










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.63 n.1 San José Jan./Mar. 2015
Life-stages, exploitation status and habitat use of Lutjanus goreensis (Perciformes: Lutjanidae) in coastal marine environments of Lagos, SW Nigeria
Ciclo de vida, estado de explotación y hábitat de Lutjanus goreensis (Perciformes: Lutjanidae) en ambientes marinos costeros de Lagos, suroeste de Nigeria
Ciclo de vida, estado de explotación y hábitat de Lutjanus goreensis (Perciformes: Lutjanidae) en ambientes marinos costeros de Lagos, suroeste de Nigeria
Kafayat, A. Fakoya1*, Martins, A. Anetekhai1, Shehu, L. Akintola1, Abdulwakil, O. Saba1 & Mikhail A. Abass2*
Abstract
The Gorean snapper, Lutjanus goreensis is an important component of artisanal fisheries and trawl landings in the Gulf of Guinea. Despite its economic importance, there is a dearth of information on size structure and life history strategies of the species. Therefore, the objectives of this study were to provide baseline data on the life stages, exploitation status and habitat use for the species in Nigeria. Monthly samples were obtained from artisanal and trawl catches in Five Cowrie Creek and Lagos coastal waters between December 2008 and December 2010, respectively. Length-frequency distributions of the fishes caught were analysed to provide preliminary information on mean and modal lengths at capture and life - history strategies based on habitat use and estuarine-dependency for L. goreensis. A total of 822 specimens of L. goreensis were collected from Five Cowrie Creek while 377 specimens were collected from Lagos coastal waters. Total length varied between 7.90-34.90cm for creek samples and from 21.90-56.10cm for marine samples. Length-frequency histograms showed polymodal size distributions in creek and marine samples. Length-frequency distributions of L. goreensis showed a high abundance of juveniles (<20cm) and sub-adults (20-35cm) which accounted for 84.1% and 68.4% of creek and marine samples examined, respectively. For the creek samples, fish in modal length class of 13.00-13.99cm were the most exploited while in the marine samples, length classes of 29.00-30.99cm and 31.00-32.99cm constituted the most frequently exploited fishes. Increase in total lengths from the creek (mean±SD; 16.19±3.73cm) to the marine habitat samples (32.89±6.14cm) indicated ontogenetic shift in habitat use. Occurrence of a predominant juvenile population in Five Cowrie Creek by L. goreensis suggests estuarine-dependency and is indicative of a temporary juvenile habitat or a migratory corridor. In conclusion, data from the presently reported study and previous studies demonstrated that juvenile L. goreensis displays estuarine dependency and habitat flexibility. Hence, this underscores the importance of preserving estuarine environments as essential fish habitats to prevent overfishing. The study also concludes that the species is vulnerable to recruitment overfishing in the marine environment especially as a consequence of shrimping. Consequently, it advocates for ban on all fishing activities during peak spawning periods in breeding grounds and shrimp ground assemblage.
Key words: Lutjanus goreensis, length-frequency distribution, habitat use, life-stages, estuarine dependency, exploitation status.
Resumen
El pargo Lutjanus goreensis es un componente importante de la pesca artesanal y de arrastre en el Golfo de Guinea. A pesar de su importancia económica, hay una escasez de información sobre la estructura de tamaño y las estrategias de historia de vida de las especies. Por lo tanto, los objetivos de este estudio fueron proporcionar datos de referencia sobre las etapas del ciclo de vida, el estado de la explotación y el hábitat de la especie en Nigeria. Se realizaron recolectas mensuales de capturas artesanales y de arrastre en Five Cowrie Creek y aguas costeras de Lagos en diciembre 2008 y diciembre 2010, respectivamente. Se analizaron las distribuciones de frecuencia de tallas de los peces capturados para proporcionar información preliminar sobre la media y longitudes modales en la captura y las estrategias de historia de vida basado en el uso del hábitat y la dependencia a los estuarios para L. goreensis. Un total de 822 ejemplares de L. goreensis se obtuvieron de Five Cowrie Creek, mientras que 377 muestras de las aguas costeras de Lagos. La longitud total varió entre 7.90-34.90cm para muestras del estuario y 21.90-56.10cm para marinas. Histogramas de frecuencia de talla mostraron distribuciones de tamaño polimodales en muestras estuarinas y marinas. Distribuciones de frecuencia de talla de L. goreensis mostraron una alta abundancia de juveniles (<20cm) y subadultos (20-35cm) que representaron el 84.1% y el 68.4% de los estuarios y las muestras marinas examinadas respectivamente. Para las muestras del estuario, peces de talla 13.00-13.99cm fueron los más explotados, mientras que en muestras marinas, las tallas de 29.00-30.99cm y 31.00-32.99cm constituyeron los peces más frecuentemente explotados. Un aumento de la longitud total de la quebrada (media±SD; 16.19±3.73cm) para el hábitat marino (32.89±6.14cm) indicó un cambio ontogenético en el uso del hábitat. La aparición de una población juvenil predominante en Five Cowrie Creek por L. goreensis sugiere una estuario-dependencia y es indicativa de un hábitat juvenil temporal o un corredor migratorio. En conclusión, los datos del estudio actual y estudios previos demostraron que juveniles de L. goreensis presentan una dependencia a los estuarios y una flexibilidad en el uso de hábitat. Por lo tanto, esto subraya la importancia de preservar los ambientes estuarinos como hábitats esenciales de peces para evitar la sobrepesca. El estudio también concluye que la especie es vulnerable a la sobrepesca de reclutamiento en el medio marino, especialmente como consecuencia de la pesca de camarón. En consecuencia, se aboga por la prohibición de todas las actividades pesqueras durante los períodos de mayor puesta de huevos.
Palabras clave: Lutjanus goreensis, distribución de frecuencia del largo, uso de hábitat, estapas de vida, dependencia estuarina, status de explotación.
Lutjanidae popularly known as snappers comprises a large family of the order Perciformes, important in both tropical and subtropical waters. In the Gulf of Guinea, snappers occur as incidental catches in trawl landings and are important components of the local subsistence fisheries (Allen, 1985). The Gorean snapper, Lutjanus goreensis (Valenciennes, 1830) is a medium to large and deep-bodied species of snappers (over 35cm) (Martinez-Andrade, 2003) common to 50cm but capable of attaining maximum total length of 80cm (Allen, 1985). It belongs to the subthermocline Lutjanid community in the Gulf of Guinea constituting part of the coastal demersal fisheries (Longhurst, 1969). Adults inhabit sandy, rocky or corally areas in the marine environment (Allen, 1985; Newman, 1995) while juveniles inhabit mangrove estuaries, creeks, coastal rivers and lower reaches of freshwater (Thys van den Audanaerde, 1966; Allen, 1985; Ezenwa, Alegbeleye, Anyanwu, & Uzukwu, 1990; Agboola, & Anetekhai, 2008).
Population structure of fish depicted in length-frequency distribution provides useful information that can aid fisheries management. It is fundamental to understanding growth, reproduction and recruitment with changes in size as an early indicator of disturbance (Johnson, & Tamatamah, 2013). Shifts in length composition of a fished population over time often reflect trends in the intensity of harvest and can be examined qualitatively to provide general and preliminary information on the dynamics of a stock (Heery, 2007). Mean sizes of fish catch derived from length frequency distributions are also better indicators than catch rates for use in stock assessment because they change in a more predictable manner with abundance (Kell, Bonhommeau, & Fromentin, 2013).
Spatial separation of size-classes of fish species suggests movement from one habitat to another with ontogeny (Cocheret de la Morinie`re, Pollux, Nagelkerken, & van der Velde, 2003). Ontogenetic migrations have been inferred from spatial separation of size - classes in many fish species including lutjanids such as Lutjanus argentimaculatus (Sheaves, 1995); L. jocu (Moura, Francini-Filho, Chaves, Minte-Vera, & Lindeman, 2011); L. argentiventris (Aburto-Oropeva, Dominguez-Guerrero, Cota-Nieto, & Plomozo-Lugo, 2009); L. apodus and L. griseus (Hammerschlag-Peyer, Allgeier, & Layman, 2013).
The concept of “estuarine dependence” acknowledges that fish use of estuarine and oceanic habitats is a continuum. An estuarine-dependent species is defined as one in which estuaries, or similar habitats, are the principal environments for at least part of the life cycle and without which a viable population would cease to exist (Nagelkerken, & van der Velde, 2002; Able, 2005).The concept of estuarine dependence is also often related to nurseries and it has so far been tested by comparing presence/absence of juvenile and adult fishes in estuaries versus offshore habitats (Cocheret de la Morinie`re, Pollux, Nagelkerken, & van der Velde, 2002 ; Nagelkerken, & van der Velde, 2002).
