










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista Geológica de América Central
On-line version ISSN 0256-7024Print version ISSN 0256-7024
Rev. Geol. Amér. Central n.50 San Pedro de Montes de Oca Jan./Jun. 2014
Late pleistocene mammals from El Hatillo, Panama
Mamíferos del pleistoceno terminal de la localidad de El Hatillo, Panamá
*Dirección para correspondencia:
Abstract
Late Pleistocene mammals from the El Hatillo locality on the Azuero Peninsula in Panama, originally reported by C. L. Gazin in 1957, are documented here for the first time. The following taxa are present: Hoplophorinae?, Glyptotherium floridanum, Eremotherium laurillardi, Paramylodon harlani, Equus conversidens, Platygonus sp., Odocoileus sp., Mixotoxodon larensis and Cuvieronius hyodon. This is a characteristic assemblage of late Pleistocene mammals from Central America that is dominated by fossils of Eremotherium in association with fossils of Mixotoxodon and Equus. I refer to such associations as EME assemblages and posit that most of these assemblages from Central America are of Late Pleistocene age. EME assemblages represent a mixed fauna of grazers and browsers that was common across Central America during one or more of the Late Pleistocene interstadials.
Keywords: Fossil mammals, Eremotherium, Mixotoxodon, Equus, Panamá, Pleistocene.
Resumen
Documentamos aquí por primera vez los mamíferos del Pleistoceno tardío de la localidad de El Hatillo en la península de Azuero en Panamá, originalmente reportado por C. L. Gazin en 1957. Los siguientes taxa están presente: Hoplophorinae?, Glyptotherium floridanum, Eremotherium laurillardi, Paramylodon harlani, Equus conversidens, Platygonus sp., Odocoileus sp., Mixotoxodon larensis y Cuvieronius hyodon. Este es un conjunto característico de mamíferos del Pleistoceno Tardío de Centroamérica destacado por fósiles de Eremotherium en asociación con fósiles de Mixotoxodon y Equus. Me refiero a este tipo de asociación como montaje EME y postulo que la mayoría de estos montajes de Centroamérica son de edad del Pleistoceno Tardío. Los montajes EME representan una fauna mixta de pasteadores y ramoneadores que eran muy común en toda Centroamérica durante uno o más de los fines interestadiales del Pleistoceno.
Palabras clave: Mamíferos fósiles, Eremotherium, Mixotoxodon, Equus, Panamá, Pleistoceno.
Introduction
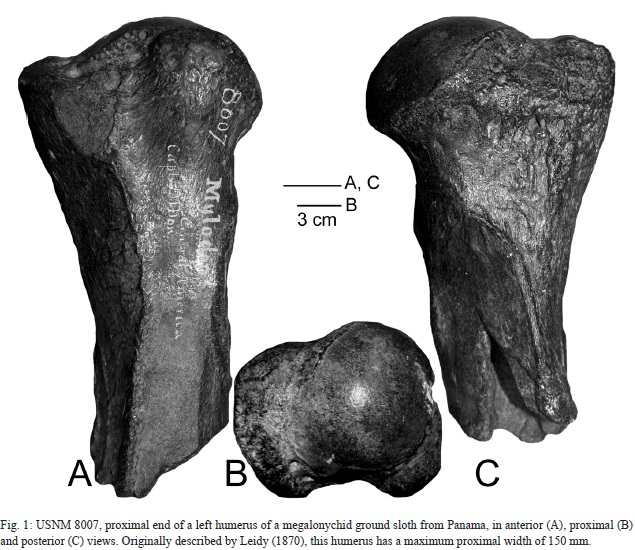
Leidy (1870) published the first report of a fossil mammal from Panama, part of the humerus of a ground sloth that he identified as Mylodon robustus (Fig. 1). This fossil, now in the collection of the Smithsonian Institution in Washington, D. C., was sent there by J. M. Dow and has no locality data other than having originated in Panama. It is the abraded proximal portion of a left humerus, but its relatively gracile morphology (note especially the small deltopectoral crest) indicates it is a megalonychid, not a mylodontid ground sloth (cf. Stock, 1925).
