











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkIntroduction
The signatures imposed by fragmentation on genetic diversity of tropical trees have been well summarized (Lowe, Boshier, Ward, Bacles, & Navarro, 2005; Aguilar, Ashworth, Galetto, & Aizen, 2006; Aguilar, Quesada, Ashworth, Herrerias-Diego, & Lobo, 2008). These works predict that adult genetic diversity is rarely going to be impacted by habitat loss and a reduction in diversity is more commonly observed in progenies (Ellstrand & Elam, 1993; Young, Boyle, & Brown, 1996; Lowe et al., 2005; Vranckx, Jacquemyn, Muys, & Honnay, 2012). Gene flow may ameliorate the negative effects of habitat loss and isolation by increasing effective population size and introducing novel alleles into fragments (White, Boshier, & Powell, 2002), thus reducing the detrimental effects of drift (Ellstrand & Elam, 1993; Couvet, 2002). For tropical trees, gene flow across fragmented landscapes partially depends on life history traits, pollinator and seed dispersal agents, as well as the size and distribution of forest fragments (Nason & Hamrick, 1997). Habitat loss was originally expected to reduce gene flow due to an increase in the distance between conspecifics (Ellstrand & Elam, 1993; Young et al., 1996) and because dispersal vectors (pollinators and seed dispersers) prefer to forage within forests and would avoid traveling over pastures or disturbed habitats to reach other trees (Sih & Baltus, 1987; Quesada, Stoner, Rosas-Guerrero, Palacios-Guevara, & Lobo, 2003; Quesada et al., 2004; Breed et al., 2015; Helbig-Bonitz et al., 2015). However, it has been shown that gene flow distances in tropical trees may actually increase in fragmented habitats (Dick, 2001; White et al., 2002; Hanson, Brunsfeld, Finegan, & Waits, 2008; Rosas, Quesada, Lobo, & Sork, 2011; Ismail et al., 2012).
Long distance gene flow, however, may be limited to some disperser types or the presence of fragments or isolated individuals that facilitate dispersal between patches (Fuchs & Hamrick, 2011). Fragments may also receive a higher proportion of long distance gene dispersal events from nearby continuous forest (Sork & Smouse, 2006; Bacles & Ennos, 2008). This rescue effect may offset the changes in the mating system and increase the effective population of male donors. However, long-distance gene flow needs to be recurrent and sustained over many generations, as well as originate from multiple sources to reduce the effects of drift (Slatkin, 1985; Bacles & Ennos, 2008; Breed et al., 2015).
Fragmentation and isolation may change seed and pollen dispersal distances within fragments, changing the spatial distribution of genetic diversity within them (Hanson et al., 2008; De-Lucas, González-Martínez, Vendramin, Hidalgo, & Heuertz, 2009; Wang, Compton, & Chen, 2011). Spatial genetic structure (SGS) is expected to increase in fragments due to limited seed dispersal and a reduction in the number of adults (Wang et al., 2011). This in turn may lower effective population sizes, even if pollen flow is extensive (Grivet, Robledo-Arnuncio, Smouse, & Sork, 2009). Seed movement may have a stronger effect on seedling structure because seeds carry both maternal and paternal gametes and determine where paternal genes will establish (Hamilton, 1999; Grivet et al., 2009).
Fragmentation of tropical ecosystems is likely to continue in the coming years (Achard et al., 2014; Haddad et al., 2015). To effectively manage and conserve tropical tree populations in fragmented and disturbed habitats, gene flow rates need to be estimated. Most studies estimate gene flow distances using open pollinated seeds. However using established seedlings to estimate gene flow jointly considers both pollen and seed flow rates, as well as other factors such as early selection on seedling establishment (Burczyk, DiFazio, & Adams, 2004; Sebbenn et al., 2011). Realized gene flow estimates, therefore, provide more accurate assessment of gene flow (Bacles, Lowe, & Ennos, 2006), which in turn may improve conservation and management strategies.
Symphonia globulifera L. f. (Clusiaceae) is a common tropical tree with a wide array of pollen and seed dispersers including large vertebrates such as birds, bats and even tapirs (seed disperser) (Pascarella, 1992; Gill, Fowler, & Mori, 1998; Torroba-Balmori et al., 2017). Bats have been proposed as the main disperser (Gill et al., 1998). These dispersers are capable of long distance gene flow, thus we hypothesize that S. globulifera should be resilient to the detrimental genetic effects of forest fragmentation and, as previously shown for other tropical tree species, we expect a significant proportion of gene flow events between fragments and continuous forests. However, fragmentation may limit pollen and seed dispersal within fragments, which should result in spatial genetic structure (SGS) within them. Previous work by Torroba-Balmori et al. (2017) found significant spatial genetic structure in several populations of S. globulifera in Africa and the Neotropics. They found that topographic complexity that modifies seed disperser behavior and consequently, their ability for long distance seed dispersal, impacted the strength of SGS in this species. They suggest that the weaker spatial genetic structure found in Neotropical populations could be attribute it to wide-ranging seed dispersers such as bats and large vertebrates that transported seeds efficiently and mixed seed shadows. Therefore, if large vertebrates were also the main dispersers in our site, we would expect to find a similar weak spatial genetic structure and no significant difference between continuous and fragmented habitats.
