











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkLa estimación de los contenidos de carbono en los bosques tropicales es de gran relevancia para entender su papel en el ciclo global de este elemento. Algunas investigaciones han demostrado que los bosques de la franja pantropical actúan como sumideros importantes, en especial aquellos ecosistemas localizados en tierras bajas (Clark 2007; Lewis et al., 2013; Langner, Achard, & Grassi, 2014), donde la diversidad y la densidad de madera de las especies, es uno de los atributos que más influye en la distribución de la biomasa aérea y el carbono (Malhi et al., 2006). Por otra parte, otros estudios han demostrado que los bosques tropicales de montaña, localizados a altitudes mayores a 1 500 msnm (Slik et al., 2010; Spracklen & Righelato, 2013), también almacenan grandes cantidades de carbono en su biomasa aérea, quizás como resultado de los bajos costos de la respiración, que se ven reflejados en un crecimiento óptimo de las especies (Larjavaara & MullerLandau, 2013). Las anteriores evidencias hacen suponer que el aumento en la concentración atmosférica de dióxido de carbono (CO2) puede ser mitigada a través de la conservación de los bosques maduros existentes, y/o el manejo de los bosques sucesionales (Kindermann et al., 2008). Por ello, conocer con precisión las existencias de carbono en estos ecosistemas y su dinámica en el tiempo, es crucial para desarrollar e implementar medidas eficientes y sostenibles a largo plazo, que permitan reducir las emisiones de Gases de Efecto Invernadero a la atmósfera Gases de Efecto Invernadero (GEI).
Se estima que los bosques tropicales contienen hasta 80 % del total del carbono almacenado en toda la vegetación terrestre (Phillips & Gentry 1994; Clark 2007). Particularmente, la biomasa aérea de estos ecosistemas representa 60 % o más del total de las reservas de carbono en la vegetación (Malhi et al., 2006; Saatchi et al., 2011). No obstante, datos sobre almacenamiento de carbono para otros compartimientos como necromasa, biomasa subterránea (raíces) y suelos, son más escasos (Houghton, Lawrence, Hackler, & Brown, 2001), lo cual limita sustancialmente una estimación detallada de las cantidades totales de carbono en estos ecosistemas (Sierra et al., 2007). Teniendo en cuenta que la distribución de las reservas de carbono no es homogénea en los bosques porque existe una marcada variación a nivel de tipos de bosques y paisajes (Asner et al., 2010), los estudios que aborden estas temáticas son útiles para dimensionar el aporte de los mismos en enfoques como la Reducción de Emisiones por Deforestación y Degradación (REDD+) que busca contribuir a la mitigación del cambio.
En Colombia, de acuerdo con el reporte nacional (IDEAM 2014), la tasa anual de deforestación en el país fue de 120 933 ha/año para el período 2012 - 2013. De ésta, el 57 % se presentó en la región Amazónica y el 22 % en la región Andina. La principal causa de deforestación en bosques andinos o de montaña, es la expansión de la frontera agrícola (González et al., 2011), que particularmente en el Este de los Andes, ha causado la fragmentación de los bosques de roble (Quercus humboldtii Bonplandy Colombobalanus excelsa Lozano, Hern. Cam, and Henao, J. E. Nixon and Crepet) (Armenteras et al., 2003). Los bosques tropicales de montaña son reconocidos porque han sido refugio para muchas especies en el pasado, son fuentes importantes de agua y presentan alto endemismo de especies (Van Der, Shoo, & Williams, 2009). A pesar de ello, y aunque en el país se han realizado estudios sobre estructura y ecología de este tipo de bosques (González, Coca, & Cantillo, 2007; León, Vélez, & Yepes, 2009; Parra, Díez, & Moreno, 2011), aún se desconoce cuál es el verdadero potencial de los mismos como reservorios de carbono. Los bosques del área de estudio son considerados bosques de montaña, siguiendo el concepto de cloud forest immsersion zone propuesto por Girardin et al., (2013), bajo el cual estos ecosistemas, inician a partir de 1 500 msnm, y presentan una notoria dominancia de especies de la familia Fagaceae como C. excelsa y Q. humboldtii, típicas de bosques propios de la formación andina descrita por Cuatrecasas (1958). Las Fagáceas en general, se destacan por presentar un patrón de distribución único, son una familia de latifoliadas dominante en zonas templadas de los bosques de Eurasia y América del Norte, y son también muy importantes en bosques montanos tropicales de hoja perenne del sudeste de Asia, y en Suramérica (González et al., 2007; León et al., 2009; Parra et al., 2011; Culmsee, Leuschner, Moser, & Pitopang, 2010).