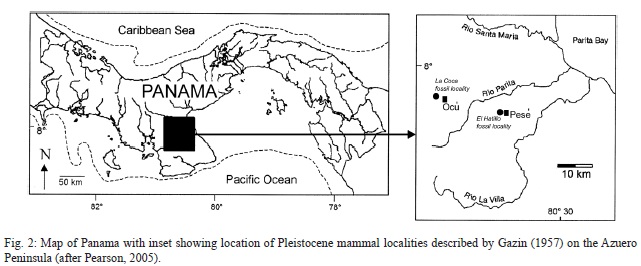
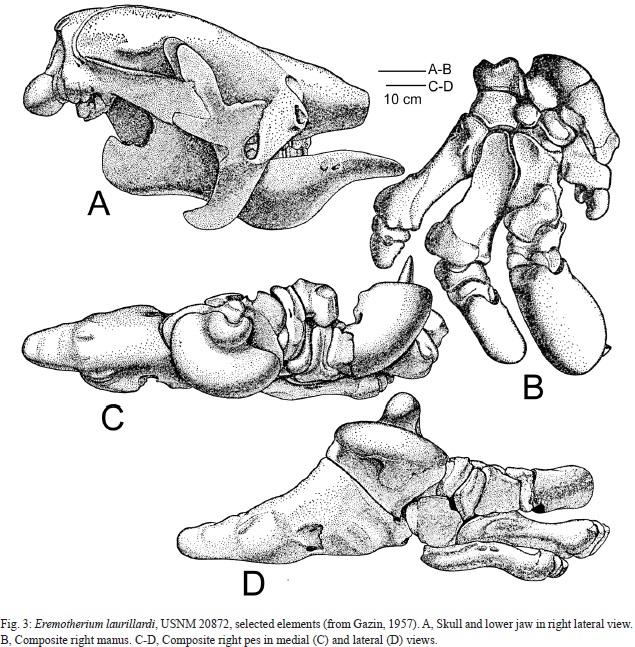
Smithsonian Institution paleontologist C. L. Gazin (1957) published the second record of fossil mammals from Panama, a diverse assemblage from Pleistocene deposits on the Azuero Peninsula (Fig. 2). However, other than that report, which did document some of the ground sloth fossils (Figs. 3-4), no further documentation of the fossils has been published. Recently, Pearson (2005) documented some other Pleistocene mammals from the Azuero Peninsula and published radiocarbon ages associated with these fossils. Miocene fossil vertebrates, including selachians, turtles, crocodylians and diverse mammals (e. g., Whitmore & Stewart, 1965; Gillette, 1984; MacFadden, 2006; Uhen et al., 2010; Cadena et al., 2012; Hastings et al., 2013; Pimiento et al., 2013; Rincon et al., 2013) have also been published from Panama.
Here, I describe and illustrate the Pleistocene fossil mammals from El Hatillo reported by Gazin (1957). They represent what I term the Eremotherium-Mixotoxodon-Equus (EME) assemblage, the most common kind of late Pleistocene mammal assemblage found at numerous localities in Central America. In this article, USNM =National Museum of Natural History, Smithsonian Institution, Washington, D.C., USA. All measurements are in mm; l = length, w = width.
Provenance
Gazin (1957) provided a detailed description of two fossil localities on the Azuero Peninsula of southern Panama: (1) La Coca, near the town of Ocú; and (2) El Hatillo near the town of Pesé (Fig. 2). He also recounted the history of their discovery and excavation.
Both localities were in surficial beds of clay associated with pond and creek deposits. Pearson (2005) found similar deposits with fossil bone at the nearby localities of La Trinidaíta and Llano Hato, from which he obtained radiocarbon ages of ~45-47,000 14C yr.B.P from charcoal associated with the bones. This apparently dates the bone de- posits to the Marine Isotope Stage 3 (MIS 3) interstadial, although I question the reliability of such relatively old radiocarbon dates. Nevertheless, given their occurrence in superficial clay deposits, preservation (relatively un-mineralized) and taxonomic composition, the vertebrate fossil assemblages from the Azuero Peninsula are certainly of Late Pleistocene age.
Systematic paleontology
Hoplophorinae?
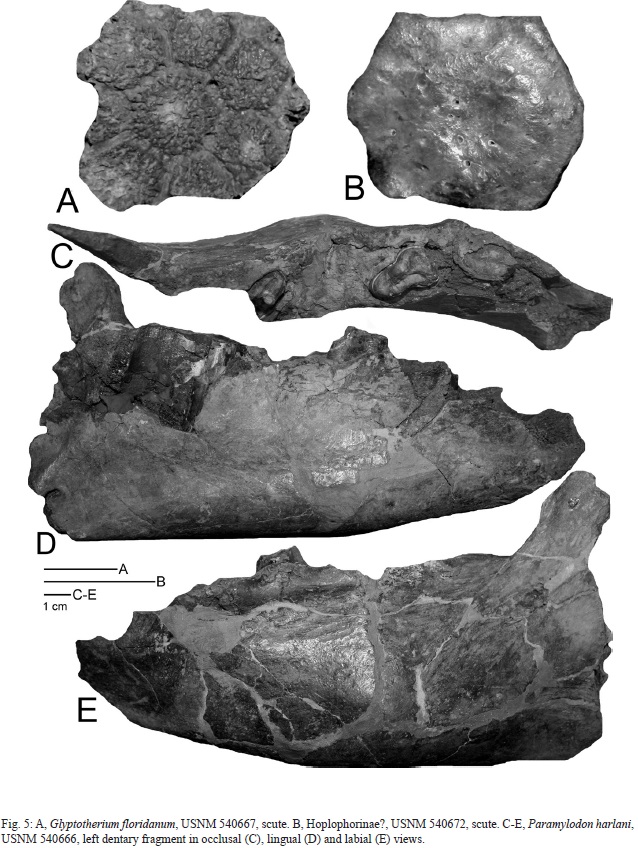
USNM 540672 (Fig. 5B) is a hexagonal scute with a maximum diameter of 24.1 mm and a max- mum thickness of 7.8 mm. It has a slightly raised central area (weak central figure) and a few small, dispersed pits (foramina) on its external surface. The internal surface is exfoliated.
