











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkIntroduction
In the last 40 years, coral reefs around the world have suffered great losses due to climatic extremes associated with the El Niño-Southern Oscillation. The 1982-1983 El Niño event caused coral mortalities of up to 90% in various locations in the Eastern Tropical Pacific (Glynn, Cortés, Guzmán, & Richmond, 1988; Glynn, Mones, Podestá, Colbert, & Colgan, 2017). The 1997-1998 event is recognized globally as the first massive coral bleaching event, with serious impacts on reef health. Subsequently, two other El Niño events have had global scale impacts (2010 and 2015). The latter was called El Niño-Godzilla (Schiermeier, 2015) because it developed into one of the largest recorded events (Claar, Szostek, McDevitt-Irwin, Schanze, & Baum, 2018), producing heat waves with devastating effects for marine ecosystems (Frölicher & Laufkötte, 2018). A global analysis of the impact of El Niño 2015-2016, which included more than 100 reefs, determined that 75% of these ecosystems suffered moderate to severe bleaching (Hughes et al., 2018, 2019).
In Costa Rica, El Niño 1982-1983 impacted several locations in the Pacific and the Caribbean, causing heavy coral bleaching and mortality (Cortés, Murillo, Guzmán, & Acuña, 1984). The best-documented effects were at Isla del Caño, where up to 50% of live coral cover (LCC) was lost between 1980 and 1984 (Guzmán, Cortés, Richmond, & Glynn, 1987). Isla del Coco also experienced a drastic reduction in LCC, reaching average values between 2 and 3% (Guzman& Cortés, 1992).
During the El Niño of 1991-1992, reefs along the central Pacific coast of Costa Rica experienced 57% bleaching and 9% coral mortality (Jiménez & Cortés, 2001). That same region was severely impacted during the 1997-1998 event, with a nearly 50% reduction in LCC at some sites (Jiménez& Cortés, 2003). In comparison to the 1991-1992 event, however, the 1997-1998 event resulted in higher percentages of coral bleaching (62%), but lower mortality (6%) (Jimenez et al., 2001). In the northern Pacific coast of Costa Rica, bleaching only affected 32% of the colonies, however, small populations of the coral Leptoseris papyracea experienced mortalities of up to 90%. In the South Pacific, specifically in Golfo Dulce, the percentage of bleaching was higher (70%) than in the northern coast of the country and mortality reached up to 12% of the colonies (Jiménez, Cortés, León, & Ruiz, 2001). In Isla del Caño, the mortality associated with the 1997-1998 event was 5%, which suggests a greater tolerance of these corals in comparison with other Pacific localities (Guzmán & Cortés, 2001). In contrast, the corals of Isla del Coco underwent intense bleaching, but with lower coral mortality compared to previous El Niño events; between 1987 and 2002, a recovery of LCC was observed, which increased from 3% to 23% (Guzmán& Cortés, 2007).
In the last 40 years, marine conservation strategies in Costa Rica have focused on the creation of Marine Protected Areas (MPAs) that can safeguard vulnerable ecosystems (Alvarado, Cortés, Esquivel, & Salas, 2012). Most of these MPAs include coral reefs as conservation elements (Alvarado, Herrera, Corrales, Asch, & Paaby, 2011; Alvarado et al., 2017), and the reduction of certain anthropogenic pressures (fishing, tourism, sewage) in these MPAs is thought to result in greater resilience and reproductive success for reefs (Hughes, Graham, Jackson, Mumby, & Steneck, 2010; Mumby & Steneck, 2011; Richmond, Tisthammer, & Spies, 2018). Therefore, this research seeks first to determine the impact of the most recent El Niño event (2015-2016) on the coral reefs of the Pacific coast of southern Costa Rica and Isla del Coco, as well as to quantify if the established conservation actions (MPAs) have been key in the resilience of these ecosystems.