En este estudio, se caracterizó la estructura de los bosques tropicales de montaña localizados en el sur del departamento del Huila (Colombia) y dominados por la especie C. excelsa, y se realizó una estimación detallada de la biomasa y los contenidos de carbono, con el objetivo de identificar su papel en el almacenamiento de carbono, evaluar la contribución de los compartimientos analizados (e.g. biomasa aérea, biomasa subterránea y necromasa), y realizar aportes para el enfoque REDD+ a escala de proyectos a partir de los resultados obtenidos.
Materiales y métodos
Área de estudio: El estudio fue realizado en el sur del departamento de Huila (Colombia), en áreas de bosque natural pertenecientes al Parque Natural Regional Corredor Biológico Parque Nacional Natural Puracé - Parque Nacional Natural Cueva de los Guácharos, entre las coordenadas 1°34'55,19" N - 76°0'24,35" W y 2°3'55,97" N - 76°33'33,45" W. El Parque tiene un área total de 73 183 ha, de las cuales 69 443 ha corresponden a áreas de conservación, y 3 740 ha a zonas de producción sostenible, y abarca los municipios de San Agustín, Palestina, Pitalito y Acevedo (CAM, 2009). Esta zona forma parte del Macizo Colombiano, el cual fue declarado como "Reserva de la biosfera" por la UNESCO en el año 1979, dado sus altos índices de diversidad biológica, la oferta hídrica de importancia regional y nacional, su función de conexión entre diversos ecosistemas de montaña (e.g. páramo, bosques altoandinos y subandinos), y la gran diversidad cultural en la región (CAM, 2009).
De acuerdo con las estaciones meteorológicas del Instituto de Hidrología, Meteorología y Estudios Ambientales de Colombia - IDEAM: Bajo Frutal (San Agustín), El Tabor (Palestina) y San Adolfo (Acevedo), a lo largo del corredor, se presenta un régimen de precipitación bimodal tetra-estacional, con valores promedio anuales de 1 201.9 mm en la estación de Suaza, hasta 1 402.3 mm en la estación de San Agustín (IDEAM, 2005). El pico de mayor precipitación se encuentra en el mes de mayo para las estaciones de Pitalito y Suaza, y en junio, en la estación de San Agustín. En las estaciones San Agustín y Acevedo, los valores medios de temperatura oscilan entre 11.1 ºC y 22.4 ºC, respectivamente (González et al., 2007). Se presenta un régimen isotérmico con variaciones promedio de 1.5ºC a lo largo del año. Los suelos del área de estudio poseen texturas desde francas hasta areno-francosas, pH promedios de 3.8 a 5.3, muy bajos contenidos de fósforo (0 - 3.79 ppm), contenidos de materia orgánica entre 1.5 % y 5.3 % y capacidad de intercambio catiónico (CIC) entre 22.6 y 37.2 meq/100 g (González et al., 2007). Las zonas de vida presentes en el área de estudio siguiendo el sistema de clasificación de zonas de vida de Holdridge (1967), son bosques húmedos y muy húmedos premontanos, y bosques húmedos y muy húmedos montanos bajo. Éstas fueron identificadas empleando el Atlas Climatológico de Colombia (IDEAM 2005).
Establecimiento de parcelas: Durante el 2010 y 2011 se establecieron 44 parcelas (50 x 50 m; 0.25 ha) en bosques maduros de la zona, para un área total inventariada de 11 ha. Veintiséis (26) de ellas fueron establecidas en 2010 por el Instituto de Hidrología, Meteorología y Estudios Ambientales (IDEAM) con el fin de validar el protocolo nacional para la estimación de biomasa y carbono. Posteriormente, en 2011, ONF Andina estableció dieciocho (18) parcelas adicionales para cumplir con los requisitos de la metodología VM0015 del estándar voluntario de carbono Verified Carbon Standard (VCS), en relación con los errores asociados a la estimación de carbono en proyectos tipo REDD+ (Pedroni, 2012). En ambos casos, los puntos fueron ubicados inicialmente de forma aleatoria sobre un mapa, y durante la fase campo fueron localizados en el terreno empleando GPS. Las parcelas fueron georreferenciadas tomando como punto base el vértice de inicio, y se empleó el sistema de coordenadas geográficas WGS84.