In this study, we will compare the genetic diversity of adults and seedlings in a continuous forest and a nearby forest fragment in southern Costa Rica and use parentage assignments to study realized gene flow rates across a disturbed landscape. We will also estimate spatial genetic structure within a fragment and a continuous forest, to determine if the spatial structure is affected by fragmentation at this site.
Materials and methods
Study Species: Symphonia globulifera L. f. (Clusiaceae) is an economically important tropical timber species also known for its medicinal properties (Bayma, Arruda, & Neto, 1998). It is a hermaphroditic species distributed from southern Mexico to Brazil and it is also found in tropical Africa (Bittrich & Amaral, 1996). Symphonia globulifera adults reach 10-40 m in height and are easily recognized by the adventitious roots in the lower trunk and bright yellow resin. The globular flowers are bright red, pink or white, produce copious amounts of nectar and hummingbirds are believed to be the main pollinator (Pascarella, 1992; Aldrich & Hamrick, 1998). In the wet tropics of Costa Rica, the most frequent flower visitors were stingless bees (Tetragonisca angustula) and the hummingbird Phaethornis longirostris (Sanfiorenzo, Sanfiorenzo, Castro, Waits, & Finegan, 2018), however in other neotropical forests butterflies and perching birds have been described as potential pollinators (Torroba-Balmori et al., 2017). Fruits are green berries 4-5 cm in diameter dispersed predominantly by bats, but monkeys (Riba-Hernández & Stoner, 2005), birds (Pascarella, 1992) and several mammals have also been observed (see table 1 in Torroba-Balmori et al, 2017). In Costa Rica this species is commonly found in wet or very wet tropical rain forests between 0-1 700 m a.s.l.
Study area and sampling design: We conducted our study in Golfo Dulce Forest Reserve (GDFR) in the South Pacific coast of Costa Rica (8°25’ - 8°50’ N & 83°15’ - 83°45’ W). This is a very humid tropical forest with annual precipitation between 4 000 and 6 000 mm and an average temperature of 27 ºC (Holdridge, 1967). GDFR is composed mainly of private farms whose forested areas are regulated by the Costa Rican government. The GDFR also includes Corcovado National Park. This creates a heterogeneous landscape of forest patches of different sizes surrounded by a matrix of small-scale plantations and pastures (Barrantes et al., 1999).
To understand how fragmentation affects gene flow patterns in S. globulifera, we sampled seedlings and adults in two sites. The first site is a large forest patch (~900 ha) of continuous mature forest, undisturbed for more than 50 years (Barrantes et al., 1999) and a forest fragment of ~40 ha located about 4 km south of the continuous forest and separated by grasslands and pastures currently used for cattle grazing. We will refer to these sites as Continuous Forest (CF) and Forest Fragment (FF), respectively. The FF was created in the 1950-60 when land was cleared for cattle farming. This fragment has experienced minor logging, but it is comparable to the larger CF in composition and structure with some secondary growth species (personal observation). Other forest patches of different sizes are located within GDFR but are more than 5 km away from the FF. We used established seedlings to estimate gene flow patterns and spatial genetic structure. We collected S. globulifera leaves from 10-20 seedlings (N = 455, 15-200 cm in height) located within a 10 m radius underneath adult trees. In order to have a similar number of seedlings in both sites, we sampled an area of approximately 10 ha in the CF and 5 ha in the FF for three consecutive years (2009-2011). For each seedling, we recorded its height, azimuth and distance from the nearest adult. We also collected leaves and cambium tissue from these adults for a total of 42 adults (DBH > 10 cm) at CF and 19 adults at FF (Table 1). Given the size of the CF, we only sampled within 100 m of the forest edge; this sampling scheme was replicated in the FF. Tissues were transported to the laboratory of Molecular Ecology at Escuela de Biología at Universidad de Costa Rica for DNA extraction and microsatellite analyses.
