










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 n.1 San José Mar. 2012
Dieta de Odocoileus virginianus (Artiodactyla: Cervidae) en un bosque templado del norte de Oaxaca, México
*Dirección para correspondencia
Abstract
Odocoileus virginianus diet (Artiodactyla: Cervidae) in a temperate forest of Northern Oaxaca, Mexico. The Sierra Madre de Oaxaca region, located in the Northern state of Oaxaca, Mexico, is an area of forest ecosystems subject to high exploitation rates, although in some areas its temperate forests are conserved by indigenous community initiatives that live there. We analyzed the diet of white tailed-deer (Odocoileus virginianus) in the localities of Santa Catarina Lachatao and San Miguel Amatlán from June 1998 to August 1999. Sampling was done during both the wet and dry seasons, and included the observation of browsing traces (238 observations), microhistological analysis of deer feces (28 deer pellet-groups), and two stomach content analysis. The annual diet of white-tailed deer was composed of 42 species from 23 botanical families. The most represented families in the diet of this deer were Fagaceae, Asteraceae, Ericaceae and Fabaceae. There were significant differences in the alpha diversity of the diet during the wet and dry seasons (H’=2.957 and H’=1.832, respectively). The similarity percentage between seasons was 56%. Differences in plant species frequency were significantly higher during the wet season. Herbaceous plants made up the greatest percentage of all the species consumed. The preferred species throughout the year were Senecio sp. (shrub), Sedum dendroideum (herbaceous), Arctostaphylos pungens (shrub) and Satureja macrostema (shrub). Diet species richness was found to be lower than that observed in a tropical forest (Venezuela), tropical dry forest (Mexico) and temperate deciduous and mixed forest (Mexico), but similar to the diet species richness observed in a tropical dry forest (Costa Rica) and temperate coniferous and deciduous forests (USA).
Key words: White-tailed deer, diet, microhistological analysis, feces, Sierra Madre de Oaxaca.
Resumen
La región de la Sierra Madre de Oaxaca, ubicada al norte del estado de Oaxaca, México, es una zona de ecosistemas con alta actividad forestal; en algunas áreas sus bosques templados son conservados por iniciativas de las comunidades indígenas que ahí habitan. Dentro de estos bosques, se analizó la dieta del venado cola blanca (Odocoileus virginianus) en San Miguel Amatlán y Santa Catarina Lachatao entre junio 1998 y agosto 1999. Se utilizó el análisis microhistológico de heces fecales, la observación de rastros de ramoneo en plantas y el análisis de contenidos estomacales. La dieta anual estuvo constituida por 42 especies de 23 familias vegetales. Las familias con el mayor número de especies fueron: Fagaceae, Asteraceae, Ericaceae y Fabaceae. La diversidad de la dieta durante la estación húmeda y seca no presentó diferencias significativas (H’= 0.918 y H’=0.867 respectivamente). El porcentaje de similitud entre ambas temporadas fue de 58%. La diferencia entre la frecuencia de aparición de las especies vegetales fue mayor en la estación húmeda. Del total de especies consumidas, el mayor porcentaje estuvo constituido por las herbáceas. Las especies preferidas a lo largo del año fueron: Sedum dendroideum (herbácea) y Satureja macrostema (arbusto). La riqueza de especies en la dieta fue menor a la observada en otras regiones con bosques tropicales y bosques mixtos, pero similar a la obtenida en un bosque tropical seco en Costa Rica y en los bosques de coníferas en Estados Unidos.
Palabras clave: venado cola blanca, dieta, análisis microhistológico, heces fecales, Sierra Madre de Oaxaca.
El venado cola blanca (Odocoileus virginianus) se distribuye a lo largo de todo México, excepto en la península de Baja California y el norte de Sonora (Hall 1981). En el estado de Oaxaca al sur de México, habitan cuatro subespecies de venado cola blanca (Odocoileus virginianus acapulcensis, O. v. oaxacensis, O. v. thomasi y O. v. toltecus) en diferentes tipos de bosques templados y tropicales (Briones- Salas & Sánchez-Cordero 2004).