Notwithstanding its economic importance, studies related to size structure, life-stages, habitat use, ontogenetic habitat shift and estuarine -dependency of L. goreensis in the estuarine and marine environments are either poorly documented or lacking. Globally, several species of shallow and deepwater snapper stocks are vulnerable to over-fishing with serious consequences for conservation and management (Grandcourt, Abdessalaam, & Franklin, 2006). Though, exploitation status of the snapper species is suggested to be over-exploited in Nigerian coastal waters (Amiengheme, 1997, 2001), there is a dearth of information on the size structure of L. goreensis. Available data from previous studies of Fakoya, Abass, Owodeinde, Lawson, and Ojo (2010); Orhibhabor and Ogbeibu (2010) in estuarine habitats consist of low sample sizes.
Therefore, this study is unique in that it provides the first data on the life-stages, exploitation status and habitat use of the Gorean snapper, L. goreensis from comparative analysis of the length-frequency distributions in estuarine and marine environments, respectively. Thus, the objectives of this study are to give information on the mean and modal lengths at capture, to describe distinct life-stages viz-a- viz: juvenile, sub-adult and adult and lastly, to define life-history strategies based on habitat use and estuarine - dependency of L. goreensis.
Materials and Methods
Study sites: This study was carried out in Five Cowrie Creek and Lagos coastal waters, South-West, Nigeria as shown in Fig. 1. Five Cowrie Creek is one of the numerous adjoining creeks to the Lagos Lagoon. It is connected to the Lagoon at two ends; the Lagos Habour which opens to the inshore (coastal) waters off Lagos and at the extreme of the Eastern part of Ikoyi, respectively. The creek has an approximate length of 7km and is located 6°26’24’’ N - 3°24’18’’ E. It is deep, tidal and subjected to the same physical conditions which are regulated by rainfall and salinity variations as the harbour (Onyema, Nwankwo, & Oduleye, 2005/2006). Seawater enters into the creek through the Lagos Harbour end at high tide and at low tide; water is drained from the Eastern part of the Lagos Lagoon through the creek to the harbour en-route to the Atlantic Ocean (Nwankwo, Okedoyin, & Adesalu, 2012). The shoreline of the creek has a brackish/fresh water swamp almost completely devoid of original mangrove vegetation except for occasional stands of Casuarina equisetifolia, Terminalia catappa and sparse occurrence of grasses Chromolaena odorata (Adekanmbi, & Ogundipe, 2009).
The fishing grounds of Lagos coastal (inshore) waters extend approximately from 6°24’54’’ N - 3°23’06’’ E covering the Nigerian-Benin Republic Border in the West to as far as Lekki in the East. To the North, it is bounded by Five Cowrie Creek and to the South by the Atlantic Ocean (Ediang, & Ediang, 2013). Off the Lagos Coast, the continental shelf is relatively narrow about 15km and predominantly soft and muddy (Tobor, 1991). Most demersal fish stocks off the Lagos Coast are concentrated in an area of about 1 800 km2 which is located between the high-water mark on the shore and lower limit of the thermocline (Ssentongo, Ukpe, & Ajayi, 1986).
Fish sampling: Fish samples were purchased from licensed trawlers (cod-ends with stretch mesh size of 44mm and 76mm of shrimp and fish trawl nets, respectively operating in the marine coastal waters of Lagos. Artisanal fishermen using hook and line exclusively were the sources of fish samples from Five Cowrie Creek and also additional samples from marine coastal waters, respectively. Fish specimens were identified with the aid of taxonomic keys of Allen (1985); Fischer, Bianchi and Scott (1981). Each fish sample was selected without bias on size, such that each fish size in the population had an equal chance of being selected. The samples were transported in a cooler packed with ice to the laboratory, where they were immediately analyzed fresh or frozen for later examination. In the laboratory, frozen specimens were sufficiently thawed and wiped to remove excess moisture.
Size-distributions, life-stages, lengths at capture and habitat use: Samples of L. goreensis from the creek and Lagos coastal waters were measured (total length, TL to the nearest 0.1cm) using the traditional fish measuring board and weighed (whole body weight to the nearest 0.1g) with an electronic balance (Mettler PM 400). Each fish specimen was cut open and sex determined by observing the gonads as there is no sexual dimorphism in this species. To minimize systematic error due to collection from commercial catches, data from all sampling dates which were assumed to be representative for the population were pooled together for size-frequency study. Data on total length were grouped using 1cm and 2cm total length intervals for creek and marine samples. For data analyses, percentage frequency of different size classes and total lengths were used for the length-frequency distribution. Length-frequency histograms were constructed to determine the size distributions of the sampled populations and then analyzed by sex and gear.
The life stages (adult, sub-adult and juvenile) of L. goreensis were determined based on total length. Juvenile and sub-adult life stages were based on size classes of L. argentiventris (Aburto-Oropeva et al., 2009); L. apodus and L. griseus (Hammerschlag-Peyer et al., 2013) as: small-sized or juveniles (<20cm); medium-sized or sub-adults (20-30cm). The large-sized or adult life-stage was estimated based on size at first sexual maturity which was defined as 43% of maximum length for shallow-water and continental snapper species (Grimes, 1987). The maximum length for the species was based on a maximum total length of 80cm in the Gulf of Guinea for the species (Allen, 1985). Hence, all fish less than 35cm but greater than 30cm were also included as sub-adults in this study. This size-class of >35cm corresponded to large-adults in L. peru (Reddy et al., 2013).
Mean and modal lengths at capture were estimated for the species in the two habitats to assess exploitation status of the species. Length-frequency distributions of sampled fishes in the two habitats were used to infer habitat use. The species was considered to undergo possible ontogenetic habitat shifts based on mean total length. If the mean total length of a fish species from the marine habitat was significantly longer than that in the juvenile habitat (Student’s t-test, p<0.05), then the species may exhibit ontogenetic habitat shift (Moura et al., 2011; Honda, Nakamura, Nakaoka, Uy, & Fortes, 2013). Comparison of the presence/absence of juvenile and adult fishes in juvenile versus marine habitats facilitated classification of estuarine-dependency in the species. The species was classified as estuarine-dependent if it was found offshore as adults and occurred as juveniles only in estuaries (Nagelkerken & van der Velde, 2002).
Data analyses were performed by IBM SPSS statistics version 20. The data on total lengths were analysed using descriptive statistics and expressed as mean ± standard deviation, mode and percentage. Student’s t-test (α=0.05) was used to assess for significant differences in mean total lengths: (i) between creek and marine samples of Lutjanus goreensis and (ii) between trawl caught fish and artisanal caught fish.
Results
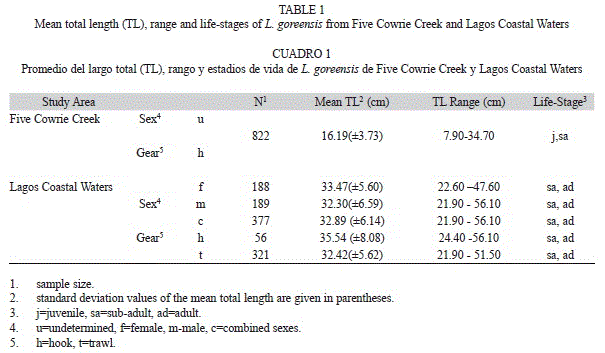
Size-distributions and life-stages: Information relating to size distributions and life- stages of creek and marine Gorean snapper, L. goreensis are given in Table 1. A total of 822 specimens were sampled from the creek while 377 specimens were examined from the coastal waters. Total lengths of creek samples ranged from 7.90cm to 34.70cm and body weight ranged from 9.51g to 695.60g whereas composite ranges for marine samples varied between 21.90cm and 56.10cm total length and from 156g to 2 975g body weight. Juveniles were more prominent in the creek while sub-adults were predominant in the marine habitat. Fish length classes of creek samples were categorized into small-size group (84.1% of total sample) and medium-size group (15.9%) consisting the remaining samples. Fish with sizes ranging from 7.90-19.99cm and 20.00-34.99cm were believed to be juveniles and sub-adult members of the species, respectively.
In marine samples, fish length classes were categorised into medium-size and large-size groups. Fish less than 20cm were absent while sub-adults (68.4% of total sample) predominated over large fish (31.6%) and was subsequently referred to as adults.
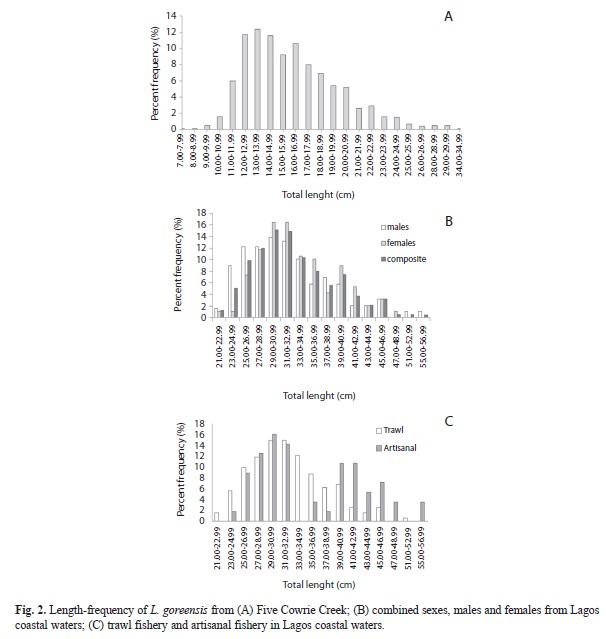
Lengths at capture: The length-frequency histograms showed polymodal distributions and percentage frequency of different size classes in samples of L. goreensis from the creek and coastal waters. Creek samples caught with hook by the artisanal fishery were largely unsexed and had a mean of 16.19cm ±3.73 standard deviation. By comparison, marine samples caught with the same gear had a mean of (35.54±8.08cm) (Table 1), indicating significant variations in sizes of the fish species in the two ecological habitats.