Se marcó y midió el diámetro (D, medido a 1.30 m de altura sobre la base del suelo para árboles sin irregularidades) de todos árboles, palmas y helechos con D ≥ 15 cm dentro de cada parcela; los árboles muertos en pie fueron incluidos. Los individuos fueron medidos a partir de 15 cm de diámetro, siguiendo los requerimientos de la metodología VM0015 del VCS (Pedroni, 2012), lo cual además era costo-efectivo. Para árboles con irregularidades a los 1.30 m de altura (e.g. bambas o raíces tabulares), el D fue medido por encima de las mismas. La medición de los árboles con irregularidades se realizó siguiendo el protocolo reportado por MacDicken (1997). Se midió la altura total (H) de las palmas para el 40 % de los individuos registrados en cada parcela siguiendo los protocolos nacionales (Yepes et al., 2015). Para los individuos con alturas inferiores a 15 m, se utilizó un mástil telescópico de fibra de vidrio (Hastings Sticks 3JF108823), y para alturas superiores, se empleó un hipsómetro láser (Nikon 550). En el caso de grandes árboles caídos, el volumen de fuste se estimó a través de la medición en campo de la longitud y los diámetros de los extremos. Se tomaron además, secciones transversales de individuos para estimar la densidad de la madera en el laboratorio. El material fue pesado in situ y fue llevado al Laboratorio de la Seccional Pitalito del Servicio Nacional de Aprendizaje (SENA) para la determinación del peso seco y la densidad, necesarios para estimar la necromasa (NM).
Ecuaciones de biomasa y preparación de los datos: Para la estimación de la biomasa aérea de los árboles (BA), se utilizaron las ecuaciones desarrolladas para Colombia por Álvarez et al. (2012), siguiendo la clasificación de zonas de Holdridge (1967), y que tienen como variables independientes el D y la densidad de la madera de las especies (ρ) (Cuadro 1). Para el uso de las mismas, fue necesario asignarle a cada individuo de las especies el valor de densidad de madera correspondiente, empleando los valores reportados por Zanne et al. (2009). Para aquellos taxones que no hacían parte de los listados consultados, se utilizó el valor promedio del género o familia correspondiente a la especie. Las estimaciones de la biomasa aérea de los helechos arbóreos y de las palmas, se realizó empleando las ecuaciones desarrolladas por Sierra et al. (2007), que fueron generadas para bosques tropicales de montaña del nordeste del departamento de Antioquia, y que presentan características biofísicas y ecológicas similares a las de los bosques del área de estudio. La altura de aquellas palmas que no fueron medidas en campo, fue estimada empleando un modelo exponencial (H = cDz; donde H es la altura total en m, D es el diámetro en cm, y z es el parámetro del modelo), que fue generado a partir de 1 645 registros de altura tomados en campo.
La biomasa aérea de cada parcela, se obtuvo como la sumatoria de la biomasa de los individuos registrados en ella. Finalmente, el valor obtenido se escaló a unidades de megagramos por hectárea (Mg/ha), y se estimó el valor promedio de biomasa aérea para los bosques del área de estudio. La biomasa subterránea (BS) o belowground biomass, fue estimada empleando la ecuación desarrollada por Cairns, Brown, Helmer, y Baumgardner (1997) y recomendada por el IPCC (2006) (Cuadro 1). La masa total (MT) fue calculada como la suma de los promedios de la biomasa aérea, la biomasa de raíces y la necromasa (NM). A partir de los resultados de masa total (MT), se realizó la estimación del carbono promedio almacenado siguiendo las recomendaciones del IPCC (2006), que sugieren calcular el carbono almacenado como 47 % de la biomasa.
Cuadro 1: Ecuaciones empleadas para las estimaciones de altura, biomasa aérea y raíces en los bosques del sur de los Andes Colombianos. Table 1: Equations used by high, aboveground biomass and belowground in the forests located in the South of the Colombian Andean
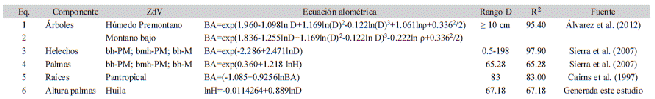
ZdV= zona de vida siguiendo la clasificación de Holdridge et al. (1971); BA= Biomasa aérea (kg); D= Diámetro (cm), ρ= Densidad de la madera (g/cm3); Rango D= rango diamétrico para el que aplica; R2= ajuste del modelo (%); H= altura total (m).