DNA extraction and marker amplification: All samples were stored at -20 °C until DNA extraction. Plant tissue was frozen and grounded in liquid nitrogen and DNeasy Plant Mini Kits were used for DNA extraction (Qiagen, Valencia, CA, USA). We genotyped individuals using five microsatellite loci (SSR) developed for S. globulifera: SgI9, SgC4 (Aldrich, Hamrick, Chavarriaga, & Kochert, 1998), Sg01, Sg06, and Sg10 (Vinson, Amaral, Sampaio, & Ciampi, 2005). PCR protocols followed the procedure suggested by the Multiplex reaction kit (Qiagen, Hilden, Germany). Thermal cycling conditions were optimized to an initial denaturation at 95 ºC for 15 min, followed by 35 cycles of 30s at 94 ºC,1 min and 30 s at 57 ºC, 1 min at 72 ºC and a final extension of 15 min at 72 ºC. Amplification products were visualized by capillary electrophoresis with LIZ 500 size standard marker in an ABI 3130 automatic sequencer (Applied Biosystems Inc., CA, USA). Allele sizes and multilocus genotypes were scored using GeneMarker v.1.91 Demo (SoftGenetics, PA, USA). One hundred samples were genotyped between three and five times to estimate genotyping error rates and the presence of null alleles.
Genetic diversity and structure: Genetic diversity was estimated separately for adults and seedlings within CF and FF, respectively, using GenAlEx 6.5 (Peakall & Smouse, 2006) and SpaGeDi v1.4 (Hardy & Vekemans, 2002). Diversity was quantified as the number of alleles (A), effective number of alleles (Ae), observed heterozygosity (Ho), expected heterozygosity (He) and inbreeding coefficient (F), averaged across loci. We also estimated the probability of exclusion using Cervus (Marshall, Slate, Kruuk, & Pemberton, 1998). With Micro Checker 2004 (Van Oosterhout, Hutchinson, Wills, & Shipley, 2004) we confirmed the absence of null alleles or allelic dropout. We estimated the genetic structure among adult and seedling populations using G st statistics calculated in GenoDive2.0 (Meirmans & Van Tienderen, 2004). Ten thousand permutations were used to assess the significant departure of G st from random expectations.
Structure 2.3.3 (Pritchard, Stephens, & Donnelly, 2000) was also used to study the genetic structure of seedlings and adults between sites. The number of clusters that best describes genetic differences among samples was estimated using the admixture model with correlated allele frequencies. Markov chains mixed well after 100 000 iterations and we discarded the first 10 000 iterations. The likelihood of different reproductive clusters (K) was determined by changing cluster numbers between K=1 and K=10 and averaging 20 repetitions for each K as suggested by Evanno, Regnaut, & Goudet (2005). The most likely number of clusters that describes the structure of adults and seedlings was determined by the ΔK method implemented in Structure Harvester (Earl & von Holdt, 2011). Replicate analyses for the most likely K were analyzed with the “greedy” algorithm in CLUMPP (Jakobsson & Rosenberg, 2007) to determine the mean probability of identity for all replicate runs. Admixture graphs were created in R (R Development Core Team, 2012).
Realized Gene flow: We used Cervus 3.0.3 (Marshall et al., 1998; Kalinowski, Taper, & Marshall, 2007) to conduct a parent-pair analysis on seedlings (N = 455) with all sampled adults (N = 61) as potential parents. S. globulifera seedlings collected underneath adult trees were considered a family. Parent-pair assignments were performed separately for each seedling family, however we always used all 61 adults as potential parents regardless of site (i.e., continuous forest and forest fragment). Adult genotypes were used to estimate allele frequencies for parentage analyses. In Cervus, we modeled hermaphrodites with the ability to self-pollinate. LOD-scores (log of the parental likelihood) and Delta (Difference between the two highest LOD-scores) values were compared with those from 100 000 simulated offspring genotypes. The proportion of sampled adults was set at 0.65 for families in CF and 0.90 for families in FF, assuming we had a larger fraction of unsampled parents in the CF. We chose a genotyping error of 0.01 since we only found conflicting genotypes in 6/516 repeats. Each offspring was assigned to a father and a mother according to the highest LOD values. When a seedling had multiple possible parent pairs, we chose the parents with the highest trio LOD-score (Trio: father-mother-seedling). When a parent could not be assigned, we considered it as a gene flow event from an unsampled adult.
Given the hermaphroditic nature of S. globulifera and that many seedlings under an adult were not related to that individual (see results), we did not assume that the closest tree was the seed donor to estimate pollen or seed flow distances, as it would likely bias our results. Instead, we tabulated parentage assignment and determined how many seedlings had both assigned parents within the same local population (PLL). We also tallied the number of seedlings with one local parent and another in a different site (PLF), which very likely represents pollen flow. Finally, we determined how many seedlings had both parents in a different population (PFF), which represent long-distance seed dispersal events. The distance between assigned parents for each seedling was estimated and considered an indirect measure of pollen flow distances. We estimated the distribution of pollen flow distances for seedlings in CF and FF.