Gazin (1957, p. 347) suggested that this scute might belong to a “hoplophorid?, possibly Lomaphorus sp.”. Comparison to complete ma- terial of hoplophorines (e.g., Zurita et al., 2009) supports possible assignment to the subfamily, but the scute is not sufficient morphology upon which to base a more precise identification.
Glyptotherium floridanum
A glyptodont scute (USNM 540667: Fig. 5A) has a large central figure surrounded by seven smaller figures. It has a maximum diameter of 46.5 mm and a maximum thickness of 13.8 mm.
This scute closely resembles scutes assigned to Glyptotherium floridanum from North America (Gillette & Ray, 1981, figs. 88-91), Mexico (Lucas, 2008b, figs. 10C-F) and Honduras (Jackson & Fernandez, 2005; Lucas, 2008c, fig. 3E).
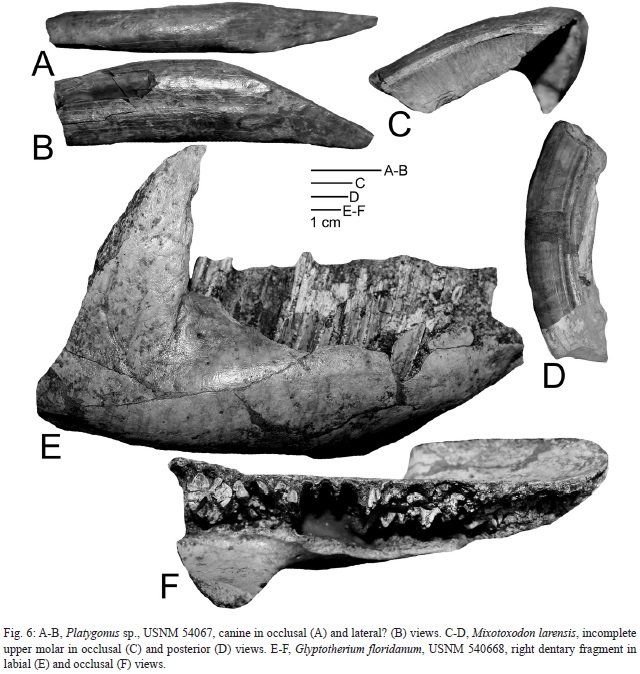
USNM 540668 (Fig. 6E-F) is the right dentary fragment of a glyptodont with very damaged teeth. The jaw closely resembles those of Glyptotherium floridanum, especially in having a flattened inferior margin of the horizontal ramus (cf. Gillette & Ray1981, figs. 16-17). A single glyptodont vertebra (USNM 540669) and other uncatalogued scute fragments from El Hatillo also likely belong to G. floridanum.
Eremotherium laurillardi
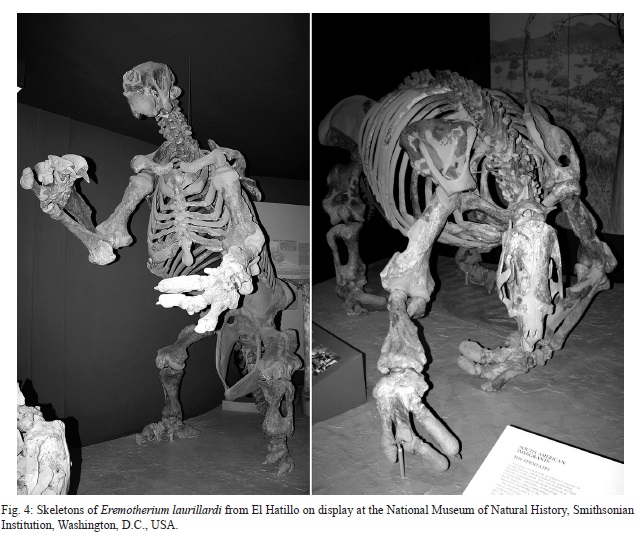
The most common fossils from El Hatillo are of the giant ground sloth Eremotherium laurillardi (Gazin, 1957; Anonymous 1958). Two skeletons from the locality are mounted and on exhibition (Fig. 4), and much more material is in the USNM collection. Gazin (1957) illustrated some of these specimens catalogued as USNM 20872 (Fig. 3). Additional specimens are catalogued as USNM 20867 (maxillary fragments) and 546680 (left dentary).
The most recent revision of Eremotherium recognizes only one polymorphic Late Pleistocene species, E. laurillardi, and identified the specimens illustrated by Gazin (1957) as E. laurillardi (Cartelle & De Iuliis, 1995). Indeed, size and some key morphological features (e.g., relatively ventral orbit and zygomatic arch, relatively shallow mandibular bulge and mandibular symphysis under m1) identify almost all of the El Hatillo sloth fossils as E. laurillardi.
