











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
Permalink
Introduction
The Anthropocene is driving many species to disappearance, as an extent that 40% of plant species are threatened by extinction (Antonelli et al. 2020). For instance, recent estimates in Orchidaceae Juss., one of the most species-rich plant families in the world, show that nearly 4342 out of 13,910 orchid species are possibly threatened by extinction (Zizka et al. 2020). Many orchid species appear to have restricted geographical distribution or low populations (Parra-Sánchez et al. in prep.), and presence of new species discovered yearly in small or remote places across the tropics shows that orchid diversity is still, to a great degree, unknown. Hence, there is an urgent need to document species before they go extinct along with their natural habitats.
Cyrtochilum Kunth, is a genus of orchids with 198 accepted species (Powo 2022). The highest diversity of species is found in the cloud forests of the Andes, and Colombia is the richest country with 85 species (Bernal et al. 2019, Ortiz Valdivieso & Uribe Vélez 2007, Uribe 2008). The geographical distribution of Cyrtochilum includes Puerto Rico and South America extending over Bolivia, Colombia, Ecuador, Peru, and Venezuela and ranging from 1500 to 3500 m of elevation, while Cyrtochilum meirax (Rchb.f.) Dalström occurs in the Caribbean (Powo 2022). The recent debate of keeping the species in a broader and more inclusive definition of Cyrtochilum (Dalström & Higgins 2020) versus splitting it into several other genera (Szlachetko et al. 2017), makes this group relevant to disentangle speciation and biogeographical processes in this family. Therefore, fieldwork provides accurate first-hand information on spatial distribution, local abundance, and morphological plasticity of species (Dalström & Higgins 2020) that ex-situ or perhaps herbaria vouchers might not supply but rather complement.
Here we propose a new species of Cyrtochilum from the province of Guavio in the east of the department of Cundinamarca, Colombia. Small populations of the new species were found dwelling epiphytically on isolated trees in a matrix of pasture.
Materials and methods
Descriptions and drawings.- The descriptions and drawings were prepared from living specimens and flowers were preserved in 70% ethanol. Flowers were dissected, measured, and photographed using Motic Digital Stereoscope SMZ-171. Vegetative structures were measured from live material and reproductive structures from spirit material. A Lankester Composite Dissection Plate (LCDP) was settled, with Adobe Photoshop 5.0 a scientific illustration was elaborated with freehand pointillism technique. The taxonomy was based on the editions of Cyrtochilum (Dalström 2001), and assisted by personal communication with Dalström. Additional graphical material was reviewed from Ortiz-Valdivieso & Uribe Vélez (2007).
Plant material.- Specimens from the herbaria: COL, JAUM, JBB, and MO (online) were consulted, and no additional material of the new species was found. Finally, material was consulted in the herbarium of the Jardin Botánico de Bogotá José Celestino Mutis (JBB), where it was included in the collection as a holotype of the species.
Taxonomic treatment
Cyrtochilum guavianum Ordóñez-Blanco & E.Parra, sp. nov. (Fig. 1 - 3)