Materials and methods
Between 2013 and 2017, a series of coral reefs health assessments in the Pacific of Costa Rica were conducted (Alvarado, Beita, Mena, Fernández-García, & Guzmán, 2015; Alvarado et al., 2016, Alvarado et al., 2018). These assessments focused solely on establishing the health status of these ecosystems, and here provides the baseline of their condition prior to the 2015-2016 global warming event. Repeated assessments at the same sampling sites were used to quantify the impact of the 2015-2016 El Niño event on benthic composition and percent cover. In Isla del Coco (5°33’N-87°02’W), two pre-event evaluations were carried out (July 2013 and February-March 2014) and one after the event (December 2016). In the southern coastal sites (9°13’N-83°50’W and 8°38’-83°17’W) the pre-El Niño assessments were carried out in February 2013 and January 2014 and one after the event in January 2017.
To describe the behavior of the sea surface temperature (SST) and the degree heating weeks (DHW), the monthly averages data were obtained, from both locations, since January 2012 to December 2017 of the Pacific Islands Ocean Observing System platform (http: //pacioos.org).
The benthic coverage of coral reef communities was estimated using underwater visual censuses via scuba. On each reef, 10 to 15 10-m long transects were arranged parallel to the coast and separated from each other by 3 m (Weinberg, 1981). A 1 m2 grid divided into 0.01 m2 cells was placed along each transect to cover the entire 10-m length. Each diver counted the number of cells occupied by different substrate categories: live coral, macroalgae, algal carpet or “turf”, and crustose coralline algae (CCA). In Isla del Coco, work was carried out at three depths (shallow 4-8 m, intermediate 9-12 m, and deep 13-16 m), while along the southern coast, two depths were surveyed (shallow 2-8 m and deep 8-16 m). The differences in the bathymetric distribution of the surveys are due to the fact that in Isla del Coco the reefs are more complex and grow deeper than the coastal reefs of the South Pacific of Costa Rica.
To determine if conservation strategies influenced the response (changes in LCC) of coral reefs to the impact of El Niño 2015-16, each site was classified into one of three management categories: 1) Marine Protected Area (MPA) (managed by the Ministry of Environment and Energy), 2) Marine Responsible Fishing Area (MRFA) (managed by the Costa Rican Institute of Fisheries and Aquaculture), and 3) unprotected site (NoMPA) (Table 1).
TABLE 1 Reef locations in the Pacific of Costa Rica where the bottom was evaluated between 2013 and 2016. MPA: Marine Protected Area; MRFA: Marine Responsible Fishing Area; NoMPA: unprotected site
| Locality | Coordinates | Management category | Year of establishment | Shortest distance to human population center |
| Isla del Coco | 5°32’45.40” N 87°3’44.30” W | MPA | 1978 | 550 km |
| Dominicalito | 9°13’31.80’’ N 83°50’47.30” W | NoMPA | - | < 1km |
| Marino Ballena | 9°6’23.80” N 83°43’34.20” W | MPA | 1992 | < 1km |
| Isla del Caño | 8°42’45.70” N 83°53’23.20” W | MPA | 1978 | 24 km |
| Península de Osa | 8°40’15.58” N 83°43’5.44” W | NoMPA | - | 10 km |
| Golfo Dulce | 8°39’25.40” N 83°16’27.80” W | MRFA | 2010 | < 1 km |
To compare the pre and post-El Niño (2015-2016) condition of the coral reefs of the Costa Rican Pacific, a Bray-Curtis similarity matrix was constructed using the average percentage cover of each substrate category, by location and year, after standardizing and transforming (log10 (x + 1)) the values. The management category of each locality (MPA, MRFA, NoMPA) and the years sampled (2013, 2014, 2016) were considered as factors. From the similarity values, a non-metric multi-dimensional scaling analysis (nMDS) and an analysis of similarities (ANOSIM) were performed. Likewise, in order to determine which substrates contributed most to the similarity between management categories and year, a similarity percentage-species contribution test (SIMPER) was performed. All analyses were performed with the software PRIMER 7 (Clarke & Gorley, 2015).
