English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkIntroduction
The group of species now recognized as belonging to the genus Pseudolepanthes (Luer) Archila was initially placed in Trichosalpinx Luer, under the subgenus Tubella sect. PseudolepanthesLuer (1986), then elevated to subgenus PseudolepanthesLuer (1997) to accommodate all species of Trichosalpinx with small habit and non-proliferating ramicauls with lepanthiform sheaths. Morphologically, the species in the genus have successively-flowered and progressively-elongated racemes longer than the leaves, the sepals more or less membranaceus, ciliate or spiculate with lateral sepals free or variously connate below the middle; petals entire or lobed at the base; simple lips with the disc bearing a large, verrucose callus with the broad, unguiculate base firmly attached to the base of the column, the footless column is short and suberect with the anther and stigma on the front surface, except for the arcuate column of Pseudolepanthes spathulata Luer (Luer 1997, Karremans & Vieira-Uribe 2020). Recent phylogenetic studies which included molecular data supported Pseudolepanthes as a correct genus for the species which share the characters mentioned above and also share the presence of a large, verrucose callus on the disc of the lip (Luer 1986, 1997, Bogarín et al. 2018, Bogarín et al. 2019, Karremans & Vieira- Uribe 2020). Currently, Pseudolepanthes includes 10 species distributed along the Andes of Colombia and Ecuador (Luer 1997). Here we present a new species of Pseudolepanthes discovered in a poorly explored forest protected by EcoMinga Foundation in northwest Ecuador.
Taxonomic treatment
Pseudolepanthes bihuae M.F.Monteros & Baquero, sp. nov. (Fig. 1-2).
TYPE: Ecuador. Carchi: Reserva Dracula, 2042 m, 29 January 2019, M. Monteros 203 (holotype, QCNE!).
Diagnosis: Pseudolepanthes bihuae is most similar to P. zunagensis (Luer & Hirtz) Archila, but it can be distinguished by its vinaceous or pale yellow flowers (vs. orange), an elongate, arcuate column (vs. erect, stout, shorter), the lateral sepals connate in the base, slightly free to the apex and oblique (vs. a shortly bifid synsepal), petals with sparsely tuberculate margins (vs. entire), and a lip with an elliptical blade bent down from the middle (vs. straight, elliptical-ovoid blade), with an erect, thick, widely rectangular central callus almost as wide as the blade (vs. oblong, subclavate callus much narrower than the blade).
Plant epiphytic, sympodial, caespitose herb up to 7 cm tall without inflorescence. Roots ca. 0.7 mm in diameter. Ramicauls erect, slender, stout, 3.5-4.0 cm, enclosed by 5-6 ciliate lepanthiform sheaths, 6-8 mm long. Leaf erect, coriaceous, green, suffused with purple beneath, elliptical, with two ribs parallel and close to the entire margin of the blade, the apex subacute to obtuse, emarginate, apiculate, the base cuneate into a petiole, 2.0-2.5 × 0.9-1.2 cm. Inflorescence a slightly sinuose, loose, successively-flowered raceme, up to 13-15 cm long, borne by a filiform, spiculate peduncle 0.5-1.0 cm long, from near the apex of the ramicaul; floral bracts spiculate, acuminate, 2-3 mm long; pedicels 1 mm long. Ovary sparsely spiculate, costate, 1 mm long. Sepals vinaceous or pale yellow, long- spiculate abaxially, slightly carinate to carinate along the veins on the abaxial surface. Dorsal sepal narrowly ovate, attenuate, concave, 3-veined, 8.0-9.0 × 1.5 mm. Lateral sepals narrowly ovate, attenuate, oblique, shallowly concave, connate at the base, 1-veined, 5-6 × 3 mm. Petals vinaceous to pale yellow, narrowly ovate, falcate, reflexed, acute, 5 × 1 mm, 1-veined, with the entire margins sparsely tuberculate. Lip flame- coloured, fleshy, elliptic, with an acute apex, bent down at the middle and with a thick, widely rectangular, papillose at the apex callus, the basal half with two flaps at each side lowering in height towards the base of the lip, 3.0 × 1.3-1.5 mm, the blade covered with several elongate, capitate processes at the apical half, the base smooth, shallowly concave, firmly adnate to the base of the column. Column green to cream, terete, slender, elongate, arcuate, 2 mm long, the anther apical and stigma ventral but facing forward. Anther cap yellowish, obovate, cucullate, 0.4 mm long. Pollinia 2, yellow, pyriform, attached to a detachable viscidium, 0.4 mm long. Capsule not seen.
Eponymy
Named in honor of Bihua Chen, founder of Cormorant Asset Management in Boston, USA. Bihua loves the natural world and has fond memories of the native orchids of her childhood home in China. She has given important help to the Rainforest Trust for its efforts to preserve this new species and other orchids in Fundación EcoMinga’s Dracula Reserve.
Habitat and ecology
Pseudolepanthes bihuae is, as far as we know, endemic to northwestern Ecuador, within a region that stands out for its high diversity of orchids. Only one population of this species has been found, with plants growing as epiphytes on the trunk of a tree at 2 m above the ground, in extremely humid cloud forest at 2000 m of elevation in an unexplored area within the limits of the Dracula Reserve. Other species of the subtribe Pleurothallidinae, like Brachionidium imperiale Luer & R.Escobar, Lepanthes generi Luer & Hirtz and Lepanthes gloris Luer & Hirtz, were found growing with P. bihuae. Although we compared P. bihuae with P. zunagensis, these are separated geographically; P. zunagensis was found in the east- central slopes of the Andes of Ecuador, while P. bihuae is found in the northwest Ecuadorian Andes.