Paramylodon harlani
USNM 540666 is a left dentary fragment with parts of three molariform teeth that immediately follow a nearly square (canine) alveolus (Fig. 5C-E). The first two molariform teeth are relatively short, but the last is long, consisting of multiple columns. The dentary is relatively short and deep (depth below the posteriormost molariform tooth is 85 mm) and lacks a mandibular bulge. Approximate dental measurements are: anteriormost molariform tooth l = 26.0, w = 18.0; second molariform tooth l = 18, w = 26; posteriormost molariform tooth l = 45, w = 22.
Gazin (1957) identified this fossil as cf. Glossotherium tropicum. However, in light of the work of McAfee (2009), I assign it to Paramylodon harlani. This is because USNM 540666 has a relatively straight, backward slope to its coronoid process and a relatively narrow symphyseal edge, suggesting a narrow predental spout, features that distinguish Paramylodon from Glossotherium in McAfee’s analysis. Metrics and morphology readily support assignment of the El Hatillo dentary fragment to P. harlani (cf. Stock, 1925; McDonald, 1995).
Equus conversidens
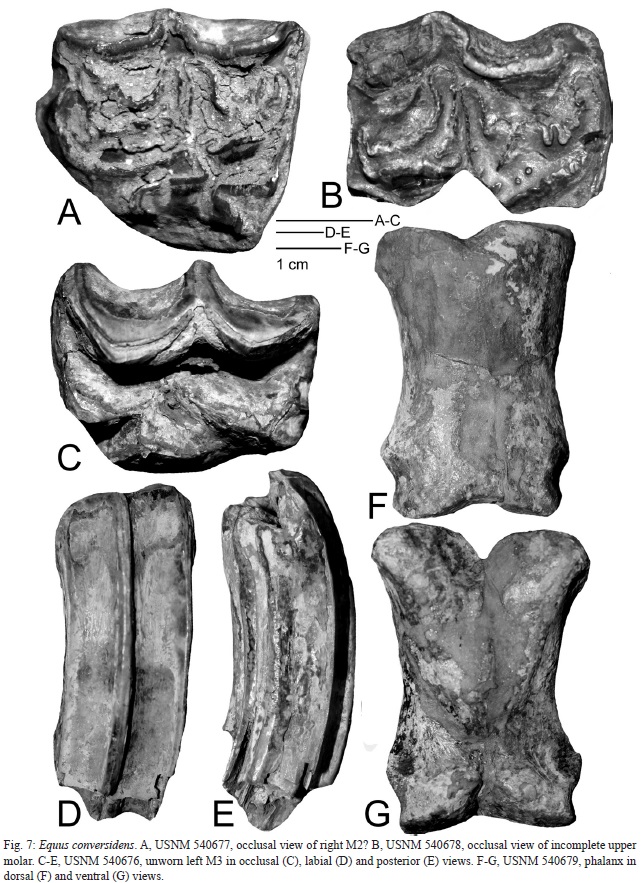
The second most abundant fossils at El Hatillo are of horses (Fig. 7). Material includes USNM 540676, an essentially unworn left M3 (Fig. 7C-E), 540677, a right M? (Fig. 7A), 540678, an incomplete upper molar (Fig. 7B) and 540679, a relatively short, broad proximal phalanx. These molars are morphologically complicated---worn crowns have relatively complex fossettes and a small pli cabillin. Measurements are: USNM 540676, l = 27.4, w = 21.5; 540677, l = 27.9, w = 23.2; 540678, l = 26.5; 540679, pha- lanx l = 75.4, proximal w = 51.5, distal w = 43.5. The proximal phalanx is short and stout, very close to the range of metrics reported for E. conversidens from New Mexico by Harris and Porter (1980, table 1). Metrically and morphologically the El Hatillo horse teeth fall within the range of variation of E. conversidens, to which they are assigned (e.g., Hibbard, 1955, fig. 3; Reynosos- Rosales & Montellano-Ballesteros, 1994).
Platygonus sp.
An isolated canine, USNM 540673 (Fig. 6A-B), belongs to a peccary. It has a crown l of 10.4 mm and a crown w of 7.2 mm. Assignment to Platygonus, the common Late Pleistocene peccary, is reasonable.
Odocoileus sp.
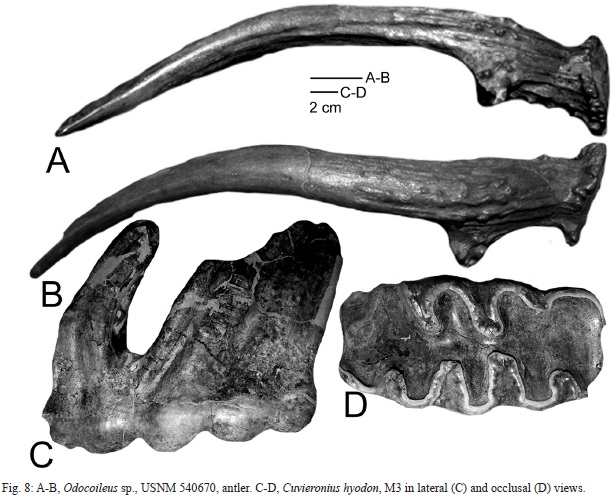
UCNM 540670 (Fig. 8A-B) is a small, dichotomously forked antler with a steeply rising beam. Total length = 167 mm, and diameter of the base = 29 mm. This antler is a good match for antlers of Odocoileus (cf. Frick, 1937), but not sufficient material upon which to base a species- level identification.