In creek samples modal class of 13.00-13.99cm with a frequency of 12.4% was the most exploited (Fig. 2A). Length classes of ≤9.00cm and 34.00-34.99cm with frequencies of 0.2% and 0.1% respectively, accounted for the least exploited fishes. Composite marine samples had a mean length of 32.89±6.14cm (Table 1). Modal class of 29.00-30.99cm was the most exploited with a frequency of 15.1% followed closely by the length class of 31.00-32.99cm with a frequency of 14.9% (Fig. 2B). Approximately, 58.1% of total sample was <33.00cm thus representing a majority of size classes fished below the modal class. Males (n=189) ranged from 21.90 to 56.10cm with a mean of 32.30±6.59cm while females (n=188) ranged from 22.60 to 47.60cm with a mean of 33.47±5.60cm (Table 1). There was no significant difference in mean total lengths observed between males and females (Student t-test, p>0.05). In males and females, specimens of 29.00-30.99cm and 31.00-32.99cm were also the most exploited (Fig. 2B).
Analysis by gear revealed that the bulk of the captured fish came from the trawl fishery (n=321) and varied in length from 21.90 to 51.50cm (32.42±5.62cm), while fish from the artisanal fishery (n=56) varied from 24.40 to 56.10cm (35.54±8.08cm) (Table 1). The Student’s t-test indicated a significant difference in the mean total lengths of trawl caught fish and artisanal caught fish (p<0.05) with larger sizes occurring in the latter. Modal classes of 29.00-30.99cm and 31.00-32.99cm represented the most exploited fishes with equivalent frequencies of 15.0% by trawl fishery. In artisanal fishery, the modal class of 29.00-30.99cm with a frequency of 16.1% was the most exploited followed closely by the length class of 31.00-32.99cm with a frequency of 14.3% (Fig. 2C).
Habitat use: Minimum sizes in the marine habitat (21.90cm) were always larger than minimum sizes in the estuarine habitat (7.90cm), indicating that the smallest individuals of the species occur only in the creek and not in the marine habitat. Similarly, maximum size in the marine habitat (56.10cm) was always larger than the maximum size in creek (34.90cm), indicating that the largest individuals occur only in the marine habitat.
Mean size of the fish was significantly higher in the marine habitat (32.89±6.14cm) than in the creek (16.19±3.73cm) (Student’s t-test, p<0.5) showing clear ontogenetic shifts in habitat use by L. goreensis. The species also showed estuarine-dependence in its juvenile stage. Juvenile life-stage of the species (<20cm) was the most dominant (84.1% of total sample) in the creek and showed gradual movement over size range of 20-35cm to the marine habitat where the species occurred only as sub-adults and adults.
Discussion
The largest size of the species from Five Cowrie Creek was much larger than the largest fish previously reported in other studies for Nigeria (Lawson, 2005; Orhihabor, & Ogbeibu, 2010; 2012), Ghana (Aheto et al., 2011; Okyere, Aheto, & Aggrey-Fynn, 2011); Bamboung, Senegal (Faye, Le Loc’h, Thiaw, & Tito de Moraïs, 2012) and Gabon (Mamonekene et al., 2006). Differences in size distributions of L. goreensis noted in the present study and previous studies is as a result of the larger sampling size collected in this study, differential gear selectivity – hook and line in this study against gillnet (stretched mesh size of 10-50mm) in Gabon (Mamonekene et al., 2006); pole seine net (stretched mesh size 5mm) in Ghana (Okyere et al., 2011); fishing pressure and lengths of sampling period. These factors, notably sampling size and gear selection, affected the size distributions of fish collected. Similarly, selectivity of the fishing gear used in collecting the specimens may be said to be biased particularly for fish above 8.00cm otherwise the absence of very small sizes can be ascribed to the absence of small size fishes less than 7.90cm in the creek.
The present study indicated that juveniles consisted of the most dominant life-stage in the creek while sub-adults predominated in the marine habitat. Creek samples were largely unsexed because of their small size and grossly undeveloped state of gonads. Previous studies classified L. goreensis from estuarine/brackishwater environments as juveniles (Lawson 2005; Orhihabor, & Ogbeibu, 2010, 2012), in coastal wetlands systems of Ghana (Aheto et al., 2011; Okyere et al., 2011) and in Complex of Protected Areas of Gabon (Mamonekene et al., 2006). Results of the present study include a sub-adult life-stage which constituted an insignificant percentage of the main creek population. Individuals in the sub-adult life-stage were either sexually inactive or immature individuals from macroscopic examination of gonads. In contrasts, previous works of Sheaves (1995); Riley (2002) and Martinez-Andrade (2003) did not recognize sub-adults but mentioned large snappers in estuaries as functional juveniles and sexually immature.
Analysis of mean, minimum and maximum total lengths by gear strongly implied that the trawl fishery selected smaller fish than artisanal fishery, though the modal lengths of the fish caught were similar. Few marine samples measured up to 50cm which is the size commonly encountered for the species in the Gulf of Guinea. The maximum size observed in this study from the marine habitat was also below the maximum total length of 80cm reported for the species in the region (Allen, 1985). This implied that larger sizes above 50cm are becoming rare in commercial catches.
Appearance of adult sizes in trawl and artisanal catches suggested that the species probably moves to shallower waters near the coast where the fisheries occur. García-Contreras, Quiñónez-Velázquez, Morán-Angulo, & Valdez-Pineda (2009) observed a similar shoreward migration by the Amarillo snapper, L. argentiventris. A plausible reason for the onshore migration could be that in Nigerian waters, increase in fish catch is linked to the rainy season between March and October, thus also bringing more large-size fishes. Shoreward movement of the species could be to avoid disturbances caused by wind stress forcing in the open sea during the heavy rains as hypothesised for L. analis (Manjarrés-Martínez, Gutiérrez-Estrada, Mazenet-González, & Soriguer, 2010). Heavy rains in Nigeria occur between May and September, often causing the sea to be rough. Another factor could be that the peak spawning period for lutjanids coincides with maximum rainfall and minimum water temperatures so that there is increased vulnerability of reproductive adults to fishing. In many shallow-water fisheries targeting snappers, fishing takes advantage of concentrations of reproductive adults which undergo spawning migrations, spawning aggregations and post-spawning migrations (Claro, de Mitcheson, Lindeman, & García-Cagide, 2009; Fernandes, de Oliveira, Travassos, & Hazin, 2012). Therefore, a consequence of these fisheries portends greater risks to the vulnerability of large-size snappers.
This study also indicated that there was no restriction on the sizes of juveniles and sub-adults fished from the creek. Mean and modal total lengths, respectively suggests that the fish is prone to growth overfishing such that replenishment by adult stock in the marine habitat declines. It is important to restrict fishing of the species in the creek because they are prominently juveniles.
In the present study, the size and age at first maturity of Lutjanus goreensis or related species in the Eastern central Atlantic are not known. Therefore, the mean total length and the most fished length class of marine samples could be below the maturation size for the species, the consequence of which would be recruitment overfishing. Sub-adults sampled from the marine habitat were particularly susceptible to the designated 44mm cod-end shrimp trawl net which portends a high retention rate of immature and small-sized fish. Major commercial shrimps stocks occur abundantly in the Lagos West fishing grounds, Niger Delta and eastwards to Cross River (Ogbonna, 2001). Exploitation of the penaeid shrimps occur in shallow waters <60m depth over soft muddy substrates of the continental shelf within the coastal waters. Co-habitation of shrimps and fishes suggested a strong predator-prey interaction between sub-adult snappers and the shrimps. Previous studies by Gallaway, Cole, Meyer, and Roscigno (1999); Martinez-Andrade (2003); Andrade-Rodriguez (2003); Amezcua, Soto, and Green (2006); Gonzalez-Ochoa, Lopez-Martinez and Hernandez-Saavedra (2009) reported the association of young snapper species with shrimp ground assemblage, and clearly emphasized on their feeding ecology and habitat preferences. Globally, snappers are reported as major by-catch (that is, they constitute more than 5% of total catch) of the industrial shrimp trawl fishery (Banks, & Macfadyen, 2010). Thus, shrimping in non-trawling zone, use of smaller trawl mesh sizes, boom in by-catch trade and unrestricted fishing of immature fish particularly by coastal artisanal shrimp beam trawl fisheries (Ambrose, 2004; Banks, & Macfadyen, 2010; Nwosu, Ita, & Enin, 2011) would merely escalate the incidence of sub-adult snappers as by-catch.