Otras características estructurales: El área basal total (m2/ha) de cada parcela, se obtuvo como la sumatoria del área basal de todos los individuos; el valor obtenido se extrapoló a una hectárea. A partir de estos resultados se calculó el área basal promedio para el bosque del área de estudio. Un procedimiento similar fue realizado para la estimación del número de árboles por hectárea (N/ha). Adicionalmente, se evaluó la distribución diamétrica de los bosques a través de la construcción de un histograma de abundancia vs. clases diamétricas.
Análisis de la información:Se calcularon los estadísticos por hectárea para las variables analizadas: área basal (m2/ha), número de árboles (N/ha), biomasa aérea, biomasa subterránea y necromasa (Mg/ha) de los bosques del área de estudio, al igual que los estadísticos respectivos de intervalo de confianza (IC 90 %) y coeficiente de variación (CV %). Adicionalmente, se calculó el error asociado (E %) a la estimación de la biomasa aérea y carbono con la ecuación: E (%)=[(t*Sy)/x]*100. Donde, Sy es la desviación estándar (Mg/ha), t es el estadístico t-student y x el promedio de la variable analizada. Con el fin de estimar las alturas de las palmas, se generó un modelo de alturas empleando técnicas de regresión lineal y no lineal, usando la transformación logarítmica y un factor de corrección. Todos los análisis estadísticos se realizaron con la versión 2.9.0 R del paquete estadístico utilizado (R Development Core Team, 2010).
Resultados
Riqueza y composición florística: Se registraron en total 12 180 individuos en 11 ha inventariadas, representados 46 familias, 95 géneros y 68 especies. Del total de los individuos, 50.5 % fueron clasificados al nivel de especie, 20.2 % al nivel de género, 0.01 % al nivel de familia, y 29.3 % quedaron como indeterminadas. En general las especies con mayor abundancia fueron C. excelsa (5 952 N/ ha), Alfaroa colombiana Lozano, Hern. Cam. & Espinal (4 768 N/ha), Quercus humboldtii (356 N/ha) y Billia rosea Planch. & Linden C. Ulloa & P. Jørg. (752 N/ha), las cuales presentaron además, altos valores de densidad de la madera (0.7-0.8 g/cm3). Del total de los individuos registrados, 10.8 % fueron palmas, 1.4 % helechos arbóreos y el 87.8 % restante, fueron especies arbóreas leñosas. En el Cuadro 2 se presenta el listado de las familias, géneros y especies más importantes en estos bosques, teniendo en cuenta la abundancia de las mismas en el ecosistema.
Cuadro 2: Familias, géneros y especies más abundantes (número de individuos totales) en los bosques del sur de los Andes Colombianos. Table 2: Families, gender and species more abundant (total number of individuals in 11 ha) in the forests located in the South of the Colombian Andean
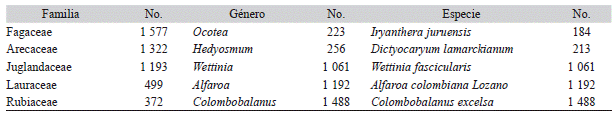
No: Número de individuos totales registrados por cada taxón en 11 ha muestreadas.
Estructura del bosque: Se encontró alta variabilidad en los parámetros estructurales del bosque. El área basal promedio fue de 37.9
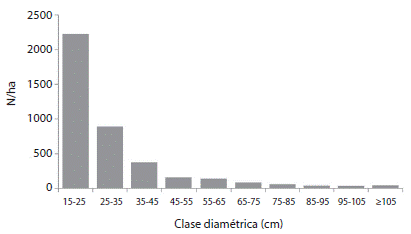
± 13.9 m2/ha. Seis de las 44 parcelas tuvieron áreas basales superiores a 55 m2/ha, siendo 75.4 m2/ha el mayor valor registrado. Estas parcelas dentro del muestreo aleatorio, fueron establecidas en sitios con aglomeraciones de la especie C. excelsa. El número de árboles por hectárea estuvo entre 368.8 ± 127.6 N/ha y tuvo relación significativa con el área basal (n= 42, F = 16.6, r2= 26.7, p= 0.0002). Respecto a la distribución diamétrica, se observó que ésta presenta forma de J invertida, la cual es típica de comunidades maduras y disetáneas (Figura 1).