Mixotoxodon larensis
USNM 540671 (Fig. 6C-D) is an incomplete cheek tooth that has a crown length of at least 50 mm and is very hypsodont (crown height exceeds 136 mm). This tooth clearly is part of an upper molar of a toxodont. It corresponds well to material from northern South America and Central America referred to Mixotoxodon larensis (Van Frank, 1957; Laurito, 1993; Lucas et al., 1997; Lucas, 2008c), so I assign it to that taxon.
Cuvieronius hyodon
The only gomphothere fossil from El Hatillo in the USNM collection is USNM 540665, a very worn M3 (Fig. 8C-D). It has five lophs, a maximum length of 185 mm and a maximum width of 92 m. The tooth closely resembles molars of Cuvieronius hyodon from Florida, Costa Rica and El Salvador (Lucas et al., 1997; Cisneros, 2005; Lucas, 2008a). However, isolated molars of Neotropical gomphotheres are not generically diagnostic of Cuvieronius or Notiomastodon (e.g., Lucas & Alvarado, 2010; Lucas, 2013). Nevertheless, the only Pleistocene gomphothere from Central America is C. hyodon (Lucas & Alvarado, 2010), so referral of the El Hatillo molar to this species seems reasonable.
Discussion
The El Hatillo fossil mammals are a characteristic assemblage of Late Pleistocene mammal fossils from Central America. Thus, like most other Central American Late Pleistocene mammal assemblages, a microfauna, especially of rodents, was not collected at El Hatillo. Also, like other Central American late Pleistocene mammal assemblages, the El Hatillo mammals are almost all large edentates and ungulates, and are a nearly equal mixture of South American (glyptodonts, sloths, toxodonts) and North American (horses, peccaries, deer, proboscideans) immigrants. Eremotherium laurillardi dominates the El Hatillo assemblage, and also dominates many other Late Pleistocene assemblages in Honduras, El Salvador, Guatemala, Nicaragua, Costa Rica and Panama (e.g., Webb & Perrigo, 1984; Lucas et al., 1997, 2007, 2008; Cisneros, 2005; Jackson & Fernandez, 2005; Lucas, 2008c). Indeed, Eremotherium is one of the most characteristic Late Pleistocene mammal fossils in Central America.
I term such Central American assemblges EME assemblages, for the association of Eremotherium, Mixotoxodon and Equus. Furthermore, most of these assemblages appear to be of Rancholabrean age in the North American land-mammal biochronology and Lujanian in the South American biochronology. Thus, in some of these assemblages, Bison is present, providing prima facie evidence of a Rancholabrean age. The minimum age of EME assemblages in Central America presumably is end-Pleistocene, ~ 10 kya, as this is the minimum age of extinction of the megafauna in Central America (cf. Barnosky & Lindsey, 2010). The oldest age of the EME assemblages is less certain. In North America, Eremotherium has its oldest record in the Blancan of Florida, so it must have been among the early immigrants from south to north after the initiation of the great American biotic interchange. And, a few Central American fossil mammal assemblages with Eremotherium and Mixotoxodon are older than Rancholabrean (e.g., Cisneros, 2005).
Thus, the association of Eremotherium and Mixotoxodon is common in Central America, and mostly of Rancholabrean age. I conclude that most of the EME assemblages in Central America are of Late Pleistocene age, between 250 and 10 kya; most are likely younger than 100 kya. They thus represent a mixed fauna of grazers and browsers that was common across Central America during one or more of the late Pleistocene interstadials.