We also performed Bayesian parentage assignments as implemented in the MasterBayes library (Hadfield, Richardson, & Burke, 2006) in the R programming language (R Development Core Team, 2012). MasterBayes’ assignments assume a distance effect based on an exponential distribution. MasterBayes also considers the possibility of unsampled sires and estimates the size of the unsampled population. For details see Hadfield et al. (2006). MasterBayes estimated the effect of distance on parentage assignment separately for each seedling population, using all 61 adults as possible pollen or seed donors. We used uninformative priors for the distance parameter (λ), genotyping errors and the number of unsampled adults. Parameters were initially estimated using 250 000 MCMC with a burnin of 50 000 runs and a thinning parameter of 100. After 250 000 iterations, the number of unsampled sires and dams failed to converge (autocorrelation r > 0.7), therefore we used genotyping errors (E1, E2) estimated from uninformative priors, which converged, and fixed those values in further simulations. We also used a distribution with mean = 100 and sd = 1, as a prior for the number of unsampled sires and dams. With informative priors, chains mixed well, and all values converged after 250 000 iterations, a burnin of 25 000 while sampling every 100th iteration (autocorrelation values r < 0.3). We used the posterior probability distribution of λ, to estimate 95 % confidence intervals (percentiles P2.5 and P97.5).
Seedling Spatial Genetic Structure (SGS): Spatial Genetic Structure (SGS) was tested for seedlings separately in both forest types (CF and FF) using SpaGeDi v1.4 (Hardy & Vekemans, 2002). Adult GPS positions were transformed to corresponding UTM coordinates and seedlings were assigned coordinates based on their relative distance (azimuth and distance) from the nearest geo-referenced adult. Spatial structure was evaluated using the kinship coefficient (F ij ) derived by Loiselle, Sork, Nason, & Graham (1995). We selected twelve distance categories; the first eight categories are separated by 2.5 m intervals, up to 20 m with the remaining categories separated by 10 m intervals. All categories had more than 100 pairs and represented more than 50 % of sampled population (SpaGeDi). For each distance category, we estimated an average kinship coefficient and its statistical significance by comparing estimates with confidence intervals from the null hypothesis of a random spatial distribution of genotypes. Confidence intervals were derived from 9 999 permutations of individuals among geographic coordinates. Standard errors for kinship coefficients per distance class were constructed from jackknife estimates provided by SpaGeDi.
To determine if our sampling design (i.e., collecting seedlings underneath adults) may have affected our SGS estimates, we used a 70 x 50 m control plot located randomly within the continuous forest (8°46’49.44’’ N & 83°22’4.80’’ W) where we mapped all S. globulifera seedlings for genetic analyses. Due to financial limitations, spatial genetic structure was analyzed on a random sample of 218 seedlings (15-200 cm in height) using the same twelve distance categories as described above. In all accounts the strength of SGS was estimated using the Sp statistic suggested by Vekemans & Hardy (2004). To compare Sp statistics between the forest fragment, the continuous forest and the control plot; CI for the Sp statistic was calculated as suggested by (Fenster, Vekemans, & Hardy, 2003) using 2SD of the b log estimate (sensu Vekemans & Hardy, 2004). We used 9 999 permutations to estimate the significance of the slope of b log .
Results
Genetic diversity and Structure: We analyzed 516 individuals (Table 1). Four of the five loci examined were polymorphic and produced reliable scores. Loci had an average of 23 alleles in adults and 29 alleles in seedlings with 100 % polymorphism and a combined exclusion probability of PE = 0.999. High levels of genetic diversity were common for adults and seedlings at both sites (Table 1). Genetic diversity estimates did not differ significantly between generations. However, a mild but significant heterozygote deficit in adults and seedlings in the forest fragment was observed (Table 1). Analysis showed a lack of structure between adults among sites (G ST = 0.008, P = 0.4) and a very small but significant structure for seedlings (G ST = 0.012, P = 0.04).
Adults were completely admixed and thus were grouped into a single cluster. Two clusters were identified as the most likely grouping for seedlings (Fig. 1). Seedlings showed low levels of admixture and most individuals were assigned to one of the two clusters (probability of assignment > 0.8). The first cluster includes seedlings predominantly from the CF. A larger fraction of the progeny in the FF was assigned to the second cluster which could be interpreted as gene flow events (Fig. 1).
TABLE 1 Genetic diversity estimates for Symphonia globulifera (Clusiaceae) adults and seedlings in continuous forest (CF) and in a forest fragment (FF), Costa Rica
| Habitat Type | Age | N | A | Ae | Ho | He | FIS |
| Continuous Forest | Adults | 42 | 19.75 (4.66) | 10.43 (2.84) | 0.86 (0.02) | 0.90 (0.06) | 0.045 (P = 0.063) |
| Seedlings | 237 | 27.00 (6.67) | 10.26 (2.25) | 0.89 (0.02) | 0.89 (0.05) | -0.003 (P = 0.621) | |
| Forest Fragment | Adults | 19 | 14.25 (2.36) | 9,64 (1.30) | 0.83 (0.07) | 0.91 (0.03) | 0.089 (P = 0.014) |
| Seedlings | 218 | 21.25 (4.09) | 8.69 (1.42) | 0.84 (0.02) | 0.88 (0.03) | 0.047 (P = 0.003) |
N: sample size, A: Average number of alleles, Ae: effective allele number, Ho: observed heterozygosity, He: expected heterozygosity, FIS: inbreeding coefficient. Standard deviations or p-values are shown in parenthesis.
