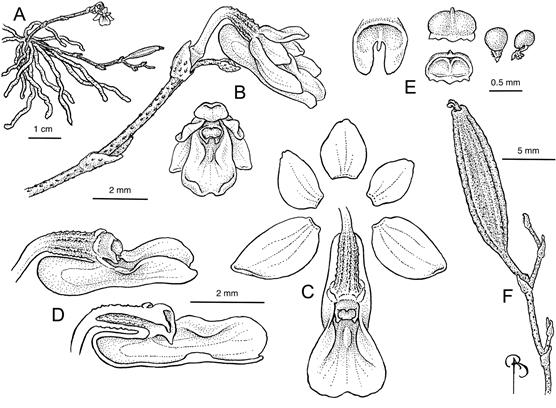
Illustrated by Bobbi Angell based on alcohol preserved specimen (later pressed and dried; E. Fernandez s.n., 28 Jun 2024; UPRRP) and Eladio Fernández photographs.
Figure 1. Dendrophylax pustulatus Ackerman & E.Fernández.

Etymology: In reference to the brown, ellipsoid pustules on the peduncle and ovary.
Distribution: Dominican Republic: province Santiago; Cordillera Central.
Taxonomic discussion
It is not entirely clear which species are most related to D. pustulatus. The most complete phylogenetic analysis published to date is of Molgo et al. (2016) and they sampled nine species, only one of which, D. porrectus, is among the short-spurred species of the Dominican Republic. DNA of D. constanzensis, D. macrocarpus (Dod) Carlsward & Whitten, D. monteverdi, and D. serpentilingua have yet to be sequenced and included in phylogenetic analyses. Nonetheless, based on morphology, we expect D. pustulatus to be related to D. serpentilingua and D. constanzensis (Table 1). This brings the number of species in the genus to 16 and increases the number of Hispaniolan endemics to six.
Ecology: Plants are epiphytic on branches of small, broadleaf trees. The population appears to be about 60 plants distributed along the southern side of Río Amina and the western side of Río Dajao. Reproductive biology is largely unknown, but flowering peak appears between June and August, and plants are likely pollinator-dependent since fruit production is sparse. Fragrance production is unknown. Pollinators of only one Dendrophylax are known, the large-flowered D. lindenii, which is pollinated by hawkmoths (Lepidoptera: Sphingidae) in Florida (USA) (Ackerman et al., 2023; Danaher et al., 2019; Houlihan et al., 2019). It is unlikely that the small flowers of the present species are pollinated by the high energy-demanding sphingids. The tiny flowers of D. pustulatus suggest that they are visited by either microlepidopterans or small bees, the latter of which are known to pollinate the small, white, short-spurred flowers of three species of Campylocentrum in Brazil (Cabral & Pansarin, 2016; Carlsward et al., 2003; Singer & Cocucci 1999).
Table 1 Species of Dendrophylax with short spurs (equal to or less than the length of ovary). HIS = Hispaniola, PR = Puerto Rico, CUB = Cuba, JAM = Jamaica, CAY = Cayman Islands, MEX = Megamexico, FLA = Florida (USA).
| Dendrophylax species | Distribution | Inflorescence | Spur | Inflorescence pubescence | Mid lobe of lip |
|---|---|---|---|---|---|
| D. constanzensis | HIS | Filiform to 7 cm long | oblong, cylindrical | setaceous-hirsute | anchor-shaped, apiculate |
| D. macrocarpa | HIS | Stout 1-2 cm long | obovoid | glabrous | ovate, acute |
| D. megarhizus | MEX | filiform to 10 cm long | scrotiform-saccate | glabrous | ovate-triangular, apiculate |
| D. monteverdi | PR, HIS, CUB, JAM | Stout < 1 cm long | oblong to obovoid | glabrous | ovate, truncate |
| D. porrectus | PR, HIS, CUB, JAM, CAY, MEX, FLA | Filiform 2-5 cm long | scrotiform-saccate | glabrous | ovate, apiculate |
| D. pustulatus | HIS | Filiform 2.2-4.5 cm long | oblong, laterally flattened | pustulate | obovate, retuse |
| D. serpentilingua | HIS | filiform 2-3 cm | obovoid, laterally flattened | hispidus | ovate, 2-tailed, apiculate |
Conservation status: We know of only one population occupying an area of approximately 2.5 hectares, with about 60 plants, all of which occur within a protected national park, Parque Nacional Armando Bermúdez. These conditions meet the IUCN Red List criterion D for endangered status (EN) (IUCN 2012).