References
ANONYMOUS, 1958: Paleontological discovery in Panama.- Discovery, 316-317. [ Links ]
BARNOSKY, A.D. & LINDSEY, E.L., 2010: Timing of Quaternary megafaunal extinction in South America in relation to human arrival and climate change.- Quat. Internat. 217: 10-29. [ Links ]
CADENA, E., BOURQUE, J.R., RINCON, A.F., BLOCH, J.I., JARAMILLO, C.A. & MACFADDEN, B.J., 2012: New turtles (Chelonia) from the late Eocene through late Miocene of the Panama Canal basin.- J. Paleont. 86: 539-557. [ Links ]
CARTELLE, C. & DE IULIIS, G., 1995: Eremotherium laurillardi: The Panamerican late Pleistocene megatheriid ground sloth.- J. Vert. Paleont. 15: 830-841. [ Links ]
CISNEROS, J.C., 2005: New Pleistocene vertebrate fauna from El Salvador.- Rev. Brasil. Paleont. 8: 239-255. [ Links ]
FRICK, C., 1937: Horned ruminants of North America.- Bull. Amer. Mus. Nat. Hist. 69: 1-669. [ Links ]
GAZIN, C.L., 1957: Exploration for the remains of giant ground sloths in Panama.- Smithson. Rep. 1956: 341-354. [ Links ]
GILLETTE, D.D., 1984: A marine ichthyofauna from the Miocene of Panama, and the Tertiary Caribbean faunal province.- J. Vert. Paleont. 4:172-186. [ Links ]
GILLETTE, D.D. & RAY, C.E., 1981: Glyptodonts of North America.- Smithson. Contrib. Paleobiol. 40: 1-255. [ Links ]
HARRIS, A.H. & PORTER, L.S.W., 1980: Late Pleistocene horses of Dry Cave, Eddy County, New Mexico.- J. Mammal. 61: 46-65. [ Links ]
HASTINGS, A.K., BLOCH, J.I., JARAMILLO, C.A., RINCON, A.F. & MACFADDEN, B.J., 2013: Systematics and biogeography of crocodylians from the Miocene of Panama.- J. Vert. Paleont. 33: 239-263. [ Links ]
HIBBARD, C., 1955: Pleistocene vertebrates from the Upper Becerra (Becerra Superior) Formation, valley of Tequixquiac, Mexico, with notes on other Pleistocene forms.- Contrib. Mus. Paleont. Univ. Mich. 12:47-96. [ Links ]
JACKSON, D.R. & FERNANDEZ, E., 2005: A small Pleistocene megafauna from southern Honduras.- Bull. Flor. Mus. Nat. Hist.45: 261-269. [ Links ]
LAURITO, C.A., 1993: Análisis topológico y sistemático del toxodonte de los Barrantes, Provincia de Alajuela, Costa Rica.- Rev. Geol. Amér. Central, 16: 61-68. [ Links ]
LEIDY, J., 1870: [Humerus of Mylodon from Panama].- Proc. Acad. Nat. Sci. Phil. 22: 8-9. [ Links ]
LUCAS, S.G., 2008a: Cuvieronius (Mammalia, Proboscidea) from the Neogene of Florida.- New Mex. Mus. Nat. Hist. Sci. Bull. 44: 31-38. [ Links ]
LUCAS, S.G., 2008b: Late Cenozoic fossil mammals from the Chapala rift basin, Jalisco, Mexico.- New Mex. Mus. Nat. Hist. Sci. Bull. 44: 39-49. [ Links ]
LUCAS, S.G., 2008c: Pleistocene mammals from Yeroconte, Honduras.- New Mex. Mus. Nat. Hist. Sci. Bull. 44: 403-407. [ Links ]
LUCAS, S.G. & ALVARADO, G.E., 2010: Fossil Proboscidea from the upper Cenozoic of Central America: Taxonomy, evolutionary and paleobiogegraphic significance.- Rev. Geol. Amér. Central, 42: 9-42. [ Links ]
LUCAS, S.G., ALVARADO, G.E. & VEGA, E., 1997: The Pleistocene mammals of Costa Rica.- J. Vert. Paleont. 17: 413-427. [ Links ]
LUCAS, S.G., ALVARADO, G.E., GARCIA, R., ESPINOZA, E., CISNEROS, J.C. & MARTENS, U., 2007: Vertebrate paleontology. En: BUNDSCHUH, J. & ALVARADO, G.E. (eds): Central America: Geology, resources and hazards: Taylor & Francis, London, 443-451. [ Links ]
LUCAS, S.G., GARCIA, R., ESPINOZA, E., ALVARADO, G.E., HURTADO DE MENDOZA, L. & VEGA, E., 2008: The fossil mammals of Nicaragua.- New Mex. Mus. Nat. Hist. Sci. Bull. 44: 417-430. [ Links ]
MACFADDEN, B.J., 2006: North American Miocene land mammals from Panama.- J. Vert. Paleont. 26: 720-734. [ Links ]
MCAFEE, R.K., 2009: Reassessment of the cranial characters of Glossotherium and Paramylodon (Mammalia: Xenarthra: Mylodontidae).- Zool. J. Linn. Soc. 155: 885-903. [ Links ]
MCDONALD, H.G., 1995: Gravigrade xenar- thrans from the early Pleistocene Leisey Shell Pit 1A, Hillsborough County, Florida.- Bull. Flor. Mus. Nat. Hist. 37:345-373. [ Links ]
PEARSON, G.A., 2005: Late Pleistocene mega- faunal deposits of the Isthmus of Panama and their paleoenvironmental implications.- Caribb. J. Sci. 41: 1-13. [ Links ]