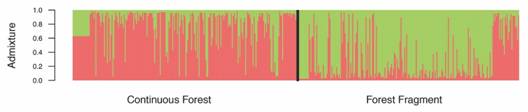
Fig 1 Population Bayesian assignment performed by Structure for Symphonia globulifera (Clusiaceae) seedlings in a continuous forest and in a forest fragment for two genetic clusters (K = 2). Each vertical bar represents an individual and is divided proportionally to the probability of assignment of each individual to a genetic cluster.
Realized gene flow: Cervus assigned at least one parent to 396 (CF = 201, FF = 195) out of 455 seedlings; the remaining 59 could not be unambiguously assigned to our adult pool and therefore were categorized as gene flow events from un-sampled parents. 51 seedlings had the same tree as a possible parent-pair, which represent selfing events. Selfing was observed in 16.9 % of seedlings in the CF (34/201) and 8.7 % of seedlings in the FF (17/195) (c2 = 3.97, d.f. = 1, P = 0.046). In our sample, more than half of the seedlings growing beneath a tree excluded that same adult as a possible parent. This was the case for 119 from a total of 201 seedlings in the CF, and in 129 out of 195 seedlings in the FF.
Parentage assignment showed that about half of seedlings in CF and FF were the result of local matings with both parents located within the same site as their progeny (PLL) (Table 2). The frequency of seedlings with one local and one external parent (PLF) was higher in the FF (41.7 % vs. 30.5 %, X2 = 5.65, d.f. = 1, P = 0.017). Outcrossed seedlings with both parents in a different site (PFF) were again more frequent in the FF compared to the CF (Table 2). Pollen flow distances follow a bimodal distribution (Fig. 2). Average (± SE) pollen flow distances were 1206.06 ± 127.5 m for the CF and 1684.46 ± 142.4 m for the FF, respectively. Within patches, mean pollen flow distances were 228.36 ± 56.6 m for the FF and 206.01 ± 59.58 for the CF.
TABLE 2 Number of outcrossed seedlings assigned by Cervus to parents in the same population as the seedling (PLL), to one parent in the same population as the seedling and a second parent in another population (PLF), and to parents that were both in a different population as the seedlings (PFF)
| PFF | PLF | PLL | Total | |
| Continuous Forest | 5 | 51 | 111 | 167 |
| Forest Fragment | 13 | 78 | 87 | 178 |
| Total | 18 | 129 | 198 | 345 |
MasterBayes produced comparable results. Confidence intervals (95 % CI) for the posterior distribution of λ showed a slight overlap between FF (λ = - 0.01957, P2.5 = -0.0246, P97.5 = -0.0148) and CF (λ = -0.0307, P2.5 = -0.042, P97.5 = -0.021), but in both cases λ < 0. Negative lambda estimates suggest that parentage assignment is more likely for adults that are closer to seedlings. The λ coefficient is greater for the FF compared to the CF, suggesting that the probability of parentage assignments declines faster in CF compared to the FF. In words, parents are located further away from seedlings in the FF. The number of unsampled sires was 167.89 (P2.5 = 142.8, P97.5 =204.9) for the CF and 70.07 (P2.5 = 35.1, P97.5 =125.8) for the FF.
Spatial Genetic Structure: We found significant SGS for seedlings at both sites (Table 3). In CF genetic relatedness between individuals deviated from random expectations for S. globulifera seedlings separated by less than 10m (Fig. 3). SGS decreased sharply after 10 meters and relatedness of seedlings separated by 12.5 m did not deviate from random expectations. Seedlings underneath a tree crown had an average kinship coefficient of F ij = 0.075 suggesting a mixture of half-sibs (HS) and unrelated individuals. For seedlings in FF, we observed a similar pattern (Fig. 3). Kinship coefficients decreased as inter-individual distances increased. However, in the forest fragment, SGS continues to be significant up to 50 m (Fig. 3). The strength of SGS was also higher in FF as suggested by the Sp statistic (Table 3). The Sp statistic in the control plot was lower but comparable to the spatial structure observed in the continuous forest (Table 3). Autocorrelograms have similar shapes between CF and FF (Fig. 2) due to similar sampling design. Although sampling in the vicinity of adults overestimates SGS, this bias should be similar for both estimates from CF and FF.