Paratypes: República Dominicana. Cordillera Central: Provincia Santiago, La Diferencia, E. Fernández s.n., 28 Jun 2023 (UPRRP, illustration voucher). Provincia Santiago, La Diferencia, a 1 km en dirección este por el sendero, después de la caseta del Parque Nacional Armando Bermúdez, a lo largo de la vegetación contigua al Río Amina, 19°16’13”N, 71°03’07”W, elev. 735 m, 28 de marzo 2023, E. Fernandez 22B (JBSD).
Key to the species of Dendrophylax
Modified from Ackerman & Collaborators (2014) and Molgo et al. (2016)
1. Inflorescences hispid to hirsute-setaceous throughout..................................................2
2. Apical lobes of the labellum short, retrorse................D. constanzensis (Garay) Nir
2a. Apical lobes of the labellum long, filiform, pendent..D. serpentilingua (Dod) Nir
1a. Inflorescences glabrous to sparsely verrucose or pustulate........................................3
3. Labellum simple or nearly so, retuse, entire or apiculate........................................4
4. Inflorescence less than 1 cm long.......................................................................5
5. Roots smooth; fruits 8-9(-14) mm long; spur 2.5-3.0 mm long; Cuba, Jamaica, Hispaniola, Puerto Rico...D. monteverdi (Rchb.f.) Ackerman & Nir
5a. Roots verrucose; fruits 25-30 mm long; spur 3.0-3.5 mm long, Hispaniola...................................D. macrocarpa (Dod) Carlsward & Whitten
4a. Inflorescences 2-10 cm long............................................................................6
6. Spur scrotiform, saccate..............................................................................7
7. Roots 0.8-1.6 mm in diameter; lip callus 0.2-0.5 long.....................................D. porrectus (Rchb.f.) Carlsward & Whitten 7a. Roots ca. 1.4-2.3 mm in diameter; lip callus 0.07-0.20 mm long........................................................D. megarhizus Molgo & Carnevali
6a. Spur infundibuliform, tubular, or laterally flattened, not scrotiform or saccate................................................................................................................8
8. Spur 1.5-5 mm long, ovary tuberculate or pustulate.............................9
9. Spur 3-5 mm long, infundibuliform...........................D. alcoa Dod 9a. Spur 1.5-2.0 mm long, straight, laterally flattened....................................D. pustulatus Ackerman & E.Fernández
8a. Spur 15-25 mm long, ovary glabrous..............................................10
10. Roots smooth; labellum without a crest; fruits less than 15 mm long; Cuba, Jamaica, and Dominican Republic....................................................D. barrettiae Fawc. & Rendle
10a. Roots tuberculate; labellum with a simple, basal crest 1 mm long; fruits 17-20 mm long; Dominican Republic.................D. elorrhiza Dod
3a. Labellum conspicuously trilobed, deeply emarginate, or apically bilobed.........11
11. Spur less than 10 cm long...............................................................................12
12. Dorsal sepal 17-29 mm long; Jamaica......D. funalis (Sw.) Benth. ex Rolfe 12a. Dorsal sepal less than 7 mm long; Cuba and Hispaniola.......................13
13. Labellum lobes falcate, lanceolate, about 9 × 3 mm, margin entire................................................................D. gracilis (Cogn.) Garay 13a. Labellum obovate, margin minutely denticulate.................................................................D. varius (Gmel.) Urb.
11a. Spur more than 10 cm long...........................................................................14
14. Labellum apical lobes rounded, neither twisted nor elongate; Grand Cayman........................................................................................D. fawcettii Rolfe
14a. Labellum with falcate, twisting apical lobes 4.5-7.0 cm long; Cuba and Hispaniola.............................................................................................................15
15. Lateral lobes of the labellum low triangles, distally spreading 1-3 mm at the isthmus; Cuba..............................................D. lindenii (Lindl.) Benth. ex Rolfe
15a. Lateral lobes of the labellum triangular, fang-like, spreading 8-14 mm at the isthmus; Hispaniola.......................................D. sallei (Rchb.f.) Benth. ex Rolfe