Paratypes
Ecuador. Carchi: Reserva Dracula, 2042 m, 31 September 2019, M. Monteros 204 (paratype, QCNE-Spirit!). Ecuador. Carchi: Reserva Dracula, 2042 m, 31 September 2019, M. Monteros 205 (paratype, QCNE!).
Phenology
This species has always been observed blooming in its habitat throughout the year, where the humidity stays constant.
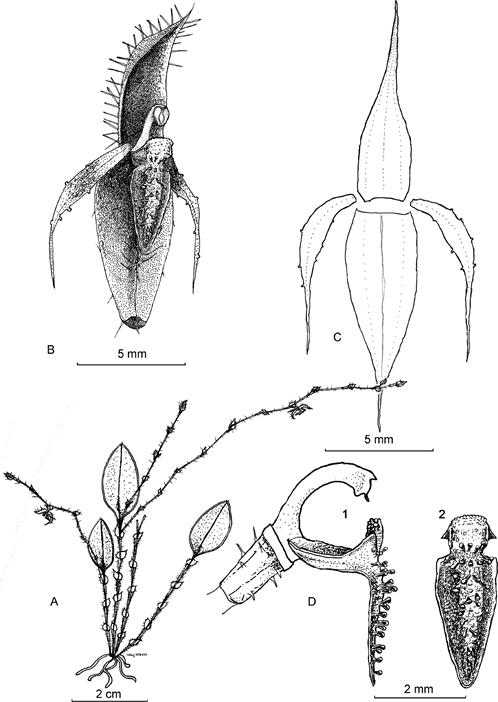
A. Habit. B. Flower. C. Dissected Perianth. D1. Column and lip, lateral view. D2. Lip, frontal view.
Drawn by Marco F. Monteros and Luis E. Baquero from the plant that served as the holotype. (MFM 203, QCNE).
Figure 1 Illustration of Pseudolepanthes bihuae M.F.Monteros & Baquero.

A. Habit. B. Flower, – view. C. Dissected perianth. D. Column and lip, lateral view. E. Column and anther, – view. F. Peduncle and floral bract.
Photographs by Marco F. Monteros from the plant that served as the holotype. (MFM 203, QCNE).
Figure 2 Lankester composite dissection plate of Pseudolepanthes bihuae M.F.Monteros & Baquero.
Pseudolepanthes bihuae is recognized by its vinaceous or pale yellow flowers, elongate, arcuate column, and also by its lip with an elliptical blade bent down from the middle with an erect, thick, widely rectangular central callus. Pseudolepanthes bihuae shares with P. zunagensis a few characteristics, such as the long spiculate, concave, ovate, acute dorsal sepal and the slender, falcate, reflexed petals, but P. bihuae is easily distinguished by the vinaceous or pale yellow flowers (vs. orange flowers in P. zunagensis), the elongate, arcuate column (vs. short, erect, stout column in P. zunagensis) and the lip with an erect, thick, widely rectangular central callus (vs. with a verrucose callus slightly clavate at the rounded apex in P. zunagensis).
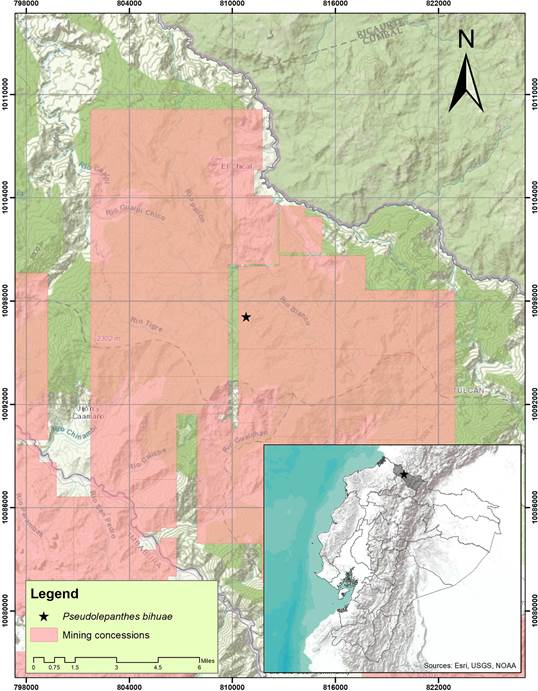
Pseudolepanthes bihuae represents the first record of the genus in northwest Ecuador. Nonetheless, most species of the Pseudolepanthes are from the western Andes of Colombia where apparently, the endemism center of this genus is located (Luer 1997). Finally, it is important to emphasize that the small population of P. bihuae in Ecuador is threatened, mainly by the illegal extraction of species and the mining projects that directly affect its distribution area in northwest Ecuador (Roy et al. 2018). For this reason, it is important and necessary to evaluate and categorize this species under the IUCN red list criteria (Fig. 3).
Conservation status
The principal threats to Pseudolepanthes bihuae are mining activity and illegal extraction of species. In Ecuador, the government owns the subsoil rights and has sold concessions in the Dracula Reserve to mining companies. Some of these mining concessions are becoming active now, and under Ecuadorian law they can work even in private protected areas such as the Dracula Reserve, where this species lives. Also, illegal mining and associated deforestation are becoming a problem in this area. We suggest considering this species as “Critically endangered” following criteria B1a, B1b(iii) IUCN (2012) since it has only been found at a single locality in Carchi province, in northwest Ecuador. If the mining activity is not controlled, the only population of P. bihuae will be strongly affected by the loss of natural forest. Therefore, this population possibly might disappear in the following years (Fig. 3).