Ecuaciones y biomasa del bosque: Las ecuaciones empleadas para la estimación de la biomasa aérea (Cuadro 1), tuvieron como variables explicativas el diámetro y la densidad de madera en el caso de individuos arbóreos, y el diámetro para helechos, mientras que para las palmas se usó la altura total (H). En contexto, la ecuación desarrollada para estimar la altura de las palmas que no fueron medidas en campo, presentó un buen ajuste a pesar de la variabilidad natural de los datos (n = 722, r2 = 67.2 %, DW = 1.3, p = 0.0001). Dicha variabilidad se debe principalmente a la presencia de tres especies de palmas: Ceroxylon quindiuense (H. Karst.) H. Wendl., Dictyocaryum lamarckianum (Mart.) H. Wendl., y Wettinia fascicularis (Burret) H.E. Moore & J. Dransf.
La biomasa aérea promedio del bosque fue 395.6 ± 55.2 Mg/ha, la cual es explicada en 99.4 % por el componente arbóreo leñoso, 0.3 % por las palmas y 0.3 % por los helechos arbóreos. Por su parte, la biomasa subterránea fue del orden de 74.3 ± 9.0 Mg/ha y la necromasa de 75.9 ± 19.9 Mg/ha. De esta, el 53.1 % fue aportado por los detritos gruesos caídos (DGC), y el 46.9 % por los árboles muertos en pie (AMP). La masa total del bosque (MT), compuesta por la biomasa área (BA), la biomasa subterránea (BS) y la necromasa (NM) fue de 545.9 ± 84.1
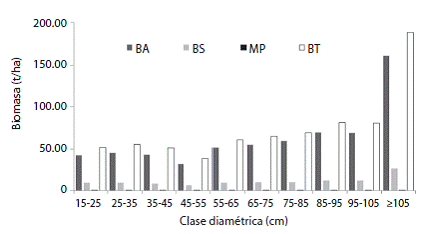
Mg/ha (Cuadro 3). Los robledales del sur del Huila almacenan en promedio un total de 272.9 ± 42.0 MgC/ha, de los cuales 72.5 % se encuentran almacenadas en la biomasa aérea, 13.6 % en la biomasa subterránea de raíces, y 13.9 % en la necromasa. En relación con la distribución de la biomasa aérea por clase diamétrica, se observó que ésta se encuentra concentrada principalmente en los árboles con diámetros grandes (e.g. 95 - 105 cm y ≥ 105 cm; Figura 2).
Cuadro 3: Estimaciones de biomasa y carbono en diferentes compartimientos en los bosques del sur de los Andes Colombianos. Table 3: Estimates of biomass and carbon in different pools in the forests located in the South of the Colombian Andean
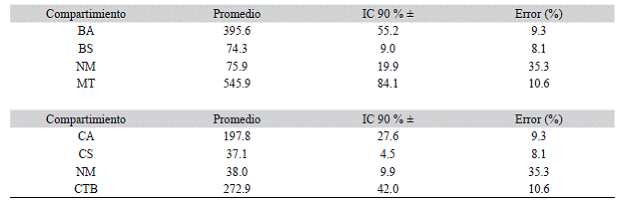
BA: biomasa aérea (Mg/ha); BS: biomasa subterránea (Mg/ha); NM: necromasa; MT: masa total del bosque (Mg/ha); CA: carbono almacenado en la biomasa aérea (MgC/ha); CS: carbono subterráneo (MgC/ha); CTB: carbono total almacenado en los bosques (MgC/ha).