TYPE: COLOMBIA. Cundinamarca: Mun. Gachetá. Vda. Monquentiva-Zolaque (Fig. 4-5), in cloud forest between 2549-2569 m a.s.l., 20 VI 2021, J. C. Ordóñez-Blanco 3320 (holotype: JBB).
PARATYPE: COLOMBIA. Cundinamarca: Mun. Gachetá. Vda. Monquentiva-Zolaque, in Cloud Forest at 2563 m a.s.l. 11 XI 2021. J. C. Ordóñez-Blanco 3394 (paratype: JBB).
Diagnosis: Cyrtochilum guavianum resembles C. parviflorum by its pseudobulbs with two linear to linear-lanceolate, acute leaves, fractiflex inflorescence that often branches with short, small for the genus flowers, and short, thick, straight and indistinctly arching column, and C. suarezii by the fused, concave apically carinate and above sepals, but differs from these species by its lateral sepals that are fused for half of their length to form a cymbiform, bifid synsepal, white to rose rhomboid lip without lobes, and by the short rounded simple calli on the lip, ornamented with short papillae at the base, and straight short white column.
Epiphytic, caespitose herb, medium in size for the genus, up to 26 cm high including the inflorescence. Rhizome abbreviated. Pseudobulbs heteroblastic green to brown, ovoid and slightly flattened, finely rough in appearance, with one or two apical leaves, 17.2-25.0 × 1.7-2.1 cm, surrounded basally by 4 foliaceous and deciduous, distichous sheaths. Leaves reddish-green, sub-petiolate, conduplicate, obovateligulate, obtuse apiculate, 18.5-25.0 × 2.0-2.6 cm. Inflorescence basal, erect, paniculate, indistinctly flexuous to instead fractiflex panicle with many few-flowered (5), indistinctly flexuous, slightly spaced side-branches, ca. 29-50 cm long. Peduncular bracts appressed, scale-like, ca. 2.60-3.99 mm. Floral bracts triangular, acute, 5.8 mm. Ovary green terete slightly striated, slightly curved, pedicellate, 1.4-1.8 cm long including the pedicel. Flowers stellate, yellow-brown with lip and column white to rose. Dorsal sepal shortly spathulate, elliptic, acute to minutely apiculate, entire, distinctly reflexed, 5.71-6.00 × 1.26-2.0 mm. Laterals sepals, concave, fused by half of their length to form a cymbiform, bifid synsepal, apex carinate, acute, 6.0-7.39 × 2.16-2.50 mm. Petals sub-sessile, spreading to slightly reflexed, free, obovate-elliptic, acute, somewhat concave; 4.8-5.12 × 3.28 mm. Lip white to rose, broadly elliptic to sub-rhomboid, rigidly fused to the column basally through a pair of simple, short, ventral, longitudinal calli, 3.18-5.0 mm; callus white, of two apically diverging, fleshy and blunt keels, with intermediate minute papillae near the base, and extending to ca. half the length of the lip lamina. Column white to rose, terete, straight, stout, slightly arching, ventrally shallowly canaliculated, ca. 0.36 cm long; anther cap white, globular, and campanulate, ca. 1 mm broad; pollinarium ca. 1 mm long, of two pyriform cleft, folded pollinia on a ca. 0.5 mm long almost diamond-shaped involute stipe, on a pulvinate viscidium. Seed capsule trilocular, greenish brown ca. 3.8 to 1.0 cm.
Phenology
The species has been observed in its natural habitat flowering in June and fruiting around November.
Etymology
Named after the Guavio province of Cundinamarca, north-east of Bogotá. It is Cundinamarca's largest region in area (2628 km2), constituted by eight municipalities: Gachalá, Gama, Guasca, Guatavita, Junín, La Calera, Ubalá, and Gachetá; the latter is the capital of the province and the place where C. guavianum was discovered.