La región de la Sierra Madre de Oaxaca, al norte del estado, tiene una superficie aproximada de 943 796ha y cerca del 70% de sus bosques se encuentran en buen estado de conservación, sobre todo los templados y nublados (Ortega del Valle et al. 2010). En parte de estos bosques, específicamente en los Municipios de Santa Catarina Lachatao y San Miguel Amatlán, empresas forestales particulares realizaron aprovechamiento forestal y de fauna silvestre durante casi 20 años sin un manejo adecuado, particularmente de venado cola blanca. Es muy posible que por esta razón, la población de esta especie haya decrecido drásticamente, de acuerdo con la información proporcionada por pobladores indígenas zapotecos y estudios realizados en la zona (2.6 a 3.9 venados/ km2; Briones-Salas & García 2005 y 1.13 a 1.15 venados/km2; Ortiz-Martínez et al. 2005). Posteriormente, a principios de los años 1990’s, la recuperación de sus bosques mediante el cese de concesiones a las empresas privadas, trajo consigo el interés para implementar procesos de recuperación y conservación mediante la gestión y ejecución de planes de manejo (Chapela 1999). Esto incluyó a la población de venado cola blanca, por ser la especie de fauna silvestre más importante para las comunidades indígenas de la región debido a su valor de uso. Es por ello que se implementó un proyecto de investigación con el objetivo de conocer la situación real de la especie basada en su densidad poblacional, uso de hábitat y dieta.
Entre los rumiantes, el venado es considerado como una especie herbívora ramoneadora, oportunista en su alimentación; sin embargo, en ocasiones selecciona a las especies vegetales con alto contenido de nutrientes (Church 1993, Galindo-Leal & Weber 1998), principalmente en áreas templadas (Hanley 1997) o bien en sitios perturbados (Ortiz-Martínez et al. 2005). Sin embargo, en regiones donde las plantas tienen bajo contenido nutricional o este varía en alguna estación del año, la especie consume una mayor diversidad para compensar estas deficiencias (DiMare 1994, Mandujano et al. 2002).
Las diferencias en la diversidad de las especies vegetales en la dieta del venado cola blanca están determinadas por la distribución geográfica, los tipos de vegetación, época del año y por las condiciones del hábitat, que afectan la disponibilidad y calidad nutricional de las plantas (Arceo et al. 2005). En bosques templados de Norteamérica y México, la riqueza varía entre 15 y 135 especies (Gallina 1993, Johnson et al. 1995, Galindo-Leal & Weber 1998, Villarreal 2000), mientras que en los bosques tropicales de Venezuela, Costa Rica y México se han encontrado desde 45 hasta 125 especies en la dieta (Granado 1989, DiMare 1994, Arceo et al. 2005). Está demostrado que los bosques tropicales son más diversos, comparados con los bosques templados; sin embargo, la gran variación que existe respecto al número de especies consumidas por el venado cola blanca en ambos tipos de bosque, refleja que los cambios estacionales y la condición del hábitat influyen significativamente en la disponibilidad y calidad del alimento.
El conocimiento de la dieta del venado cola blanca durante las diferentes estaciones del año permite determinar qué elementos vegetales son indispensables, las temporadas críticas del año y la capacidad de adaptación de la especie a las variaciones en la disponibilidad de alimento (Arceo et al. 2005). El presente estudio tiene como objetivo conocer la disponibilidad estacional de los recursos vegetales y su uso por el venado cola blanca en los bosques templados de la Sierra Madre de Oaxaca, mediante el análisis de la composición botánica de la dieta, durante la estación seca y húmeda del año. Esto como base para definir estrategias futuras de conservación (conocimiento, cuidado y uso sustentable) de la especie en una zona con una fuerte actividad forestal, que puedan beneficiar a las comunidades indígenas zapotecas que habitan esta región.
Materiales y métodos
Área de estudio: El estudio se llevó a cabo en las localidades de San Miguel Amatlán y Santa Catarina Lachatao, ubicados en el distrito de Ixtlán, en la región de la Sierra Madre de Oaxaca, entre los paralelos 17°05’-17°10’ N - 96°20’-96°30’ W (INEGI 2007). El área se encuentra en un sistema montañoso que ocupa la mitad septentrional del estado de Oaxaca con un total de 13 570ha, caracterizado por tener topografía muy accidentada, con altitud variable entre los 2 400 y 3 300m (Ortiz Pérez et al. 2004). El clima es templado subhúmedo con lluvias de verano. La vegetación está conformada por bosque de Pinus-Quercus que cubre el 56% de la superficie total, el bosque de Quercus-Pinus (21.8%), bosque de Abies (1.2%), Pinus-Abies (0.35%.), Pinus (0.03%) y zonas agrícolas (12.6%). La zona en general tiene una fuerte actividad forestal (CEMASREN 1999) y un empobrecimiento en la cobertura vegetal (Ortiz-Martínez et al. 2005).