Comparison between length-frequency distributions of L. goreensis also suggests that an increase in mean total lengths from the creek to the marine habitat denotes habitat connectivity through ontogenetic shift in habitat use. The absence of large adults in the creek suggests they probably remain in the marine habitat. In the creek, the sub-adult population consisted of migratory individuals which moved from the creek through the Lagos Harbour to the Atlantic Ocean. Similarly, Sheaves (1995) reported migratory individuals as those in the larger size classes which moved through the lower part of the estuaries during offshore migration. Migration of lutjanid adults offshore was permanent once they left the estuary (Sheaves, 1995).
Two migration patterns are suggested in this study. First, a mono- or uni-directional migration pattern from the creek to the marine habitat in which fish size increases. The strategy employed by the species appears to be that of an ontogenetic shifter where habitat use is dependent on life-stage as described by Jaxion-Harm, Saunders and Speight (2012). Gonad development and dietary shifts are known theoretically to initiate ontogenetic migrations in fishes (Gillanders, Able, Brown, Eggleston, & Sheridan, 2003; Moura et al., 2011; Jaxion-Harm et al., 2012). In the present study, the most important fish feeding areas usually lie within the shallowest areas of continental shelf, which constitute the most productive fishing grounds along the Nigerian coastline. Therefore, it is possible that expansion of dietary needs act as a trigger to migration of sub-adult L. goreensis from the creek to the marine habitat where may be an abundance of highly diverse spectrum of preferred preys exist. Similarly, Cocheret de la Moriniere et al. (2003); Case, Westneat, and Marshall (2008); Jaxion-Harm et al. (2012) had expressed the larger role of diet as influencing migration pattern in some lutjanids. The probability of gonad maturation as a contributing factor in initiating migration to the marine habitat cannot be overruled. This is more obvious judging from lack of sexually mature individuals in the creek. Some researchers had postulated sexual maturation of sub-adult lutjanids in the shallow marine areas as a key factor before migration to deeper waters (Cocheret de la Moriniere et al., 2002; Aburto-Oropeva et al., 2009).
Predominance of sub-adults in marine samples lends anecdotal evidence that sub-adult staging areas for the species were located within the larger marine habitat. This is consistent with the views of Gillanders et al. (2003). From the foregoing, ecological conditions required for sexual maturation of L. goreensis were obviously absent in the creek. However, diet shift could be more significant when its influence on sexual maturation is considered. The physiology of sexual maturation in some lutjanids is enhanced by the consumption of more fish than crustaceans because of higher energy content (Tripp-Valdez, & Arreguin-Sanchez, 2009). Therefore, the interaction of these two key factors could account for differences in habitat resources between the juvenile and adult habitats and thus necessitate habitat shift in the species.
Secondly, the probability of a cross- or bi-directional migration between the creek and the marine habitat may also be inferred from the concurrent size classes of sub-adults in both habitats. Feeding may be a fundamental reason for the marine sub-adult population of the species to return to the creek where there may be preferred small preys. Similarly, Okyere et al. (2011); Aheto et al. (2011) suggested occasional migration of adult marine fish species (interpreted here to include sub-adults forms) to estuaries where they once lived as juveniles for the purpose of feeding. Feeding migration has also been reported in immature L. griseus and L. analis (Claro et al., 2009). Adult-sized L. griseus and mullids also migrate daily into seagrass/mangrove habitats which serve as juvenile habitats for feeding (Honda et al., 2013). Thus, it may be assumed that based on the observations of sub-adults in both habitats, the species is a habitat generalist.
Occurrence of a predominant juvenile population in the creek is indicative of a juvenile habitat. The postulated life cycle in medium to large species in subfamily Lutjaninae (Martinez- Andrade, 2003) and movement of snapper larvae in Barrier Lagoon complex and Strand Coasts (Ezenwa et al., 1990), both insinuate migration of early juveniles to the estuarine environment from the coastal waters where they occurred as larvae, while spawning most apparently must have occurred in the marine habitat. Data from the present study supports this. From the smaller size classes (<10cm) in the creek relative to those of the coastal waters, the choice of nursery habitat for the species is presumed to occur in estuarine environments. Therefore, the paucity of juvenile L. goreensis in marine samples strongly suggests that coastal waters are clearly not a nursery ground for the species.
In addition, the paucity of juveniles in the marine habitat clearly demonstrates that juvenile phase of L. goreensis in the creek has a clear dependency on the estuarine environment for its survival. The species spends only part of its life cycle in the creek. Martinez-Andrade (2003) also considered L. goreensis as an estuarine-dependent species for its juvenile phase; Aheto et al. (2011); Okyere et al. (2011); Brochier, Ecoutin, de Morais, Kaplan, and Lae (2013) considered L. goreensis as a marine species accessory in estuaries implying that it resides in estuaries from the juvenile phase but reproduces only in the sea as adults.
The ability to utilise multiple estuaries as putative juvenile habitats and later to reassemble as adults in larger groups in the marine habitat is suggestive of the evolution of meta-population structures, a trait of marine fish species which exhibit estuarine dependency (Vasconcelos, 2009). Both tidal creeks and mangroves are habitats of juvenile snappers (Harbone et al., 2006). Besides, a predominant juvenile L. goreensis occurring in the mangrove-depauperate tidal creek such as Five Cowrie Creek, Omogoriola et al. (2012) had reported their occurrence in mangroves of the Eastern parts of Lagos Lagoon.
A mangrove-poor habitat has a negative impact on estuarine-dependent taxa and productivity of fisheries (Able, 2005). The present study aligns with this view on the basis that a mangrove-poor environment such as Five Cowrie Creek portends loss of suitable and productive feeding grounds necessary for the survival of the species’ juvenile phase, particularly if it is an obligatory mangrove user. Several studies including those of Sedberry and Carter (1993); Aburto-Oropeza et al. (2009); Monteiro, Giarrizzo, and Isaac (2009); Conboy and Haynes (2011) have highlighted the importance of mangroves as essential nursery habitats for juveniles of many lutjanid species. However if the juvenile phase of species is not an obligatory user of mangroves as a juvenile habitat, then it may be implied to be a facultative user of the mangrove-poor creek as a juvenile habitat. This also suggests that the species reflects flexibility in habitat use, because it is inferred that mangroves could be facultative habitats for the species.
Otherwise, by virtue of being one of the two routes channelling water from the Lagos Lagoon Complex into the Atlantic Ocean via the Lagos Harbour, Five Cowrie Creek is suggested to serve merely as a migratory corridor connecting the Lagos Lagoon, a larger juvenile/ nursery habitat to the Atlantic Ocean, a marine habitat for many economically important estuarine-dependent fishes.
In conclusion, the species is more of an ontogenetic shifter than a habitat generalist. Estuarine dependency and habitat flexibility are characteristics of the juvenile phase of the species. Indiscriminate fishing of the species from the creek could result in growth overfishing. Therefore, the length-frequency distribution of L. goreensis underscores the importance of preserving estuarine environments as essential fish habitats for juveniles of commercially important fish species. Subtle evidence suggests that large reproductive adults may be more vulnerable to exploitation during the rainy season while biodiversity loss inferred could be from the paucity of snappers exceeding 50cm in commercial catches. In addition, there is also a tendency for recruitment overfishing from shrimping when fish modal length is less than maturation size for the species. Thus, to prevent further over-exploitation of reproductive adults and sexually immature L. goreensis in the marine environment, there is need to identify spawning period of the species and shrimp ground assemblage for proper delineation of closed areas and time to fishing while placing a full ban on fishing of the species from estuarine environments.
Acknowledgments
We thank the Chief technologist, Richard Ajepe and his entire staff for laboratory facilities and assistance with collection of morphometric data of fish specimens.