A. Habit.
B. Inflorescence detail.
C. Flower, side view.
D. Flower, front view.
E. Perianth dissection whit detail of the callus.
F. Column, ventral and side view.
G. Anther cap and pollinarium.
Drawing by Omar David Bernal from the holotype.
Figure 1 Illustration of Cyrtochilum guavianum.

A. Plant habit.
B. Detail of the branch of the inflorescence.
C. Flower.
D. Perianth dissection.
E. Column.
F. Anther cap and pollinarium.
G. Capsule.
Photographs by J. C. Ordóñez-Blanco & Daniel Amaya from the holotype. LCDP by Daniel Amaya.
Figure 2 Cyrtochilum guavianum.

Photograph by Ortiz-Valdivieso, from Ortiz-Valdivieso & Uribe Vélez (2007).
Figure 3 Additional material studied of Cyrtochilum guavianum.
Habitat and ecology
Cyrtochilum guavianum is only known from one locality in the Eastern Cordillera (Fig. 4). The species was found in a pastureland close to a high Andean forest fragment. Plants grow epiphytically ca. 2-4 m above the ground on Saurauia scabra (Kunth) D.Dietr. (Actinidaceae) and Pentacalia sp. (Asteraceae).
Additional material studied
Colombia. Cyrtochilum sp. 22 in Ortiz-Valdivieso & Uribe-Vélez (2007). (Fig. 3).
Taxonomic notes
There is a debate on maintaining a broad circumscription of Cyrtochilum (Dalström & Higgins 2020) or segregating species into six smaller separated genera commonly referred to as the Cyrtochilum alliance: Buesiella C.Schweinf., Dasyglossum Königer & Schildh., Neodryas Rchb.f., Rusbyella Rolfe ex Rusby, Siederella Szlach., Mytnik, Górniak & Romowicz, and Trigonochilum Königer & Schildh (Szlachetko et al. 2017). We believe there is a need for in-depth research on the molecular signalling and morphological characters of Cyrthochilum species to understand their inherent complexity, nevertheless it will not be addressed in this paper. We have here opted for a conservative broad generic approach by keeping Cyrtochilum sensu lato for the new species described herein until further evidence shows otherwise.
Cyrtochilum guavianum is characterized by fused sepals, white to rose, broadly elliptic to sub-rhomboid lip, parallel to the column, with two simple calli ornamented with short papillae, and a straight short white column. We would traditionally refer to this species, that have fused sepals, as species in the ''Rusbyella'' group, but simple calli, sepals, and petals projections and short, thick arching column as species in the ''Parviflorum'' group. Many species in these groups appear to be synonymised by major taxonomical databases, for instance, Cyrtochilum myrianthum (Rchb.f.) Kraenzl. is considered a synonym of Dasyglossum myanthum (Lindl.) Königer & Schildh., Odontoglossum ligulatum (Ruiz & Pav.) Garay, O. myrianthum Rchb.f., while C. caespitosum (Rolfe) Dalström was synonymized with Rusbyella caespitosa Rolfe (tropicos.org; theplantlist.org). However, as we include new information, the boundaries of these dynamic groups might change. Thus, until a new updated taxonomic treatment includes the most recent molecular and taxonomic data, we would not place C. guavianum in any group (Fig. 6).
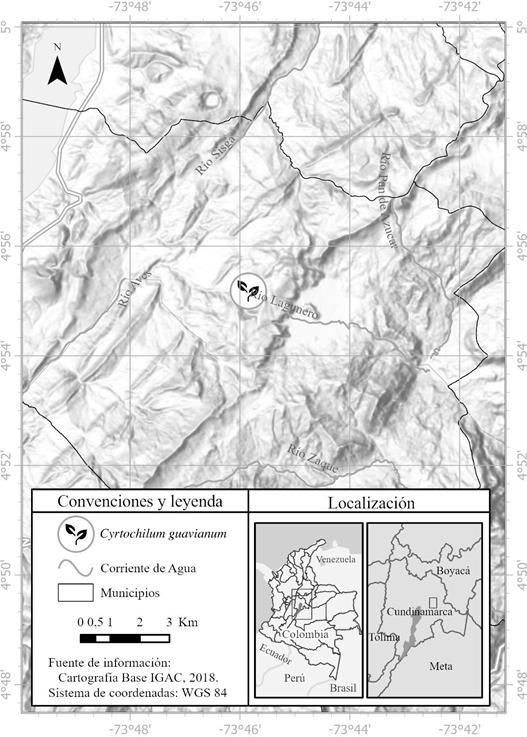
Map composed by Mónica Sandoval Ramos using the Software ArcGIS pro version 2.8.
Figure 4 Map showing the known distribution of Cyrtochilum guavianum in Colombia.

Photo by J. C. OrdóñezBlanco.
Figure 5 The disturbed habitat of Cyrtochilum guavianum in the department of Cundinamarca.

Conservation status
Cyrtochilum guavianum has only been reported in the Municipality of Gachetá in Vereda Monquentiva-Zolaque, Cundinamarca. Hence, the currently known species distribution does not allow us to make a full assessment under the International Union for Conservation of Nature red list criteria (IUCN 2020).