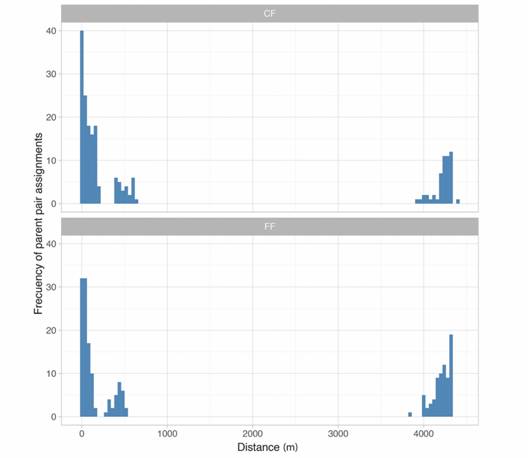
Fig. 2 Frequency distribution of distances between parent pairs (m) assigned with Cervus for seedlings of Symphonia globulifera (Clusiaceae) growing in a continuous forest (CF) and in a forest fragment (FF), Costa Rica.
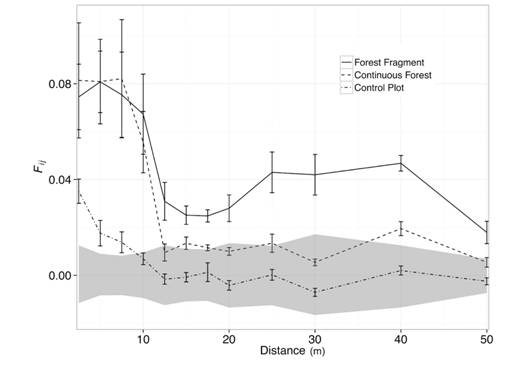
Fig. 3 Kinship estimates relative to distance between seedlings of Symphonia globulifera (Clusiaceae) in a forest fragment (solid line), a continuous forest (dashed line) and a control plot (dash-dot line). See text for details. The shaded area represents the 95 % confidence envelope about the null hypothesis of no spatial genetic structure for the forest fragment, however similar conclusions are drawn with 95 % confidence intervals for the continuous forest. F ij : kinship coefficient. Error bars depict standard errors for kinship coefficient estimates.
TABLE 3 Spatial genetic structure statistics for Symphonia globulifera (Clusiaceae) seedlings in two forest types and in a control plot in the Osa Península, Costa Rica
| Location | F(1) | b log | Sp |
| Forest Fragment | 0.0745 ± 0.0032 | -0.0223528 ± 0.004 *** | 0.02415 ± 0.008 |
| Continuous Forest | 0.0814 ± 0.0058 | -0.0175115 ± 0.001 *** | 0.01906 ± 0.002 |
| Control plot | 0.0351 ± 0.0043 | -0.0079149 ± 0.001 *** | 0.00820 ± 0.002 |
F(1): kinship coefficient for the first distance category, b log : regression slope between kinship coefficient and the log of the distance category, Sp: strength of spatial structure. SE for Sp was calculated as 2 SE of b log as suggested by Fenster et al 2003. ***P < 0.001
Discussion
Realized gene flow patterns in Symphonia globulifera, a tropical tree with mobile pollen and seed vectors, in a fragmented landscape in southern Costa Rica suggest that fragmentation may structure the seedling cohort by limiting gene flow among sites. Parentage analyses provided evidence of frequent long-distance gene flow across a fragmented landscape, which possibly occurs more commonly from the continuous forest into the forest fragment. Our results also indicate that fragmentation leads to an increase in the extent of SGS, likely a result of limited seed dispersal within fragments from a limited number of reproductive individuals.
S. globulifera has a wide geographic distribution (Dick & Heuertz, 2008) with genetically diverse populations (Aldrich et al., 1998; Degen, Bandou, & Caron, 2004; Vinson et al., 2005; Dick & Heuertz, 2008; Carneiro, Degen, Kanashiro, de Lacerda, & Sebbenn, 2009). Congruently, adults and seedlings had both high levels of genetic diversity and fragmentation appears not to have had a significant impact on genetic diversity regardless of age. Only a slight decrease in heterozygosity was observed in adults and progenies in the forest fragment, as well as an increase in inbreeding coefficients. Genetic diversity in adult tropical trees is rarely impacted by fragmentation because not enough time has passed since fragmentation for drift to act upon their diversity (Lowe et al., 2005; Julio et al., 2008; Vranckx et al., 2012). The lack of effect observed in the progeny of this study disagrees with other tropical tree studies that reported mild to strong reductions in genetic diversity of the progeny after habitat fragmentation or isolation (Lowe et al., 2005; Aguilar et al., 2008). These studies attribute the reduction in genetic diversity to a low number of pollen donors and a subsequent increase in geitonogamous matings. Although we see an increase in inbreeding coefficients in the FF, our paternity assignments, challenge the expectation of previous works that suggest that fragmentation increases selfing (Cascante, Quesada, Lobo, & Fuchs, 2001; Fuchs, Lobo, & Quesada, 2003). Selfing was more common in the continuous forest. Selfing estimates derived from parentage analyses were comparable to those found by Aldrich & Hamrick (1998) in a population of S. globulifera in Costa Rica, but higher than those reported by Carneiro in Brazil (Carneiro et al., 2009, 2011). Hummingbirds are effective outcross pollen vectors and may fly across fragmented landscapes if they are close to continuous forests (Schulke & Waser, 2001) as in our case. Another plausible explanation is that selfed seeds have lower fitness or express inbreeding depression, as expected for a predominantly outcrossing species and thus have lower germination rate (Charlesworth & Charlesworth, 1999; Wang, Hill, Charlesworth, & Charlesworth, 1999). If germination and establishment is lower in selfed progeny, by sampling plants that were between 15 cm and 200 cm in height we would be unable to accurately detect selfing events that were eliminated by inbreeding depression (Konuma, Tsumura, Lee, Lee, & Okuda, 2000; Hufford & Hamrick, 2003; Roberts, Ottewell, Whelan, & Ayre, 2014). By analyzing established seedlings, any change in the mating system caused by fragmentation may not transfer into the seedling cohorts, as selfed seeds may be less fit and fail to germinate. This is an important result suggesting that fragmentation studies that use seeds collected from maternal trees (Cascante et al. 2001, Fuchs et al., 2003) may overestimate the effects of fragmentation on selfing rates.