References
Able, K. W. (2005). An examination of fish estuarine dependence: evidence for connectivity between estuarine and ocean habitats. Estuarine, Coastal and Shelf Science, 64, 5-17. [ Links ]
Aburto-Oropeza, O., Dominguez-Guerrero, I., Cota-Nieto, J., & Plomozo-Lugo, T. (2009). Recruitment and ontogenetic habitat shifts of the Yellow snapper (Lutjanus argentiventris) in the Gulf of California. Marine Biology, 156, 2461-2472. [ Links ]
Adekanmbi, O. H., & Ogundipe, O. (2009). Mangrove biodiversity in the restoration and sustainability of the Nigerian natural environment. Journal of Ecology and Natural Environment, 1(3), 064-072. [ Links ]
Agboola, J. I., & Anatekhai, M. A. (2008). Length weight relationships of some fresh and brackish water fishes in Badagry creek, Nigeria. Journal of Applied Ichthyology, 24, 623-625. [ Links ]
Aheto, D. W., Mensah, E., Aggrey-Fynn, J., Obodai, E. A., Mensah, C. J., Okyere, I., & Aheto, S. P. K. (2011). Spatio-temporal analysis of two coastal wetland systems in Ghana: Addressing ecosystem vulnerability and implications for fisheries development in the context of climate and land use changes. Archives of Applied Science Research, 3(3), 499-513. [ Links ]
Allen, G. R. (1985). FAO Species Catalogue. Snappers of the World. An annotated andillustrated catalogue of Lutjanid species known to date. FAO Fisheries Synopsis, 6, 125. [ Links ]
Ambrose, E. (2004). Seasonal Variations in shrimp, Nematopalaemon hastatus and by-catch compositions from coastal artisanal shrimp beam trawl fisheries off Lagos coast and Applied Zoology & Environmental Biology, 6, 1-5. [ Links ]
Amezcua, F., Soto, C., & Green, Y. (2006) Age, growth, and mortality of the Spotted rose snapper Lutjanus guttatus from the Southeastern Gulf of California. Fisheries Research, 77, 293-300. [ Links ]
Amiengheme, P. (1997). Problems of industrial fisheries in Nigeria - Report on the utilization of Automotive Gas Oil (AGO) bunkered by the Nigerian Trawler Owner Association (NITOA) for that period August 6-31, 1997. In T. O. Ajayi, & B. I. O. Ezenwa (Eds.), Scientists on nationally co-coordinating resource programme (NCRP) of the National Agricultural Research Programmes (NCRP)of the National Agricultural Research Project (NARP), National Meeting Report No. 1 (1998),(pp.145-149). Lagos, Nigeria: Nigerian Institute for Oceanography and Marine Research. [ Links ]
Amiengheme, P. (2001). Nigeria’s fisheries regulations and their enforcement. Seminar on the Nigerian fishing industry, Sheraton Hotels and Towers, Abuja, Nigeria. [ Links ]
Andrade-Rodriguez, H. A. (2003). Age determination in the snapper Lutjanus guttatus (Pisces, Lutjanidae) and investigation of fishery management strategies in the Pacific Coast of Guatemala. M.Sc. Thesis. Norwegian College of Fisheries Science, University of Tromsø, Norway. [ Links ]
Banks, R., & Macfadyen, G. (2010). A blueprint for sustainable tropical shrimp trawl fisheries. Poseidon Aquatic Resource Management Ltd., WWF. [ Links ]
Brochier, T., Ecoutin, J. M., de Morais, L. T., Kaplan, D. M., & Lae, R. (2013). A multi- agent ecosystem model for studying changes in a tropical estuarine fish assemblage within a marine protected area. Aquatic Living Resources, 26, 147-158. [ Links ]
Case, J. E., Westneat, M. W., & Marshall, C. D. (2008). Feeding biomechanics of juvenile red snapper (Lutjanus campechanus) from the North-Western Gulf of Mexico. The Journal of Experimental Biology, 211, 3826-3835. [ Links ]
Claro, R., Sadovy de Mitcheson, Y., Lindeman, K. C., & García-Cagide, R. A. (2009). Historical analysis of Cuban commercial fishing effort and the effects of management interventions on important reef fishes from 1960–2005. Fisheries Research, 99, 7-16. [ Links ]
Cocheret de la Morinie` re, E., Pollux, B. J. A., Nagelkerken, I., & van der Velde, G. (2002). Post-settlement life cycle migration patterns in relation to biotope preference of coral reef fish that use seagrass and mangrove habitats as nurseries. Estuarine, Coastal and Shelf Science, 55, 309-321. [ Links ]
Cocheret de la Morinie`re, E., Pollux, B. J. A., Nagelkerken, I., & van der Velde, G. (2003). Diet shifts of Caribbean grunts (Haemulidae) and snappers (Lutjanidae) and the relation with nursery-to-coral reef Migrations. Estuarine, Coastal and Shelf Science, 57, 1079-1089. [ Links ]
Conboy, I. C., & Haynes, J. M. (2011). Potential of Pigeon Creek, San Salvador, Bahamas, as nursery habitat for juvenile reef fish. The International Journal of Bahamian Studies, 17(2), 9-26. [ Links ]
Ediang, O. A., & Ediang, A. A. (2013). Beyond data regulation: finding a solution to the persistent problem of marine debris and sea surface temperature measurement along the coastline of Lagos, Nigeria. Data Science Journal, 12, 129-133. [ Links ]
Ezenwa, B. I. O., Alegbeleye, W. O., Anyanwu, P. E., & Uzukwu, P. U. (1990). Cultivable fish seeds in Nigerian coastal waters: A research survey (Second Phase: 1986-1989). Technical Paper, Nigerian Institute for Oceanography and Marine Research, 66, 1-37. [ Links ]
Fakoya, K. A., Abass, M. A., Owodeinde, F. G., Lawson, E. O., & Ojo, F. R. (2010). A preliminary study of diet in the juvenile Gorean snapper, Lutjanus goreensis (Valenciennes, 1830), pp. 232-236. In H. A. Fashina-Bombata, E. J. Ansa, & P. E. Ndimele (Eds.), Proceedings of the 25th Annual FISON International Conference and Exhibition, Topo-Badagry, Lagos, Nigeria. October 25-29, 2010. (CD - ROM). [ Links ]
Faye, D., Le Loc’h, F., Thiaw, O. T., & Tito de Moraïs, L. (2012). Mechanisms of food partitioning and ecomorphological correlates in ten fish species from a tropical estuarine marine protected area (Bamboung, Senegal, West Africa). African Journal of Agricultural Research, 7(3), 443-455. [ Links ]
Fernandes, C. A. F., de Oliveira, P. G. V., Travassos, P. E. P., & Hazin, F. H. V. (2012). Reproduction of the Brazilian snapper, Lutjanus alexandrei Moura & Lindeman, 2007 (Perciformes: Lutjanidae), off the Northern coast of Pernambuco, Brazil. Neotropical Ichthyology, 10(3), 587-592. [ Links ]
Fischer, W., Bianchi, G., & Scott, W. B. (Eds.). (1981). FAO species identification sheets for fishery purposes. Eastern Central Atlantic; fishing areas 34, 47 (in part). Ottawa, Canada: Canada Funds-in-Trust, Department of Fisheries and Oceans and the Food and Agriculture Organization of the United Nations. [ Links ]
Gallaway, B. J., Cole, J. G., Meyer, R., & Roscigno, P. (1999). Delineation of essential habitat for juvenile red snapper in the Northwestern Gulf of Mexico. Transactions of the American Fisheries Society, 128(4), 713-726. [ Links ]
García-Contreras, O. E., Quiñónez-Velázquez, C., Morán-Angulo, R. E., & Valdez-Pineda, M. C. (2009). Age, growth, and age- structure of Amarillo snapper off the coast of Mazatlán, Sinaloa, Mexico. North American Journal of Fisheries Management, 29(1), 223-230. [ Links ]
Gillanders, B. M., Able, K. W., Brown, J. A., Eggleston, D. B., & Sheridan, P. F. (2003). Evidence of connectivity between juvenile and adult habitats for mobile marine fauna: an important component of nurseries. Marine Ecology Progress Series, 247, 281-295. [ Links ]
Gonzalez-Ochoa, O. A., Lopez-Martinez, J., & Hernandez-Saavedra, N. Y. (2009). Population characteristics of Spotted rose snapper Lutjanus guttatus caught as shrimp by catch in the Gulf of California. Interciencia, 34 (11), 808-813. [ Links ]
Grimes, C. B. (1987). Reproductive biology of the Lutjanidae: a review. In J. J. Polovina, & S. Ralston (Eds.), Tropical Snappers and Groupers: Biology and Fisheries Management (p.p. 239-294). Ocean Resources. Marine Policy Series. Boulder, Colorado, USA: Westview Press. [ Links ]
Grandcourt, E. M., Abdessalaam, T. Z., & Franklin, F. (2006). Age, growth, mortality and reproduction of the Blackspot snapper, Lutjanus fulviflamma (Forsskal, 1773) in the Southern Arabian Gulf. Fisheries Research, 71(2-3), 203-210. [ Links ]
Hammerschlag-Peyer, C. M., Allgeier, J. E., & Layman, C. A. (2013). Predator effects on faunal community composition in shallow seagrass beds of the Bahamas. Journal of Experimental Marine Biology and Ecology, 446, 282-290. [ Links ]