Results
For both locations, an increase in the SST is observed for the second semester 2015 and the first semester 2016, with average temperatures above 29.5°C. However, there is a slight difference between Isla del Coco and Golfo Dulce, in terms of the duration of these hot periods. It was observed that DHW, which may be evidence of coral bleaching (> 4°C-weeks), were more persistent for Golfo Dulce than for Isla del Coco (Fig. 1).
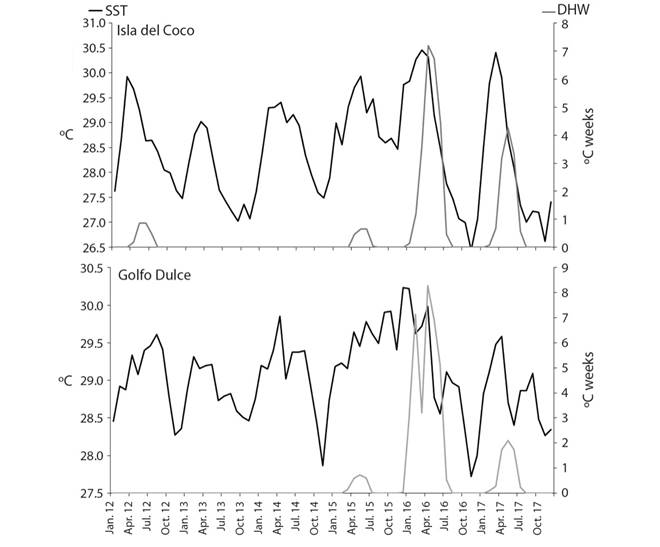
Fig. 1 Surface sea temperature (SST) (ºC) (black line) and degree heating weeks (DHW) (ºC weeks) (gray line) in Isla del Coco and Golfo Dulce form January 2012 until December 2017.
Over the study period, LCC was higher in either MPA or MRFA sites (18.17±3.86% and 24.72±8.61%, respectively), compared to NoMPA sites (7.39±3.23 %) (Fig. 2A). Within MPAs, CCA coverage (12.25±2.70%) was also higher than in NoMPAs (6.00±1.40%). In NoMPAs, the predominant substrates were macroalgae (2.56 ± 0.87%) and turf (67.51±4.89%) (Fig. 2A).
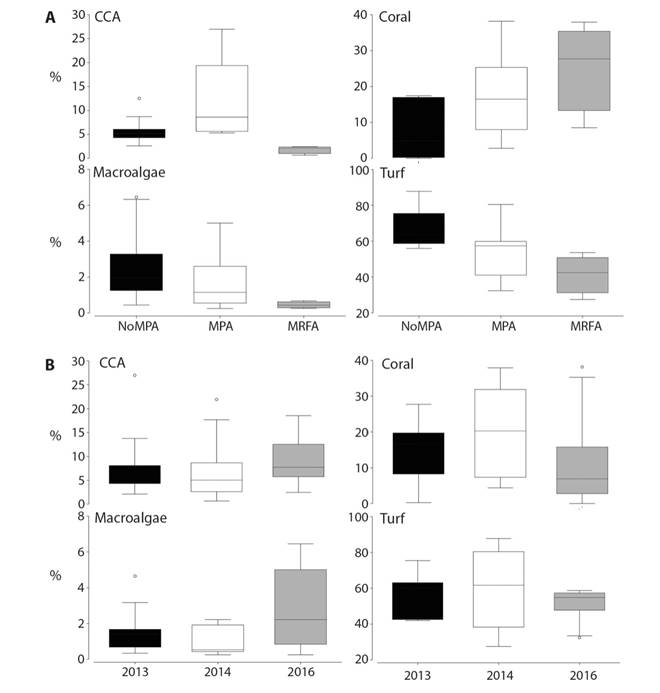
Fig. 2 Average percentage coverage (%) of crustose coralline algae (CCA), live coral, macroalgae and turf according to A) Management category (MPA: Marine Protected Area, MRFA: Marine Responsible Fishing Area, NoMPA: Unprotected site) and B) year of sampling; in the coral reefs of the Pacific of Costa Rica.