As expected, our parentage analyses showed that near neighbor matings were the norm in both CF and FF ( Stacy et al., 1996; Carneiro et al., 2009; Segura & Jimenez Vargas, 2014). Optimal foraging theory predicts that even pollinators that are able to travel long distances should prefer to visit nearby trees (but see Pyke, 1984; Harder & Barrett, 1996; Bezemer, Krauss, Phillips, Roberts, & Hopper, 2016). If near neighbor mating continues to be the norm in this species, and the number of possible mates is kept low due to habitat restriction or deterioration, this may eventually lead to an increase in biparental inbreeding and structure in these populations (Hamrick & Loveless, 1987). Structure within fragments may be an early sign that if isolation persists and densities decrease, genetic diversity will decline, compromising the long term viability of these populations (Newman & Pilson, 1997; Reed & Frankham, 2003; Allendorf, Luikart, & Aitken, 2012). Structure within fragments may also be a result of declines in the populations of pollinators and seed dispersers (Neuschulz, Mueller, Schleuning, & Böhning-Gaese, 2016). Lower population sizes of gene vectors should increase the detrimental effects of fragmentation by limiting gene flow, lowering genetic diversity and increasing within population genetic structure.
Despite the significant genetic structure found in seedlings, our parent-pair assignment for S. globulifera showed that dispersers effectively transfer pollen and seeds within and between the CF and FF. Within each site, more than half of the progeny underneath the canopy of a tree had another adult as its mother. This was previously observed in other tropical tree species with vertebrate seed dispersers capable of long distance travel (Hardesty, Hubbell, & Bermingham, 2006; Sezen, Wang, Sork, Leong, & Smith, 2007; Chazdon, & Holsinger, 2009). Seed dispersers are probably collecting seeds on one tree and dropping them under another conspecific in their feeding routes. This behavior should increase gene flow distances and mix progenies from multiple adults reducing the spatial genetic structure of future cohorts. Mixing progeny from different adults may also aid seedling recruitment by avoiding pathogen attacks (Hardesty, Dick, Kremer, Hubbell, & Bermingham, 2005) and density-dependent regulation (Marden et al., 2017).
A significant portion of seedlings both in the CF and in the FF had both parents on a different population located 4 km away, which represents long distance seed dispersal across a disturbed landscape. If PLF and PFF are respectively interpreted as indirect estimates of long-distance pollen and seed dispersal frequencies, long-distance gene flow was more likely to occur into the FF.Our results are congruent with previous observations on tropical trees, which show that fragmentation and habitat loss do not prevent long distance gene flow into fragments or isolated individuals, in fact, it may increase the fraction of these long distance gene flow events (Dick, 2001; White et al., 2002; Bacles et al., 2006; Rosas et al., 2011). Seed dispersal represents diploid gene flow and thus has a greater effect on increasing effective population sizes on fragments (Bacles et al., 2006). If seed dispersal is maintained between a continuous forests and a forest fragments across disturbed landscapes, it may reduce the negative effects of drift on fragments.
Maximum likelihood and Bayesian parent-pair assignments suggested that a larger fraction of pollen and seed flow events originated in the CF and were directed into the FF. These results propose that in our study system, gene flow patterns are asymmetric. Similar gene flow patterns into smaller fragments or isolated individuals have been found for other tree species (Hanson et al., 2008; Lander, Boshier, & Harris, 2010; Melo & Franceschinelli, 2016). Previous work in S. globulifera showed that reproduction by a few dominant individuals might result in asymmetric gene flow from pastures into the continuous forests (Aldrich & Hamrick, 1998). We did not find evidence of dominant adults as the same number of trees sired progeny in both the CF as in the FF (data not shown). More gene flow into fragments may be due to pollen or seed disperser behavior, foraging routes (Schulke & Waser, 2001) or because forest fragments and even isolated individuals may be attractive destinations when travelling across a disturbed landscape deprived of forest cover. Ripperger, Kalko, Rodríguez-Herrera, Mayer, & Tschapka (2015) showed that Dermanura watsoni, a possible seed disperser of S. globulifera, increased foraging distances with increased perturbation and foraged commonly from larger continuous habitats into smaller degraded forest across disturbed matrices. This foraging behavior may help explain the higher proportion of gene flow events into the FF.