Harborne, A. R., Mumby, P. J., Micheli, F., Christopher, T., Perry, C. T., Dahlgren, C. P., Holmes, K. E., & Brumbaugh, D. R. (2006).The functional value of Caribbean coral reef, seagrass and mangrove habitats to ecosystem processes. Advances in Marine Biology, 50, 57-189. [ Links ]
Heery, E. C. (2007). The impact of bias in length frequency data on an age-structured fisheries stock assessment model (M.Sc. Thesis). Virginia Polytechnic Institute and State University, Virginia, USA. [ Links ]
Honda, K., Nakamura, Y., Nakaoka, M., Uy, W. H., & Fortes, M. D. (2013) Habitat use by fishes in coral reefs, seagrass beds and mangrove habitats in the Philippines. PLoS ONE 8(8), doi:10.1371/journal.pone.0065735. [ Links ]
Jaxion-Harm, J., Saunders, J., & Speight, M. R. (2012). Distribution of fish in seagrass, mangroves and coral reefs: life-stage dependent habitat use in Honduras. Revista de Biología Tropical, 60(2), 683-698. [ Links ]
Johnson, M. G., & Tamatamah, A. R. (2013). Length frequency distribution, mortality rate and reproductive biology of Kawakawa (Euthynnus affinis -Cantour, 1849) in the coastal waters of Tanzania. Pakistan Journal of Biological Sciences, 16, 1270-1278. [ Links ]
Kell, L. T., Bonhommeau, S., & Fromentin, J. M. (2013). Length- based catch analysis for East Atlantic Mediterranean Bluefin Tuna. Collect. Vol. Sci. Pap. ICCAT, 69, 204-209. [ Links ]
Lawson, E. O. (2005). A survey of fish fauna and fisheries of Badagry Creek. Journal of Agriculture and Environmental Research Studies, 1(1): 87-99. [ Links ]
Longhurst, A. R. (1969). Species assemblages in tropical demersal fisheries. Symposium on the Oceanography and Fisheries Resources of the UNESCO/FAO/OAU Tropical Atlantic. Results of ICITA and GTS, 20-28 October 1966, Abidjan, Ivory Coast (p.p. 147-168). UNESCO, Paris, France. [ Links ]
Mamonekene, V., Lavoué, S., Olivier, S. G., Pauwels, O. S. G., MveBeh, J. H., Mackayah, J. E., & Tchignoumba, L. (2006). Fish diversity at Rabi and Gamba, Ogooué- Maritime Province, Gabon. Bulletin of the Biological Society of Washington, 12, 285-296. [ Links ]
Manjarrés-Martínez, L. M., Gutiérrez-Estrada, J. C., Mazenet-González, J., & Soriguer, M. C. (2010). Seasonal patterns of three fish species in a Caribbean coastal gill-net fishery: Biologically induced or climate-related aggregations? Fisheries Research, 106, 358-367. [ Links ]
Martinez-Andrade, F. (2003). A comparison of life histories and ecological aspects among snappers (Pisces: Lutjanidae) (Ph.D. Thesis). Louisiana State University, Baton Rouge, LA, USA. [ Links ]
Monteiro, D. P., Giarrizzo, T., & Isaac, V. (2009): Feeding ecology of juvenile Dog snapper Lutjanus jocu (Bloch and Shneider, 1801) (Lutjanidae) in Intertidal Mangrove Creeks in Curuçá Estuary (Northern Brazil). Brazilian Archives of Biology and Technology, 52(6), 1421-1430. [ Links ]
Moura, R. L., Francini-Filho, R. B., Chaves, E. M., Minte-Vera, C. V., & Lindeman, K. C. (2011). Use of riverine through reef habitat systems by Dog snapper (Lutjanus jocu) in eastern Brazil. Estuarine, Coastal and Shelf Science, 95, 274-278. [ Links ]
Nagelkerken, I., & van der Velde, G. (2002). Do non-estuarine mangroves harbour higher densities of juvenile fish than adjacent shallow-water and coral reef habitats in Curaçao (Netherlands Antilles)? Marine Ecology Progress Series, 245, 191-204. [ Links ]
Newman, S. J. (1995). Spatial variability in the distribution, abundance, growth, mortality and age structures of tropical snappers (Pisces: Lutjanidae) in the Central Great Barrier Reef, Australia (Ph.D. Thesis). James Cook University, Queensland, Australia. [ Links ]
Nwankwo, D. I, Okedoyin, J. O., & Adesalu, T. A. (2012). Primary Productivity in Tidal Creeks of South-West Nigeria II. Comparative Study of Nutrient Status and Chlorophyll-a variations in two Lagos Harbour creeks. World Journal of Biological Research, 5(1), 41-48. [ Links ]
Nwosu, F. M., Ita, E. O., & Enin, U. I. (2011). Fisheries management in Nigeria: A case study of the marine fisheries policy. International Research Journal of Agricultural Science and Soil Science, 1(3), 070-076. [ Links ]
Ogbonna, J. C. (2001). Reducing the impact of tropical shrimp trawling fisheries on the living marine resources through the adoption of environmentally friendly techniques and practices in Nigeria. In FAO - Food and Agriculture Organization. Tropical shrimp fisheries and their impact on living resources. Shrimp fisheries in Asia: Bangladesh, Indonesia and the Philippines; in the Near East: Bahrain and Iran; in Africa: Cameroon, Nigeria and the United Republic of Tanzania; in Latin America: Colombia, Costa Rica, Cuba, Trinidad and Tobago, and Venezuela (pp. 188-215). Rome: FAO Circular No. 974. [ Links ]
Okyere, I., Aheto, D. W., & Aggrey-Fynn, J. (2011). Comparative ecological assessment of biodiversity of fish communities in three coastal wetland systems in Ghana. European Journal of Experimental Biology, 1(2), 178-188. [ Links ]
Omogoriola, H. O., Williams, A. B., Ukaonu, S. C., Adegbile, O. M., Olakolu, F. C., Mbawuike, B. C., Akinnigbagbe, A. E., & Ajulo, A. A. (2012). Survey, biodiversity and impacts of economic activities on mangroves ecosystem in Eastern part of Lagos Lagoon, Nigeria. Nature and Science, 10(10), 30-34. [ Links ]
Onyema, I. C., Nwankwo, D. I., & Oduleye, T. (2005/2006). Diatoms and dinoflagellates of an estuarine creek in Lagos. Journal of Scientific Research and Development, 10, 73-82’ [ Links ].
Oribhabor, B. J., & Ogbeibu, A. E. (2010). The ecological impact of anthropogenic activities on the predatory fish assemblage of a tidal creek in the Niger Delta. Research Journal of Environmental Sciences, 4(3), 271-279. [ Links ]
Oribhabor, B. J., & Ogbeibu, A. E. (2012). The food and feeding habits of fish species assemblage in a Niger Delta Mangrove creek, Nigeria. Journal of Fisheries and Aquatic Science, 7(2), 134-149. [ Links ]
Reddy, S. M. W., Wentz, A., Aburto-Oropeza, O., Maxey, M., Nagavarapu, S., Heather, M., & Leslie, H. M. (2013). Evidence of market-driven size-selective fishing and the mediating effects of biological and institutional factors. Ecological Applications, 23(4), 726-741. [ Links ]
Riley, K. L. P. (2002). Refrigerated storage and cryopreservation of sperm for the production of Red snapper and snapper hybrids (M.Sc. Thesis). Louisiana State University, Louisiana, USA. [ Links ]
Sedberry, G. R., & Carter, J. (1993). Fish community of a shallow tropical lagoon in Belize. Estuaries, 16(2), 198-215. [ Links ]
Sheaves, M. J. (1995). Estuaries as juvenile habitats for lutjanid and serranid fishes in tropical Northeastern Australia (Ph.D. Thesis). James Cook University, Australia. [ Links ]
Ssentongo, G. W., Ukpe, E. T., & Ajayi, T. O. (1986). Marine fishery resources of Nigeria: a review of exploited fish stocks. CECAF/ECAF SERIES 86/40. [ Links ]
Thys van den Audenaerde, D. (1966). List of freshwater fishes presently known from the Island of Fernando Poo. Bonner Zoologische Beiträge, 16, 316-317. [ Links ]
Tobor, J. G. (1991). Marine fish resources of West Africa: Potentials, management, development and constraints to their utilization to satisfying increasing demand. Technical Paper, Nigerian Institute for Oceanography and Marine Research, 70, 1-25. [ Links ]
Tripp-Valdez, A., & Arreguin-Sanchez, F. (2009). The use of stable isotopes and stomach contents to identify dietary components of Spotted rose snapper, Lutjanus guttatus (Steindachner, 1869), off Eastern coast of the Southern Gulf of Carlifornia. Journal of Fisheries and Aquatic Science, 4, 274-284. [ Links ]
Vasconcelos, R. M. P. (2009). Contribution of estuarine nursery areas to the structure and biocontamination of stocks of commercially important fish species along the Portuguese coast (Ph.D. dissertation). University of Lisboa, Portugal. [ Links ]
Aburto-Oropeza, O., Dominguez-Guerrero, I., Cota-Nieto, J., & Plomozo-Lugo, T. (2009). Recruitment and ontogenetic habitat shifts of the Yellow snapper (Lutjanus argentiventris) in the Gulf of California. Marine Biology, 156, 2461-2472. [ Links ]