LCC averaged across all sites decreased by 25-50% between pre-El Nino (2013: 14.90±3.89% and 2014: 20.35±5.42%) and post El Niño (11.75±5.73%) surveys. The coverages of CCA, macroalgae, and turf remained stable and even showed a slight increase post El Niño (Fig. 2B).
El Niño impacts differed across the evaluated locations. For example, the benthic compositions of coral reefs of Isla del Coco and Isla del Caño were not substantially affected during the study period (Fig. 3). Reefs of Isla del Coco experienced a post-El Niño increase in LCC from an average of 28.83±6.44% in 2013/2014 to 38.16% in 2016. LCC at Isla del Caño decreased slightly from 19.71% prior El Niño to 15.79% post El Niño. In contrast, moderate to drastic losses in LCC were observed at Marino Ballena (8.26% to 2.80%) and Dominical (4.3% to 0.05%). In both locations, dissimilarities in benthic composition were also observed pre- and post El Niño (ANOSIM: R = 0.22, p <0.05) (Fig. 3). The most drastic reductions in LCC occurred on reefs of the Peninsula de Osa (from 17.00% to 5.23%), and of Golfo Dulce (from 37.90% to 8.54%).

Fig. 3 Non-metric multi-dimensional scaling analysis (nMDS) based on a Bray-Curtis similarity matrix, using the average coverage values of the benthos by location and using the following factors: 1) Management category (MPA: Marine Protected Area; MRFA: Marine Responsible Fishing Area, NoMPA: Unprotected site) and 2) year of sampling; in the coral reefs of the Pacific of Costa Rica.
These differences largely reflect the conservation strategy implemented at each locality. At the protected sites (MPA and MRFA) LCC and CCA were higher than at the unprotected sites (Table 2). Differences between sites within the same conservation category, however, reflected the contribution of CCA, as observed at Isla del Coco or Isla del Caño (Fig. 3). LCC and CCA also accounted for the largest differences in pre and post El Niño compositions, where some sites maintained similar LCC and CCA coverage over time, while others changed.
TABLE 2 Percentage Similarity Analysis (SIMPER) of the bottom coverage of the coral reefs of the Pacific coast of southern Costa Rica and Coco Island according to management category and year of sampling
| Group | Average and standard similarity | % Contribution | % Cumulative |
| Management category | |||
| Similarity | |||
| NoMPA | |||
| Turf | 46.48±11.74 | 55.90 | 55.90 |
| CCA | 18.27±5.88 | 21.98 | 77.88 |
| MPA | |||
| Turf | 37.40±10.73 | 42.58 | 42.58 |
| Coral | 23.46±4.33 | 26.71 | 69.29 |
| MRFA | |||
| Turf | 41.73±22.89 | 47.39 | 47.39 |
| Coral | 32.27±5.49 | 36.65 | 84.04 |
| Dissimilarity | |||
| NoMPA & MPA | |||
| Coral | 7.37±1.27 | 45.48 | 45.48 |
| CCA | 3.63±1.33 | 22.42 | 67.90 |
| NoMPA & MRFA | |||
| Coral | 10.16±1.43 | 48.46 | 48.46 |
| CCA | 4.56±1.47 | 21.75 | 70.21 |
| MPA & MRFA | |||
| CCA | 6.90±2.13 | 43.45 | 43.45 |
| Coral | 4.39±1.39 | 27.67 | 71.12 |
| Year | |||
| Similarity | |||
| 2014 | |||
| Turf | 41.07±9.09 | 48.73 | 48.73 |
| Coral | 24.92±4.10 | 29.57 | 78.30 |
| 2016 | |||
| Turf | 40.09±8.37 | 48.64 | 48.64 |
| CCA | 20.46±5.43 | 24.82 | 73.47 |
| Dissimilarity | |||
| 2014 & 2016 | |||
| Coral | 6.53±1.22 | 38.04 | 38.04 |
| CCA | 4.56±1.44 | 26.58 | 64.61 |
Discussion
The present study showed that the 2015-2016 El Niño had a negative impact on the coral reefs and coral communities of the Pacific of Costa Rica. Across the sites, LCC decreased by an average of 50%. Golfo Dulce reefs were the most affected by the phenomenon, with a 75% reduction in LCC. Other localities such as Isla del Caño and Isla del Coco experienced coral bleaching (C. Sánchez com. pers., G. Golfín com. pers.) but either did not experience a significant decrease in LCC or experienced a rapid recovery to LCC values similar to or higher than those prior to the El Niño. Our results also suggest that both MPA and MRFA conservation strategies have a positive effect on the condition of the reefs.