Discusión
Los bosques tropicales de montaña son considerados los ecosistemas más biodiversos del mundo (Myers, Mittermeier, Mittermeier, da Fonseca, & Kent, 2000) debido a que la variedad en las condiciones climáticas, orográficas y edáficas, favorecen el endemismo de especies y la co-existencia de ellas incluso en gradientes altitudinales cortos (Richter, 2008; Girardin et al., 2013). En el área de estudio, los resultados de esta investigación indican que se pueden encontrar alrededor de 68 especies en 11 ha, que incluyen individuos de porte arbóreo, palmas y helechos. La especie dominante es C. excelsa con 5 952 N/ha, una especie de la familia Fagaceae endémica de la zona andina de Colombia (Parra et al., 2011). Un número similar de especies, se encuentra en bosques de montaña en Antioquia, donde la especie dominante es Q. humboldtii (León et al., 2009). Las demás especies registradas, coinciden en su mayoría con las reportadas previamente por González et al. (2007) donde se caracterizó la estructura y composición florística de la vegetación del corredor biológico entre los PNN Puracé y PNN Cueva de los Guácharos. En relación con el área basal y el número de árboles por hectárea, el promedio obtenido fue de 37.9 ± 13.9 m2/ha y 368.8 ± 127.6 N/ha, respectivamente. Dichos valores se encuentran dentro de los rangos reportados en otros bosques tropicales de montaña (Álvarez, García, Krasilnikov, & García, 2013). No obstante, el límite superior en ambos casos, solo es comparable con los valores reportados en bosques tropicales de tierras bajas de la Amazonia y Asia (Spracklen & Righelato 2013).
En relación con la estructura diamétrica, el bosque de la zona de estudio presenta una estructura disetánea con una distribución por tamaño diamétrico balanceada, en la cual hay mayor cantidad de individuos en las categorías diamétricas inferiores; en este caso en la clase de 15 - 25 cm de diámetro. Esta condición está determinada por las características de las especies dominantes, las cuales presentan mayores individuos en las clases diamétricas más bajas (León et al., 2009). De esta manera, si se tiene en cuenta que los bosques del sur del Huila están dominados por individuos de C. excelsa, se podría pensar que esta especie es la que condiciona y determina dicha estructura. Resultados similares se han descrito para bosques de roble dominados por la especie Q. humboldtii en el departamento de Antioquia. Sin embargo, de acuerdo con Parra et al. (2011) aunque la estructura diamétrica de C. excelsa es en forma de J invertida, aun incluyendo individuos con D ≥ 1 cm, no es suficiente para asegurar la permanencia de la especie en el tiempo, dado que los renuevos no alcanzan el número y la altura requerida para ello. Esto ocurre por la alta mortalidad natural de las plántulas recién germinadas, y sugiere que la especie podría estar en riesgo de persistencia (Parra et al., 2011). Teniendo en cuenta que en el presente estudio el muestreo solo incluyó árboles con D ≥ 15cm, se deja abierta la hipótesis de Parra et al. (2011) para ser probada en futuras investigaciones.
Respecto a la BA, el valor estimado tuvo un error asociado de 9.3 %, y superó el valor reportado en las estimaciones nacionales que se habían realizado para Colombia empleando técnicas de teledetección y datos de campo (395.6 ± 55.2 Mg/ha). Por ejemplo, Anaya, Chuvieco y Palacios (2009) reportaron una biomasa aérea de 264 Mg/ha para los bosques maduros nacionales, mientras que Saatchi et al. (2011) encontraron una biomasa promedio total (aérea+subterránea) de 282 Mg/ha, de la cual 200 Mg/ha correspondió a biomasa aérea. Más recientemente, Phillips et al. (2014) reportaron un promedio nacional de 383.3 ± 3.9 Mg/ha empleado información de inventarios de campo generados durante el período 1990 - 2012, y que incluye la medición de individuos con D ≥ 10 m. De otro lado, el valor obtenido en esta investigación fue menor y más cercano al de un estudio previo realizado en el área de estudio, donde se concluyó que la BA promedia podría alcanzar hasta 448.1 ± 183.6 Mg/ha cuando se incluyeron todos los árboles con D ≥ 10 cm (Yepes et al., 2015). No obstante, vale la pena anotar que el valor obtenido en este estudio a pesar de que no consideró individuos con 1 ≥ D > 15 cm, si mostró una menor variabilidad en el intervalo de confianza, al emplear mayor cantidad de unidades muestrales (26 vs. 44 parcelas). De cualquier modo, es claro que los resultados de ambos estudios, a pesar de sus diferencias, sugieren que los bosques tropicales de montaña, tienen una participación importante en el potencial de almacenamiento de carbono a nivel nacional, que se debería considerar al momento de priorizar zonas para la implementación de acciones tempranas en el marco de la Estrategia Nacional para la Reducción de la Deforestación y Degradación (ENREDD+) en Colombia, de manera similar a lo que se ha hecho con los bosques de la Amazonía colombiana, para los cuales ya se ha sometido a la Convención Marco de las Naciones Unidas sobre el Cambio Climático (CMNUCC) un primer nivel de referencia de emisiones forestales en el marco de REDD+ (UNFCCC, 2014). De esta manera, los resultados de esta investigación, aportan un elemento adicional para toma de decisiones en relación con las estrategias de conservación de los bosques tropicales de montaña, si se tiene en cuenta que este tipo de ecosistemas han estado sujetos por años a la presión por deforestación (Armenteras et al., 2003, González et al., 2011; IDEAM 2014), y que además, contienen especies forestales valiosas desde el punto de vista ecológico y funcional (Van Der et al., 2009).