Adekanmbi, O. H., & Ogundipe, O. (2009). Mangrove biodiversity in the restoration and sustainability of the Nigerian natural environment. Journal of Ecology and Natural Environment, 1(3), 064-072. [ Links ]
Agboola, J. I., & Anatekhai, M. A. (2008). Length weight relationships of some fresh and brackish water fishes in Badagry creek, Nigeria. Journal of Applied Ichthyology, 24, 623-625. [ Links ]
Aheto, D. W., Mensah, E., Aggrey-Fynn, J., Obodai, E. A., Mensah, C. J., Okyere, I., & Aheto, S. P. K. (2011). Spatio-temporal analysis of two coastal wetland systems in Ghana: Addressing ecosystem vulnerability and implications for fisheries development in the context of climate and land use changes. Archives of Applied Science Research, 3(3), 499-513. [ Links ]
Allen, G. R. (1985). FAO Species Catalogue. Snappers of the World. An annotated andillustrated catalogue of Lutjanid species known to date. FAO Fisheries Synopsis, 6, 125. [ Links ]
Ambrose, E. (2004). Seasonal Variations in shrimp, Nematopalaemon hastatus and by-catch compositions from coastal artisanal shrimp beam trawl fisheries off Lagos coast and Applied Zoology & Environmental Biology, 6, 1-5. [ Links ]
Amezcua, F., Soto, C., & Green, Y. (2006) Age, growth, and mortality of the Spotted rose snapper Lutjanus guttatus from the Southeastern Gulf of California. Fisheries Research, 77, 293-300. [ Links ]
Amiengheme, P. (1997). Problems of industrial fisheries in Nigeria - Report on the utilization of Automotive Gas Oil (AGO) bunkered by the Nigerian Trawler Owner Association (NITOA) for that period August 6-31, 1997. In T. O. Ajayi, & B. I. O. Ezenwa (Eds.), Scientists on nationally co-coordinating resource programme (NCRP) of the National Agricultural Research Programmes (NCRP)of the National Agricultural Research Project (NARP), National Meeting Report No. 1 (1998),(pp.145-149). Lagos, Nigeria: Nigerian Institute for Oceanography and Marine Research. [ Links ]
Amiengheme, P. (2001). Nigeria’s fisheries regulations and their enforcement. Seminar on the Nigerian fishing industry, Sheraton Hotels and Towers, Abuja, Nigeria. [ Links ]
Andrade-Rodriguez, H. A. (2003). Age determination in the snapper Lutjanus guttatus (Pisces, Lutjanidae) and investigation of fishery management strategies in the Pacific Coast of Guatemala. M.Sc. Thesis. Norwegian College of Fisheries Science, University of Tromsø, Norway. [ Links ]
Banks, R., & Macfadyen, G. (2010). A blueprint for sustainable tropical shrimp trawl fisheries. Poseidon Aquatic Resource Management Ltd., WWF. [ Links ]
Brochier, T., Ecoutin, J. M., de Morais, L. T., Kaplan, D. M., & Lae, R. (2013). A multi- agent ecosystem model for studying changes in a tropical estuarine fish assemblage within a marine protected area. Aquatic Living Resources, 26, 147-158. [ Links ]
Case, J. E., Westneat, M. W., & Marshall, C. D. (2008). Feeding biomechanics of juvenile red snapper (Lutjanus campechanus) from the North-Western Gulf of Mexico. The Journal of Experimental Biology, 211, 3826-3835. [ Links ]
Claro, R., Sadovy de Mitcheson, Y., Lindeman, K. C., & García-Cagide, R. A. (2009). Historical analysis of Cuban commercial fishing effort and the effects of management interventions on important reef fishes from 1960–2005. Fisheries Research, 99, 7-16. [ Links ]
Cocheret de la Morinie` re, E., Pollux, B. J. A., Nagelkerken, I., & van der Velde, G. (2002). Post-settlement life cycle migration patterns in relation to biotope preference of coral reef fish that use seagrass and mangrove habitats as nurseries. Estuarine, Coastal and Shelf Science, 55, 309-321. [ Links ]
Cocheret de la Morinie`re, E., Pollux, B. J. A., Nagelkerken, I., & van der Velde, G. (2003). Diet shifts of Caribbean grunts (Haemulidae) and snappers (Lutjanidae) and the relation with nursery-to-coral reef Migrations. Estuarine, Coastal and Shelf Science, 57, 1079-1089. [ Links ]
Conboy, I. C., & Haynes, J. M. (2011). Potential of Pigeon Creek, San Salvador, Bahamas, as nursery habitat for juvenile reef fish. The International Journal of Bahamian Studies, 17(2), 9-26. [ Links ]
Ediang, O. A., & Ediang, A. A. (2013). Beyond data regulation: finding a solution to the persistent problem of marine debris and sea surface temperature measurement along the coastline of Lagos, Nigeria. Data Science Journal, 12, 129-133. [ Links ]
Ezenwa, B. I. O., Alegbeleye, W. O., Anyanwu, P. E., & Uzukwu, P. U. (1990). Cultivable fish seeds in Nigerian coastal waters: A research survey (Second Phase: 1986-1989). Technical Paper, Nigerian Institute for Oceanography and Marine Research, 66, 1-37. [ Links ]
Fakoya, K. A., Abass, M. A., Owodeinde, F. G., Lawson, E. O., & Ojo, F. R. (2010). A preliminary study of diet in the juvenile Gorean snapper, Lutjanus goreensis (Valenciennes, 1830), pp. 232-236. In H. A. Fashina-Bombata, E. J. Ansa, & P. E. Ndimele (Eds.), Proceedings of the 25th Annual FISON International Conference and Exhibition, Topo-Badagry, Lagos, Nigeria. October 25-29, 2010. (CD - ROM). [ Links ]
Faye, D., Le Loc’h, F., Thiaw, O. T., & Tito de Moraïs, L. (2012). Mechanisms of food partitioning and ecomorphological correlates in ten fish species from a tropical estuarine marine protected area (Bamboung, Senegal, West Africa). African Journal of Agricultural Research, 7(3), 443-455. [ Links ]
Fernandes, C. A. F., de Oliveira, P. G. V., Travassos, P. E. P., & Hazin, F. H. V. (2012). Reproduction of the Brazilian snapper, Lutjanus alexandrei Moura & Lindeman, 2007 (Perciformes: Lutjanidae), off the Northern coast of Pernambuco, Brazil. Neotropical Ichthyology, 10(3), 587-592. [ Links ]
Fischer, W., Bianchi, G., & Scott, W. B. (Eds.). (1981). FAO species identification sheets for fishery purposes. Eastern Central Atlantic; fishing areas 34, 47 (in part). Ottawa, Canada: Canada Funds-in-Trust, Department of Fisheries and Oceans and the Food and Agriculture Organization of the United Nations. [ Links ]
Gallaway, B. J., Cole, J. G., Meyer, R., & Roscigno, P. (1999). Delineation of essential habitat for juvenile red snapper in the Northwestern Gulf of Mexico. Transactions of the American Fisheries Society, 128(4), 713-726. [ Links ]
García-Contreras, O. E., Quiñónez-Velázquez, C., Morán-Angulo, R. E., & Valdez-Pineda, M. C. (2009). Age, growth, and age- structure of Amarillo snapper off the coast of Mazatlán, Sinaloa, Mexico. North American Journal of Fisheries Management, 29(1), 223-230. [ Links ]
Gillanders, B. M., Able, K. W., Brown, J. A., Eggleston, D. B., & Sheridan, P. F. (2003). Evidence of connectivity between juvenile and adult habitats for mobile marine fauna: an important component of nurseries. Marine Ecology Progress Series, 247, 281-295. [ Links ]
Gonzalez-Ochoa, O. A., Lopez-Martinez, J., & Hernandez-Saavedra, N. Y. (2009). Population characteristics of Spotted rose snapper Lutjanus guttatus caught as shrimp by catch in the Gulf of California. Interciencia, 34 (11), 808-813. [ Links ]
Grimes, C. B. (1987). Reproductive biology of the Lutjanidae: a review. In J. J. Polovina, & S. Ralston (Eds.), Tropical Snappers and Groupers: Biology and Fisheries Management (p.p. 239-294). Ocean Resources. Marine Policy Series. Boulder, Colorado, USA: Westview Press. [ Links ]
Grandcourt, E. M., Abdessalaam, T. Z., & Franklin, F. (2006). Age, growth, mortality and reproduction of the Blackspot snapper, Lutjanus fulviflamma (Forsskal, 1773) in the Southern Arabian Gulf. Fisheries Research, 71(2-3), 203-210. [ Links ]
Hammerschlag-Peyer, C. M., Allgeier, J. E., & Layman, C. A. (2013). Predator effects on faunal community composition in shallow seagrass beds of the Bahamas. Journal of Experimental Marine Biology and Ecology, 446, 282-290. [ Links ]
Harborne, A. R., Mumby, P. J., Micheli, F., Christopher, T., Perry, C. T., Dahlgren, C. P., Holmes, K. E., & Brumbaugh, D. R. (2006).The functional value of Caribbean coral reef, seagrass and mangrove habitats to ecosystem processes. Advances in Marine Biology, 50, 57-189. [ Links ]