Recolección de muestras y análisis de laboratorio: Para determinar la composición botánica de la dieta del venado cola blanca se utilizó: el análisis microhistológico de heces fecales, la observación de plantas ramoneadas por el venado cola blanca y el análisis de contenidos estomacales (Middleton & Sánchez 1994). La información sobre las dos primeras, se recolectó en 12 itinerarios de censo fijos (transectos) de 400m de longitud, distribuidos de acuerdo con la superficie cubierta por cada tipo de vegetación (Tellería 1986, Wilson et al. 1996). Se establecieron cinco itinerarios de censo fijos en el bosque de Pinus-Quercus, tres en el bosque de Quercus-Pinus, dos en el de Abies y uno en el bosque de Pinus-Abies y en el de Pinus. El área de muestreo abarcó un total de 1 200ha.
Las recolectas se realizaron entre junio 1998 y agosto 1999. Se llevaron a cabo seis muestreos, cada dos meses, tres en la estación seca y tres durante la estación húmeda del año. Se recolectaron un total de 28 grupos fecales; debido a que según Anthony & Smith (1974), una muestra entre 15 y 50 grupos fecales es adecuada para determinar la dieta estacional. Cada grupo fue rotulado con la fecha de recolecta, localidad y tipo de vegetación.
Posteriormente, se procedió con la identificación y cuantificación de los tejidos epidérmicos vegetales presentes en las heces fecales. El primer paso fue elaborar una colección de referencia de las plantas disponibles en el área de estudio; para ello, se recolectaron muestras de todas las plantas con alturas no mayores a los dos metros y que se encontraron a lo largo de los itinerarios fijos. Las muestras se secaron a 75°C y después fueron trituradas. A las muestras de plantas de referencia y heces fecales, se les realizó un tratamiento con el objeto de disminuir la presencia de pigmentos y algunos tejidos blandos (Middleton & Sánchez 1994).
Una muestra de hojas, flores y tallos de cada especie vegetal se colocó en hidróxido de sodio al 5% y se hirvió por espacio de cinco minutos, se enjuagó con agua destilada y se le agregó hipoclorito de sodio, dejando la muestra hasta su decoloración. Se volvió a enjuagar con agua destilada y se colocó en diluciones de alcohol en concentraciones de 30, 50, 80, 90%, y absoluto, durante 20min en cada una para lograr la deshidratación; enseguida se agregó xilol para aclarar los tejidos vegetales. Por último, una pequeña porción del material tratado se colocó sobre un portaobjetos, se mezcló con resina sintética, se colocó en el cubreobjetos y se selló con esmalte transparente.
Para la elaboración de las laminillas de las heces fecales se aplicó el mismo procedimiento, sólo que en el momento de montar las muestras en el portaobjetos se utilizó una laminilla metálica con orificios de aproximadamente 7mm de diámetro. Se realizaron 10 preparaciones mixtas de los grupos fecales recolectados, cinco de la estación húmeda y cinco de la estación seca. Para identificar y cuantificar las especies de plantas presentes en la dieta, se colocaron porciones similares del material tratado en cada uno de los orificios, también mezclado con resina sintética y sellada con esmalte. La identificación microhistológica se llevó a cabo con un microscopio compuesto con ocular de 10x y objetivos de 10, 20 y 40x. De manera sistemática se observaron 20 campos con el objetivo de 10x y ocular de 10x (100 aumentos) para cada una de las cinco preparaciones mixtas. Se observó un total de 100 campos por estación. Se consideraron las siguientes características epidermales e histológicas: tamaño y forma de los estomas y pelos, tricomas, glándulas, cristales, drusas, paredes celulares, células de silicio y disposición de las células en el tejido epidérmico (Peña & Habib 1980, Middleton & Sánchez 1994). Se realizó una colección fotográfica que se utilizó como referencia para identificar los fragmentos que se encontraron en las heces fecales.
Los recorridos para observar las plantas ramoneadas se realizaron al mismo tiempo y en los mismos sitios en los que se recolectaron las heces fecales. Además de esto, un guía local apoyó en la identificación del ramoneo por el venado cola blanca, con lo que disminuimos el riesgo de confundir éste con las marcas que deja el ganado bovino. Una de las características que se consideró para determinar si la planta había sido ramoneada por el venado fue que presentara rastros de mordiscos en brotes tiernos y hojas a una altura máxima de dos metros del suelo. Posteriormente, se recolectó una muestra de cada planta, se prensó, secó y determinó. Estas también formaron parte de la colección de referencia para los análisis microhistológicos. Finalmente, se analizó el contenido de dos estómagos que fueron proporcionados por cazadores de la zona, ambas muestras correspondieron a la estación húmeda. Los contenidos estomacales fueron conservados en alcohol al 70% y etiquetados con los datos de cada animal. Se analizaron los fragmentos de las plantas para determinar la especie, mediante un microscopio compuesto con aumentos de 10, 20 y 40x. La identificación de la especie se basó en las características de las microestructuras de la planta, pelos y tricomas, así como la forma de las hojas, tallos, bordes, semillas o frutos; posteriormente, fueron comparados con la colección de referencia.