Our findings reveal that the Pacific reefs of Costa Rica have different levels of resilience to the impact of El Niño phenomenon. The differences observed between localities throughout the study period could be related to factors such as the years elapsed since the creation of the protected areas, the distance to human population centers and other sources of anthropic pressure, as well as the level of implementation of conservation strategies (Edgar et al., 2014). In turn, the difference in the impact of coral bleaching is due to the particular oceanographic conditions between the locations and the time the surface waters remain warm (Fig. 1). Coco Island is influenced by more oceanic conditions where the circulation time of surface waters is faster (Lizano, 2008). On the other hand, Golfo Dulce is a semi-closed bay, with a deep basin (about 200 m) and a shallow mouth (about 40 m), where the circulation time is slower (Svendsen et al., 2006; Morales-Ramírez, Acuña-González, Lizano, Alfaro, & Gómez, 2015). This slower circulation means that the hot waters in the Gulf are retained for a longer period, favoring bleaching. The highest mortality in the Golfo Dulce is due to the fact that the water was hot for more than four months, causing almost 100% of the colonies to bleach during that period (J. Kleypas com. pers.). In this regard, Cocos Island can function as a refuge from temperature changes that can drastically affect corals as proposed elsewhere (Riegl & Piller, 2003; McClannahan, Ateweberhan, Muhando, Maina, & Mohammed, 2007; Van Woesik et al., 2012).
The oldest conservation actions were established at the end of the 1970s in Isla del Coco and Isla del Caño. Both locations have been under a management category (National Park and Biological Reserve, respectively) for 40 years, and are categorized as Type I (highly restrictive) according to the International Union for the Conservation of Nature (IUCN) (Alvarado et al. 2012). However, the level of implementation of conservation standards differs greatly between both locations. Isla del Coco National Park is considered one of the five best MPAs in the world (Edgar et al., 2014, Strain et al., 2018), due to the adequate implementation of management actions and the high economic investment aimed at conservation. At the same time, its geographical position (located 550 km from the continental coast of Costa Rica) confers a high degree of isolation protecting these marine ecosystems from sources of anthropogenic pressure such as agricultural runoff, wastewater, sedimentation, and eutrophication. In addition, fishing in this MPA has been totally prohibited since 2001 (Alvarado et al., 2016). Finally, tourism pressure at this locality is lower than in other MPAs of the continent, with an annual average of 2700 visitors (Moreno-Díaz, 2012). In contrast, the Isla del Caño Biological Reserve is located only 24 km from the nearest population center (Drake Bay) and has a high tourist visitation all year round (about 30000 people in 2017), the main activities being scuba, snorkeling (Naranjo-Arriola, 2018) and sightseeing of cetaceans (Montero-Cordero & Lobo, 2010). Although the management actions in Isla del Caño have not been rigorously evaluated, they are likely to be low relative to the need for adequate personnel and equipment to implement them. Despite this, the geographical isolation of Caño Island does protect it to a large extent from direct impacts of normal human activities in the coastal region.