Por otro lado, los resultados obtenidos muestran que la biomasa aérea de los bosques tropicales de montaña puede ser similar a la documentada para bosques húmedos tropicales de tierras bajas como los de la Amazonía, Borneo y países africanos como Uganda y Kenia (288.6 Mg/ha, 457.1 Mg/ha, 603.9 Mg/ ha, y 607 Mg/ha, respectivamente), donde los valores de biomasa aérea son superiores a 250 Mg/ha (Malhi et al., 2006; Slik et al., 2010; Lewis et al., 2013; Omoro, Starr, & Pellikka, 2013). Los altos valores de biomasa aérea encontrados en la presente investigación, pueden estar relacionados con la abundancia de árboles de gran tamaño (D ≥ 70 cm) de la especie C. excelsa, que aportan hasta 81% de la biomasa aérea total del bosque (Yepes et al., 2015), al igual que ocurre en los bosques de Asia dominados por especies de Fagáceas (Culmsee et al., 2010). En general, los árboles grandes desempeñan un papel importante en los bosques tropicales de montaña, y determinan los patrones de distribución de la biomasa y carbono a escalas de paisaje (Slik et al., 2010; Slik et al., 2013; Larjavaara & Muller-Landau, 2013), al igual que lo hacen en bosques de tierras bajas de la Amazonia (Malhi et al., 2006). Sin embargo, estos árboles se encuentran altamente amenazados por la tala selectiva ilegal que se presenta en el área de estudio, a partir de alteraciones localizadas, que podrían poner en peligro su papel como reservorios de carbono.
En relación con los demás compartimientos, los estimados obtenidos para la biomasa subterránea y necromasa, tuvieron un error de estimación de 9.0 % y 19.9 % respectivamente. En el caso de la biomasa subterránea los valores obtenidos fueron cercanos a los reportados por Sierra et al. (2007) para un bosque tropical de montaña en el noroeste de los Andes colombianos (83.7 ± 17.2 Mg/ha), pero inferior al hallado para bosques de tierras bajas en la Amazonia brasileña (89 Mg/ha; Nogueira et al., 2008) y Singapur (86.3 Mg/ha; Ngo et al., 2013), si se considera únicamente el valor promedio. Con relación a la necromasa, los valores encontrados en este estudio fueron superiores a los obtenidos para un bosque con condiciones biofísicas similares en los Andes colombianos (14.7±2.2 Mg/ha; Sierra et al., 2007), a los reportados para bosques del oeste de la Amazonía sobre suelos ricos en arcilla y de arenas blancas (30.9-45.8 Mg/ha; Chao, Phillips & Bakera, 2008), y para los bosques montanos de Ecuador (Wilcke et al., 2005). No obstante, se encuentra en el rango conocido para bosques de tierras bajas (Baker et al., 2007), y muy cercanos a los reportes existentes para bosques tropicales de montaña en el departamento de Antioquia (62.2-80.2 Mg/ha; Navarrete et al., 2011). Es de anotar, que en este estudio la mayor proporción de necromasa fue aportada por los detritos gruesos caídos (DGC), lo cual al parecer es característico de bosques tropicales de montaña (Wilcke et al., 2005; Navarrete et al., 2011), mientras que en bosques tropicales de tierras bajas, el mayor aporte lo realizan los árboles muertos en pie (Baker et al., 2007; Chao et al., 2008). Algunos estudios han concluido que la cantidad de necromasa en un ecosistema depende de diferentes factores como la edad, la estructura y la biomasa aérea del mismo. Otros factores, tal como eventos naturales extremos (e.g. tormentas, huracanes), también podrían tener un impacto en el flujo de carbono almacenado en la necromasa (Ifo, Koubouana, Jourdain, & Nganga, 2015). En el caso de los bosques del sur del Huila, la baja tasa de descomposición y la abundancia de árboles grandes que al morir aportan gran cantidad de detritos gruesos, puede ser un factor que determina el patrón observado.