PIMIENTO, C., GONZALEZ-BARBA, G., HENDY, A.J.W., JARAMILLO, C., MACFADDEN, B.J., MONTES, C., SUAREZ, S.C. & SHIPPRITT, M., 2013: Early Miocene chondrichthyans from the Culebra Formation, Panama: A window into marine vertebrate faunas before closure [of] the Central American seaway.- J. South Amer. Earth. Sci. 42: 159-170. [ Links ]
REYNOSOS-ROSALES,V.H. & MONTELLANO- BALLESTEROS, M., 1994: Revisión de los équidos de la fauna Cedazo del Pleistoceno de Aguascalientes, México.- Rev. Mex. Cien. Geol. 11:87-105. [ Links ]
RINCON, A.F., BLOCH, J.I., MACFADDEN, B.J. & JARAMILLO, C.A., 2013: First Central American record of Anthracotheriidae (Mammalia, Bothriodontidae) from the early Miocene of Panama.- J. Vert. Paleont. 3: 421-433. [ Links ]
STOCK, C. 1925: Cenozoic gravigrade eden- tates of western North America with special reference to the Pleistocene Megalonychinae and Mylodontidae of Rancho la Brea.- Carneg. Inst. Wash. Pub. 331: 1-206. [ Links ]
UHEN, M.D., COATES, A.G., JARAMILLO, C.A., MONTES, C., PIMIENTO, C., RINCON, A., STRONG, N. & VELEZ- JUARBE, J., 2010: Marine mammals from the Miocene of Panama.- J. South Amer. Earth Sci. 30: 167-175. [ Links ]
VAN FRANK, R., 1957: A fossil collection from northern Venezuela 1. Toxodontidae (Mammalia, Notoungulata).- Amer. Mus. Novit. 1850: 1-38. [ Links ]
WEBB, S.D. & PERRIGO, S.C., 1984: Late Cenozoic vertebrates from Honduras and El Salvador.- J. Vert. Paleont. 4: 237-254. [ Links ]
WHITMORE, F.C., JR. & STEWART, R.H., 1965: Miocene mammals and Central American seaways.- Science, 148: 180-185. [ Links ]
ZURITA, A.E., CARLINI, A.A. & SCILLATO- YANÉ, G.J., 2009: Paleobiogeography, biostratigraphy and systematics of the Hoplophorini (Xenarthra, Glyptodontoidea, Hoplophorinae) from the Ensenadan Stage (early Pleistocene to early-middle Pleistocene).- Quat Internat. 201: 82-92. [ Links ]
BARNOSKY, A.D. & LINDSEY, E.L., 2010: Timing of Quaternary megafaunal extinction in South America in relation to human arrival and climate change.- Quat. Internat. 217: 10-29. [ Links ]
CADENA, E., BOURQUE, J.R., RINCON, A.F., BLOCH, J.I., JARAMILLO, C.A. & MACFADDEN, B.J., 2012: New turtles (Chelonia) from the late Eocene through late Miocene of the Panama Canal basin.- J. Paleont. 86: 539-557. [ Links ]
CARTELLE, C. & DE IULIIS, G., 1995: Eremotherium laurillardi: The Panamerican late Pleistocene megatheriid ground sloth.- J. Vert. Paleont. 15: 830-841. [ Links ]
CISNEROS, J.C., 2005: New Pleistocene vertebrate fauna from El Salvador.- Rev. Brasil. Paleont. 8: 239-255. [ Links ]
FRICK, C., 1937: Horned ruminants of North America.- Bull. Amer. Mus. Nat. Hist. 69: 1-669. [ Links ]
GAZIN, C.L., 1957: Exploration for the remains of giant ground sloths in Panama.- Smithson. Rep. 1956: 341-354. [ Links ]
GILLETTE, D.D., 1984: A marine ichthyofauna from the Miocene of Panama, and the Tertiary Caribbean faunal province.- J. Vert. Paleont. 4:172-186. [ Links ]
GILLETTE, D.D. & RAY, C.E., 1981: Glyptodonts of North America.- Smithson. Contrib. Paleobiol. 40: 1-255. [ Links ]
HARRIS, A.H. & PORTER, L.S.W., 1980: Late Pleistocene horses of Dry Cave, Eddy County, New Mexico.- J. Mammal. 61: 46-65. [ Links ]
HASTINGS, A.K., BLOCH, J.I., JARAMILLO, C.A., RINCON, A.F. & MACFADDEN, B.J., 2013: Systematics and biogeography of crocodylians from the Miocene of Panama.- J. Vert. Paleont. 33: 239-263. [ Links ]
HIBBARD, C., 1955: Pleistocene vertebrates from the Upper Becerra (Becerra Superior) Formation, valley of Tequixquiac, Mexico, with notes on other Pleistocene forms.- Contrib. Mus. Paleont. Univ. Mich. 12:47-96. [ Links ]
JACKSON, D.R. & FERNANDEZ, E., 2005: A small Pleistocene megafauna from southern Honduras.- Bull. Flor. Mus. Nat. Hist.45: 261-269. [ Links ]
LAURITO, C.A., 1993: Análisis topológico y sistemático del toxodonte de los Barrantes, Provincia de Alajuela, Costa Rica.- Rev. Geol. Amér. Central, 16: 61-68. [ Links ]
LEIDY, J., 1870: [Humerus of Mylodon from Panama].- Proc. Acad. Nat. Sci. Phil. 22: 8-9. [ Links ]