The remaining reef sites evaluated here are coastal (Table 2), they are close to sources of anthropogenic pressure, and implementation of established conservation actions are low to nonexistent (e.g. Marino Ballena and Golfo Dulce). The inadequate protection of these locations makes them more vulnerable to marine warming events since their resilience is lowered by these other stress factors (Hughes et al., 2010; Mumby & Steneck, 2011; Richmond et al., 2018). Anthropogenic pressures such as overfishing, pollution, and coastal erosion add to the natural disturbances that impact coral reefs, and this greatly reduces their capacity for recovery. The lack of execution of management actions in Marino Ballena National Park and the Golfo Dulce Responsible Fishing Marine Area, for example, is becoming a harmful factor for coral health.
The resilience of the coral reefs of Isla del Coco is a key factor for the conservation of the coral reefs of the Pacific of Costa Rica and other regions of the Eastern Tropical Pacific (ETP). A recent study determined that this island is a possible source of larval export for other ETP reefs (Romero-Torres, Treml, Acosta, & Paz-García, 2018), thus the resilience of these corals amplifies the sites’ importance in seeding other locations with bleaching-resistant corals. In addition to this, it is important to mention the key role of the changes in the clades of more resistant zooxanthellae (clade D) that have been able to occur in the reefs of Costa Rica, as has been determined for Panama and the Galapagos Islands (Baker, Starger, McClanahan, & Glynn, 2004; Cunning, Glynn, & Baker, 2013; Cunning et al., 2015). In turn, these adaptations are favored by greater reproductive success in their corals (Guzmán & Cortés, 2001, Guzmán & Cortés, 2007). In this context, understanding the zooxanthellae composition of Coco Island corals would be crucial for coral reefs resilience across the ETP region.
Although it is true that the intensity of 2015-2016 El Niño phenomenon was not as intense as was originally projected (Varotsos, Tzanis, & Sarlis, 2016), its impact on continental coastal waters was enough to reduce the LCC of the reefs in Pacific coast of southern Costa Rica except for those Isla del Coco. Although MPAs are not sufficient to protect coral reefs from future bleaching events - the Great Barrier Reef, for example, which is one of the best-managed marine parks in the world, lost 50% of its corals during bleaching events in 2016 and 2017 (Hughes et al., 2018, Hughes et al., 2019) - they do appear to be excellent tools to increase resilience to bleaching, and to enhance recovery following bleaching events. To be effective, however, MPAs need to be accompanied by efficient and effective management actions (Strain et al., 2018). They should also be complemented with other initiatives, such as effective marine spatial planning of production activities at sea, initiatives such as coral restoration that help propagate species that are more resistant to stress (Rinkevich, 2005;Rinkevich,2014; Van Oppen, Olivera, Putnamb, & Gates, 2015; Van Oppen et al., 2017), and citizen-based initiatives (Hesley, Burdeno, Drury, Schopmeyer, & Lirman, 2017) and marine environmental education projects (Chabanet et al., 2018) that support all of the above. It is the synergy of these conservation actions that ensures a promising future for current coral reefs.
Only forty years ago, we were accustomed to healthy reefs with close to 60-80% of live coral cover (Sale, 2008; Mumby, 2017). We currently live in a period of change where the average global coral coverage is less than 25%, a value that in the ‘80s was considered “mediocre” but has now become the norm (Mumby, 2017). The 2015-2016 El Niño decreased LCC of the coral reefs of the Costa Rican Pacific, but at the same time, it revealed an improved resistence of some reefs to bleaching that correlated with their degree of protection. While we cannot rule out other factors, such as past bleaching events which may have winnowed out the less resilient corals, or regional differences in the severity and/or duration of the thermal stress, this analysis provides evidence of the need to reinforce all management actions, as a primary step toward maintaining a level of reef health that can be sustained into the future.
Ethical statement: authors declare that they all agree with this publication and made significant contributions; that there is no conflict of interest of any kind; and that we followed all pertinent ethical and legal procedures and requirements. All financial sources are fully and clearly stated in the acknowledgements section. A signed document has been filed in the journal archives.