El análisis estadístico se aplicó a los datos obtenidos de las especies vegetales encontradas mediante la técnica microhistológica y a las observaciones de ramoneo. Se obtuvo la frecuencia de aparición de cada especie y/o género, obtenida del análisis microhistológico; y la frecuencia de observación de las especies ramoneadas en los itinerarios fijos. A partir de las frecuencias obtenidas, se calculó la densidad relativa por medio del cuadro de Fracker & Brischle (Peña & Habib 1980). Asimismo, se estimaron el aporte porcentual de cada especie vegetal y la riqueza y diversidad de especies en la dieta durante las dos temporadas del año, aplicando el índice de Shannon-Wiener (H’) log base 10. Los índices fueron comparados entre sí mediante el uso de una t-Student modificada por Hutcheson (Zar 1999). Para conocer el nivel de dependencia de la dieta a los cambios estacionales, se utilizó la prueba de heterogeneidad de ji cuadrada (Zar 1999).
Se estimó el porcentaje de similitud en la composición botánica de la dieta a nivel de especie, entre la estación seca y húmeda, mediante el índice de Kulczynski (Hubálek 1982). Por último, se estimó el índice de preferencia sobre las especies vegetales determinadas en cada estación del año (Korschgen 1980). Para ello, se dividió el porcentaje estimado de cada alimento en la dieta del venado entre el porcentaje estimado del alimento en el hábitat (basado en la disponibilidad del recurso). Los datos de la biomasa disponible de las especies vegetales en la zona estudiada se obtuvieron del trabajo realizado por Ortiz-Martínez et al. (2005), durante el mismo periodo y en los mismos sitios de muestreo de este estudio. Los valores del índice mayores a uno indicaron preferencia, valores menores a uno indicaron rechazo o no preferencia y, cuando el valor se aproximó a uno indicó que el alimento fue consumido en igual proporción que su disponibilidad.
Resultados
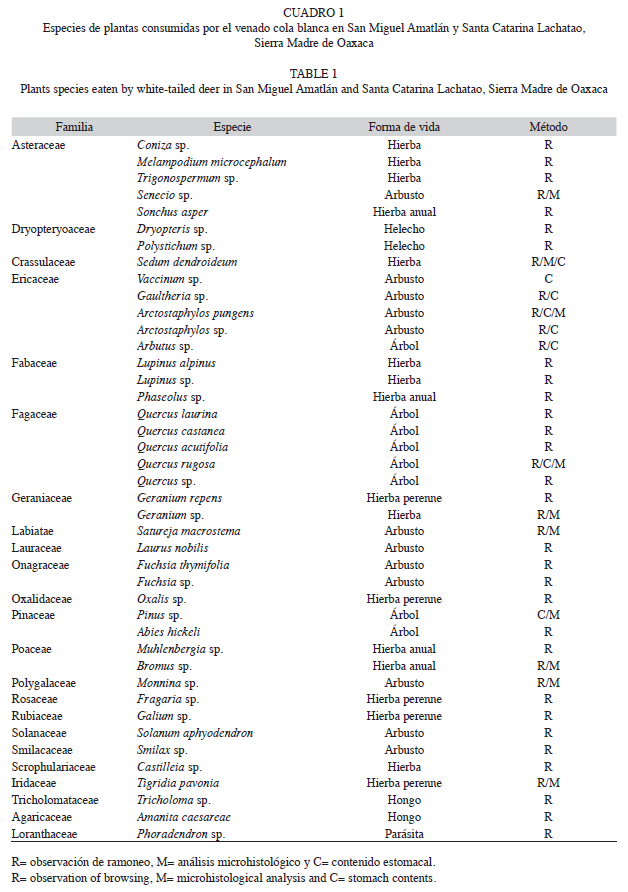
La dieta anual del venado cola blanca estuvo constituida por 42 especies vegetales incluidas en 23 familias botánicas. Mediante el análisis microhistológico se registraron 10 especies de 10 familias; por medio de la observación del ramoneo se registraron 40 especies de 23 familias y finalmente, mediante el análisis de contenido estomacal se determinaron ocho especies de cuatro familias. Las familias mejor representadas en la dieta fueron: Fagaceae (seis especies), Asteraceae (cinco), Ericaceae (cinco) y Fabaceae (tres). La mayor riqueza se encontró en la estación húmeda con 42 especies y la menor en la estación seca con 27 especies (Cuadro 1).