LUCAS, S.G., 2008a: Cuvieronius (Mammalia, Proboscidea) from the Neogene of Florida.- New Mex. Mus. Nat. Hist. Sci. Bull. 44: 31-38. [ Links ]
LUCAS, S.G., 2008b: Late Cenozoic fossil mammals from the Chapala rift basin, Jalisco, Mexico.- New Mex. Mus. Nat. Hist. Sci. Bull. 44: 39-49. [ Links ]
LUCAS, S.G., 2008c: Pleistocene mammals from Yeroconte, Honduras.- New Mex. Mus. Nat. Hist. Sci. Bull. 44: 403-407. [ Links ]
LUCAS, S.G. & ALVARADO, G.E., 2010: Fossil Proboscidea from the upper Cenozoic of Central America: Taxonomy, evolutionary and paleobiogegraphic significance.- Rev. Geol. Amér. Central, 42: 9-42. [ Links ]
LUCAS, S.G., ALVARADO, G.E. & VEGA, E., 1997: The Pleistocene mammals of Costa Rica.- J. Vert. Paleont. 17: 413-427. [ Links ]
LUCAS, S.G., ALVARADO, G.E., GARCIA, R., ESPINOZA, E., CISNEROS, J.C. & MARTENS, U., 2007: Vertebrate paleontology. En: BUNDSCHUH, J. & ALVARADO, G.E. (eds): Central America: Geology, resources and hazards: Taylor & Francis, London, 443-451. [ Links ]
LUCAS, S.G., GARCIA, R., ESPINOZA, E., ALVARADO, G.E., HURTADO DE MENDOZA, L. & VEGA, E., 2008: The fossil mammals of Nicaragua.- New Mex. Mus. Nat. Hist. Sci. Bull. 44: 417-430. [ Links ]
MACFADDEN, B.J., 2006: North American Miocene land mammals from Panama.- J. Vert. Paleont. 26: 720-734. [ Links ]
MCAFEE, R.K., 2009: Reassessment of the cranial characters of Glossotherium and Paramylodon (Mammalia: Xenarthra: Mylodontidae).- Zool. J. Linn. Soc. 155: 885-903. [ Links ]
MCDONALD, H.G., 1995: Gravigrade xenar- thrans from the early Pleistocene Leisey Shell Pit 1A, Hillsborough County, Florida.- Bull. Flor. Mus. Nat. Hist. 37:345-373. [ Links ]
PEARSON, G.A., 2005: Late Pleistocene mega- faunal deposits of the Isthmus of Panama and their paleoenvironmental implications.- Caribb. J. Sci. 41: 1-13. [ Links ]
PIMIENTO, C., GONZALEZ-BARBA, G., HENDY, A.J.W., JARAMILLO, C., MACFADDEN, B.J., MONTES, C., SUAREZ, S.C. & SHIPPRITT, M., 2013: Early Miocene chondrichthyans from the Culebra Formation, Panama: A window into marine vertebrate faunas before closure [of] the Central American seaway.- J. South Amer. Earth. Sci. 42: 159-170. [ Links ]
REYNOSOS-ROSALES,V.H. & MONTELLANO- BALLESTEROS, M., 1994: Revisión de los équidos de la fauna Cedazo del Pleistoceno de Aguascalientes, México.- Rev. Mex. Cien. Geol. 11:87-105. [ Links ]
RINCON, A.F., BLOCH, J.I., MACFADDEN, B.J. & JARAMILLO, C.A., 2013: First Central American record of Anthracotheriidae (Mammalia, Bothriodontidae) from the early Miocene of Panama.- J. Vert. Paleont. 3: 421-433. [ Links ]
STOCK, C. 1925: Cenozoic gravigrade eden- tates of western North America with special reference to the Pleistocene Megalonychinae and Mylodontidae of Rancho la Brea.- Carneg. Inst. Wash. Pub. 331: 1-206. [ Links ]
UHEN, M.D., COATES, A.G., JARAMILLO, C.A., MONTES, C., PIMIENTO, C., RINCON, A., STRONG, N. & VELEZ- JUARBE, J., 2010: Marine mammals from the Miocene of Panama.- J. South Amer. Earth Sci. 30: 167-175. [ Links ]
VAN FRANK, R., 1957: A fossil collection from northern Venezuela 1. Toxodontidae (Mammalia, Notoungulata).- Amer. Mus. Novit. 1850: 1-38. [ Links ]
WEBB, S.D. & PERRIGO, S.C., 1984: Late Cenozoic vertebrates from Honduras and El Salvador.- J. Vert. Paleont. 4: 237-254. [ Links ]
WHITMORE, F.C., JR. & STEWART, R.H., 1965: Miocene mammals and Central American seaways.- Science, 148: 180-185. [ Links ]
ZURITA, A.E., CARLINI, A.A. & SCILLATO- YANÉ, G.J., 2009: Paleobiogeography, biostratigraphy and systematics of the Hoplophorini (Xenarthra, Glyptodontoidea, Hoplophorinae) from the Ensenadan Stage (early Pleistocene to early-middle Pleistocene).- Quat Internat. 201: 82-92. [ Links ]
*Correspondencia a:
Spencer G. Lucas: New Mexico Museum of Natural History and Science, 1801 Mountain Road N.W., Albuquerque, New Mexico 87104 USA spencer.lucas@state.nm.us
1. New Mexico Museum of Natural History and Science, 1801 Mountain Road N.W., Albuquerque, New Mexico 87104 USA spencer.lucas@state.nm.us
Recibido: 4/12/2013; aceptado: 4/03/2014


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}