En términos de biomasa y carbono total almacenado, la estimación tuvo un error total de 10.6 %, y se observó que el valor obtenido (545.9 ± 84.1 Mg/ha) fue superior a los que se encuentran en la literatura para bosques maduros o primarios de los Andes colombianos (incluyendo el compartimiento de suelo; 383.7 ± 55.5 Mg/ha; Sierra et al., 2007), los bosques tropicales de tierras bajas de la Amazonía (449.8 Mg/ha; Nogueira et al., 2008), o en los bosques tropicales de tierras bajas en Singapur (451.6 Mg/ha; Ngo et al., 2013). Esto se explica principalmente por tres razones: i) muchos estudios relacionados con la estimación de biomasa y carbono, solo incluyen el componente de biomasa aérea subestimando el aporte de los demás compartimientos; ii) cuando se incluye la biomasa subterránea, se ha visto que ésta es mayor en bosques montanos que en los bosques de tierras bajas (Sanford & Cuevas, 1996), al igual que el aporte de la necromasa, debido a las bajas tasas de descomposición en este tipo de ecosistemas (Powers et al., 2009; Bruijnzeel, Scatena, & Hamilton, 2011); y iii) las características del microclima, tipo de suelos y la dominancia de ciertas especies, pueden condicionar el patrón de distribución de biomasa a escalas locales (Culmsee et al., 2010; Slik et al., 2010; Slik et al., 2013; Girardin et al., 2013; Larjavaara & Muller-Landau, 2013).
Los resultados resaltan además, que la inclusión en la estimaciones totales, de los contenidos de carbono que se almacenan en otros compartimientos diferentes a la biomasa aérea, es importante porque refleja de manera más realista el potencial de almacenamiento de carbono en los bosques tropicales (IPCC 2006; Sierra et al., 2007; Nogueira et al., 2008; Ngo et al., 2013), y su papel en el ciclo global del carbono. Por ello se recomienda que su estimación o cuantificación, sea incluida en iniciativas REDD+ en las diferentes escalas y mercados (voluntario o regulado). De otro lado, aunque REDD+ como enfoque de mitigación ante la CMNUCC representa una oportunidad sin precedentes para la conservación de la biodiversidad forestal, existe una preocupación generalizada en torno a la posibilidad de resultados ambientales negativos, si no se da una consideración adecuada de la biodiversidad en todo el proceso (Gardner et al., 2012). De acuerdo con algunos autores (Venter et al., 2009, Gardner et al., 2012, Duque et al., 2014) priorizar la conservación de los bosques en virtud únicamente de los contenidos de carbono, podría poner en peligro a ecosistemas que presentan alto endemismo y diversidad, pero bajos contenidos de carbono. Al respecto, se considera que los bosques del sur del Huila se podrían ver beneficiados por iniciativas de este tipo, donde además de disminuir la deforestación histórica en la zona, se conservarían especies importantes de flora y fauna que cumplen roles ecológicos vitales para el ecosistema en general, como lo es C. excelsa o roble negro.
Se concluye entonces, que las estimaciones que consideran únicamente la biomasa aérea, subestiman en gran medida las existencias de carbono en los ecosistemas forestales, ya que otros compartimientos pueden contribuir con más del 20 % del total, al igual que ocurrió en el presente estudio en bosques tropicales de montaña, donde es evidente su papel como reservorios de carbono. Finalmente, se considera que es clave realizar esfuerzos para estudiar el balance global de carbono y mitigar los efectos del cambio climático, a través de enfoques de paisajes heterogéneos que se han originado como consecuencia de las alteraciones antropogénicas y la dinámica natural de los bosques, y no solo en función de un ecosistema en particular. Estudios a nivel de paisaje, mejorarían la capacidad de abordar las preguntas globales relacionadas con el balance de carbono en los bosques, y tomar decisiones más acordes con las realidades y circunstancias nacionales de un país.