Given that we did not sample adults in other fragments, and the large number of estimated unsampled sires, gene flow into fragments may also originate from these sites. Symphonia globulifera is a relatively common species in the area and misidentification due to a limited number of makers is a possibility. However, even with this caveat, our results show that fragments may not be completely disconnected from other populations, at least at the geographic scale of this study. Gene flow into the FF may have a rescue effect (Dick, 2001) by increasing both male and female effective population sizes. This rescue effect may depend on the presence and the proximity of mature forest patches and highlights the importance of keeping large forests as sources of genetic diversity, particularly if fragments act as sinks (Pulliam, 1988). Large forest patches may also house large populations of pollinators and seed dispersers, which may be necessary for long distance dispersal.
We consistently found significant SGS at CF, FF and our control plot. Spatial structure, however, was lower than expected for an animal dispersed tropical tree (Sp Œ 0.01…0.03), but comparable to those found for other neotropical populations of S. globulifera (Torroba-Balmori et al., 2017). If we compare SGS estimates from FF and CF, our results suggest that seed dispersal dynamics may change within the FF. Spatial genetic structure in S. globulifera seedlings was stronger and extended further in the FF relative to the CF. SGS is caused by a combination of limited pollen and seed dispersal, however, localized seed dispersal is the most likely cause (Epperson, 1992; Hamrick, Murawski, & Nason, 1993; Torroba-Balmori et al., 2017). Previous evidence corroborates that habitat fragmentation increases SGS in tropical trees (De-Lucas et al., 2009; Wang et al., 2011; Browne, Ottewell, & Karubian, 2015; Leclerc, Vimal, Troispoux, Périgon, & Scotti, 2015). Higher SGS in fragments is probably caused by either a change in adult tree density or changes in the abundance and behavior of seed dispersers. For S. globulifera, Degen et al. (2004) and Carneiro et al. (2009) suggested that differences in tree density across populations may shape SGS. Topographic complexity, which may influence seed disperser behavior, and adaptation to microenvironments in a heterogeneous habitat have also been proposed as possible causes of SGS in S. globulifera. However given the close proximity of the CF and the FF, these factors are unlikely to increase SGS in the FF. The spatial autocorrelogram in the FF is consistent with recruitment from a few individuals at lower densities (Fig. 2a in Hamrick & Trapnell, 2011). Therefore, we conclude that fewer seed donors may be responsible for recruitment in the FF, which increases the magnitude and length of SGS (Pardini & Hamrick, 2008). A reduction in the abundance of seed disperser in the FF could also increase the magnitude of SGS compared to the CF, however, we currently do not have information on pollinator or disperser abundances to support this.
SGS in the seedling stage generally decreases in later cohorts due to thinning (Chung, Epperson, & Chung, 2003; Fuchs & Hamrick, 2010). However, related individuals may eventually recruit into adult stages. Near neighbor mating and a lower number of possible mates in fragments could lead to an increase in biparental inbreeding and further reductions in genetic diversity within fragments (Hamrick & Loveless, 1987). This increase in the sub-structuring of genetic diversity within fragments may be an early sign that if fragmentation persists or increases, genetic diversity may continue to decline, compromising the long term viability of populations within fragments (Newman & Pilson, 1997; Reed & Frankham, 2003; Allendorf et al., 2012). An increase in the SGS of fragmented populations and significant genetic structure among seedling populations may all be telltale signs that fragmentation has had an impact on the long-term ability of fragments to sustain preexistent levels of genetic diversity.
We would like to highlight that our results are based on one fragment and one continuous forest, which could limit our generalizations. However, since our conclusions are comparable to the growing corpus of information on gene flow patterns in tropical forest fragments and parallels previous results in S. globulifera, one of the most studied tropical tree species (Aldrich & Hamrick, 1998; Carneiro et al., 2011, 2009; Torroba-Balmori et al, 2017); we are confident that our conclusions contribute valuable information on gene flow dynamics in fragmented tropical landscapes. We suggest that further studies should analyze pollen and seed flow patterns for mobile gene vectors in fragments of different sizes and pollinator and seed disperser densities.
Ethical statement: authors declare that they all agree with this publication and made significant contributions; that there is no conflict of interest of any kind; and that we followed all pertinent ethical and legal procedures and requirements. All financial sources are fully and clearly stated in the acknowledgements section. A signed document has been filed in the journal archives.