Heery, E. C. (2007). The impact of bias in length frequency data on an age-structured fisheries stock assessment model (M.Sc. Thesis). Virginia Polytechnic Institute and State University, Virginia, USA. [ Links ]
Honda, K., Nakamura, Y., Nakaoka, M., Uy, W. H., & Fortes, M. D. (2013) Habitat use by fishes in coral reefs, seagrass beds and mangrove habitats in the Philippines. PLoS ONE 8(8), doi:10.1371/journal.pone.0065735. [ Links ]
Jaxion-Harm, J., Saunders, J., & Speight, M. R. (2012). Distribution of fish in seagrass, mangroves and coral reefs: life-stage dependent habitat use in Honduras. Revista de Biología Tropical, 60(2), 683-698. [ Links ]
Johnson, M. G., & Tamatamah, A. R. (2013). Length frequency distribution, mortality rate and reproductive biology of Kawakawa (Euthynnus affinis -Cantour, 1849) in the coastal waters of Tanzania. Pakistan Journal of Biological Sciences, 16, 1270-1278. [ Links ]
Kell, L. T., Bonhommeau, S., & Fromentin, J. M. (2013). Length- based catch analysis for East Atlantic Mediterranean Bluefin Tuna. Collect. Vol. Sci. Pap. ICCAT, 69, 204-209. [ Links ]
Lawson, E. O. (2005). A survey of fish fauna and fisheries of Badagry Creek. Journal of Agriculture and Environmental Research Studies, 1(1): 87-99. [ Links ]
Longhurst, A. R. (1969). Species assemblages in tropical demersal fisheries. Symposium on the Oceanography and Fisheries Resources of the UNESCO/FAO/OAU Tropical Atlantic. Results of ICITA and GTS, 20-28 October 1966, Abidjan, Ivory Coast (p.p. 147-168). UNESCO, Paris, France. [ Links ]
Mamonekene, V., Lavoué, S., Olivier, S. G., Pauwels, O. S. G., MveBeh, J. H., Mackayah, J. E., & Tchignoumba, L. (2006). Fish diversity at Rabi and Gamba, Ogooué- Maritime Province, Gabon. Bulletin of the Biological Society of Washington, 12, 285-296. [ Links ]
Manjarrés-Martínez, L. M., Gutiérrez-Estrada, J. C., Mazenet-González, J., & Soriguer, M. C. (2010). Seasonal patterns of three fish species in a Caribbean coastal gill-net fishery: Biologically induced or climate-related aggregations? Fisheries Research, 106, 358-367. [ Links ]
Martinez-Andrade, F. (2003). A comparison of life histories and ecological aspects among snappers (Pisces: Lutjanidae) (Ph.D. Thesis). Louisiana State University, Baton Rouge, LA, USA. [ Links ]
Monteiro, D. P., Giarrizzo, T., & Isaac, V. (2009): Feeding ecology of juvenile Dog snapper Lutjanus jocu (Bloch and Shneider, 1801) (Lutjanidae) in Intertidal Mangrove Creeks in Curuçá Estuary (Northern Brazil). Brazilian Archives of Biology and Technology, 52(6), 1421-1430. [ Links ]
Moura, R. L., Francini-Filho, R. B., Chaves, E. M., Minte-Vera, C. V., & Lindeman, K. C. (2011). Use of riverine through reef habitat systems by Dog snapper (Lutjanus jocu) in eastern Brazil. Estuarine, Coastal and Shelf Science, 95, 274-278. [ Links ]
Nagelkerken, I., & van der Velde, G. (2002). Do non-estuarine mangroves harbour higher densities of juvenile fish than adjacent shallow-water and coral reef habitats in Curaçao (Netherlands Antilles)? Marine Ecology Progress Series, 245, 191-204. [ Links ]
Newman, S. J. (1995). Spatial variability in the distribution, abundance, growth, mortality and age structures of tropical snappers (Pisces: Lutjanidae) in the Central Great Barrier Reef, Australia (Ph.D. Thesis). James Cook University, Queensland, Australia. [ Links ]
Nwankwo, D. I, Okedoyin, J. O., & Adesalu, T. A. (2012). Primary Productivity in Tidal Creeks of South-West Nigeria II. Comparative Study of Nutrient Status and Chlorophyll-a variations in two Lagos Harbour creeks. World Journal of Biological Research, 5(1), 41-48. [ Links ]
Nwosu, F. M., Ita, E. O., & Enin, U. I. (2011). Fisheries management in Nigeria: A case study of the marine fisheries policy. International Research Journal of Agricultural Science and Soil Science, 1(3), 070-076. [ Links ]
Ogbonna, J. C. (2001). Reducing the impact of tropical shrimp trawling fisheries on the living marine resources through the adoption of environmentally friendly techniques and practices in Nigeria. In FAO - Food and Agriculture Organization. Tropical shrimp fisheries and their impact on living resources. Shrimp fisheries in Asia: Bangladesh, Indonesia and the Philippines; in the Near East: Bahrain and Iran; in Africa: Cameroon, Nigeria and the United Republic of Tanzania; in Latin America: Colombia, Costa Rica, Cuba, Trinidad and Tobago, and Venezuela (pp. 188-215). Rome: FAO Circular No. 974. [ Links ]
Okyere, I., Aheto, D. W., & Aggrey-Fynn, J. (2011). Comparative ecological assessment of biodiversity of fish communities in three coastal wetland systems in Ghana. European Journal of Experimental Biology, 1(2), 178-188. [ Links ]
Omogoriola, H. O., Williams, A. B., Ukaonu, S. C., Adegbile, O. M., Olakolu, F. C., Mbawuike, B. C., Akinnigbagbe, A. E., & Ajulo, A. A. (2012). Survey, biodiversity and impacts of economic activities on mangroves ecosystem in Eastern part of Lagos Lagoon, Nigeria. Nature and Science, 10(10), 30-34. [ Links ]
Onyema, I. C., Nwankwo, D. I., & Oduleye, T. (2005/2006). Diatoms and dinoflagellates of an estuarine creek in Lagos. Journal of Scientific Research and Development, 10, 73-82’ [ Links ].
Oribhabor, B. J., & Ogbeibu, A. E. (2010). The ecological impact of anthropogenic activities on the predatory fish assemblage of a tidal creek in the Niger Delta. Research Journal of Environmental Sciences, 4(3), 271-279. [ Links ]
Oribhabor, B. J., & Ogbeibu, A. E. (2012). The food and feeding habits of fish species assemblage in a Niger Delta Mangrove creek, Nigeria. Journal of Fisheries and Aquatic Science, 7(2), 134-149. [ Links ]
Reddy, S. M. W., Wentz, A., Aburto-Oropeza, O., Maxey, M., Nagavarapu, S., Heather, M., & Leslie, H. M. (2013). Evidence of market-driven size-selective fishing and the mediating effects of biological and institutional factors. Ecological Applications, 23(4), 726-741. [ Links ]
Riley, K. L. P. (2002). Refrigerated storage and cryopreservation of sperm for the production of Red snapper and snapper hybrids (M.Sc. Thesis). Louisiana State University, Louisiana, USA. [ Links ]
Sedberry, G. R., & Carter, J. (1993). Fish community of a shallow tropical lagoon in Belize. Estuaries, 16(2), 198-215. [ Links ]
Sheaves, M. J. (1995). Estuaries as juvenile habitats for lutjanid and serranid fishes in tropical Northeastern Australia (Ph.D. Thesis). James Cook University, Australia. [ Links ]
Ssentongo, G. W., Ukpe, E. T., & Ajayi, T. O. (1986). Marine fishery resources of Nigeria: a review of exploited fish stocks. CECAF/ECAF SERIES 86/40. [ Links ]
Thys van den Audenaerde, D. (1966). List of freshwater fishes presently known from the Island of Fernando Poo. Bonner Zoologische Beiträge, 16, 316-317. [ Links ]
Tobor, J. G. (1991). Marine fish resources of West Africa: Potentials, management, development and constraints to their utilization to satisfying increasing demand. Technical Paper, Nigerian Institute for Oceanography and Marine Research, 70, 1-25. [ Links ]
Tripp-Valdez, A., & Arreguin-Sanchez, F. (2009). The use of stable isotopes and stomach contents to identify dietary components of Spotted rose snapper, Lutjanus guttatus (Steindachner, 1869), off Eastern coast of the Southern Gulf of Carlifornia. Journal of Fisheries and Aquatic Science, 4, 274-284. [ Links ]
Vasconcelos, R. M. P. (2009). Contribution of estuarine nursery areas to the structure and biocontamination of stocks of commercially important fish species along the Portuguese coast (Ph.D. dissertation). University of Lisboa, Portugal. [ Links ]
1. Department of Fisheries, Faculty of Science, Lagos State University, Ojo, Lagos State Nigeria; kafade@yahoo.com, kafayat.fakoya@lasu.edu.ng, anetekhaimartins@gmail.com, drsilfa@yahoo.com, shehu.akintola@lasu.edu.ng ,sabaola@gmail.com
2. Nigerian Institute for Oceanography and Marine Research, 3 Wilmot Point, Off Bar Beach Road, Victoria Island, Lagos State, Nigeria; mikailabass@yahoo.com
Received 06-V-2014. Corrected 25-IX-2014. Accepted 24-X-2014.


{kind=link}
{kind=link}
{kind=link}