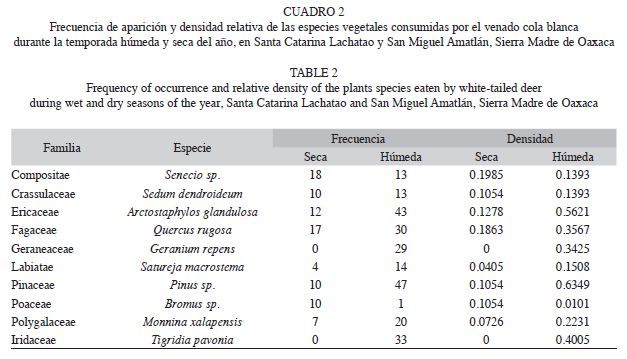
Las especies arbóreas y arbustivas fueron las más frecuentes en la dieta; sin embargo, el 42% de las especies consumidas fueron herbáceas, 25.5% arbustos, 20.9% árboles, 4.6% hongos y helechos, y 2.3% una planta parásita. Los géneros y especies que presentaron la mayor frecuencia y densidad relativa (expresada en porcentaje), durante la estación seca fueron: Senecio sp. (n=57 y 0.522%), Arctostaphylos glandulosa (n=41 y 0.456), Sedum dendroideum (n=40 y 0.421), Satureja macrostema (n=38 y 0.417), Pinus sp. (n=36 y 0.398), Quercus rugosa (n=29 y 0.304) y Bromus sp. (n=24 y 0.278). Mientras que durante la estación húmeda, fueron: Pinus sp. (n=77 y 0.772), A. glandulosa (n=73 y 0.724), Senecio sp. (n=68 y 0.701), Geranium repens (n=55 y 0.631), Tigridia pavonia (n=51 y 0.608), Quercus rugosa (n=36 y 0.600), Vaccinum sp. (n=36 y 0.600), Satureja macrostema (n=31 y 0.568) y Muhlenbergia sp. (n=31 y 0.557). La frecuencia de aparición de las especies fue significativamente mayor durante la estación húmeda (x2=56.22, g.l.=41, p<0.05) (Cuadro 2).
Los valores de diversidad de la dieta fueron estadísticamente significativas (t=0.01, p=0.02); mayores en la estación húmeda (H’=2.957, p=0.06) y menores en la estación seca (H’=1.832, p=0.05). El valor de equitatividad obtenido durante la estación seca fue E=0.960 y en la húmeda E=0.918, lo que indicó un elevado grado de uniformidad en las abundancias relativas de las especies en ambas estaciones. El índice de similitud en la composición botánica de la dieta entre cada estación fue de 56%.
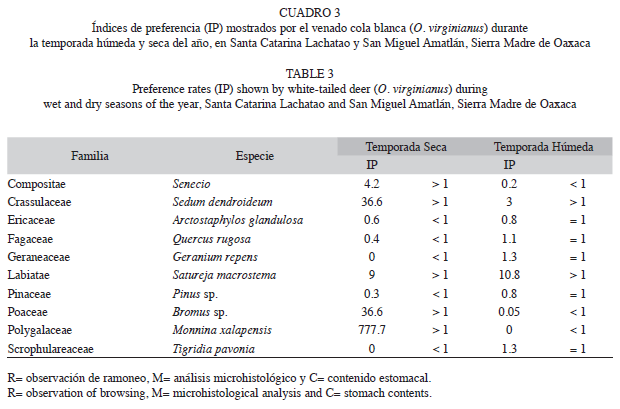
Las especies que fueron preferidas durante ambas estaciones del año mediante el índice de preferencia (Ip), fueron: Senecio sp. (Ip=42.2 en la estación seca, Ip=7.1 en la estación húmeda), A. pungens (Ip=38.1 y 49.0) y Satureja macrostema (Ip=19.0, Ip=10.8, respectivamente). Las especies preferidas durante la estación húmeda fueron Geranium repens (Ip=51.3, Ip=0) y Tigridia pavonia (Ip=22.1 y Ip=0). Por último, las especies preferidas durante la temporada seca y consumidas de acuerdo con su disponibilidad o bien rechazadas durante la estación húmeda fueron: M. xalapensis (Ip=777.7, Ip=0.02) y Bromus sp. (Ip=36.6, Ip=0.05). Las 35 especies restantes fueron consumidas de acuerdo con la disponibilidad en alguna de las estaciones del año (Cuadro 3).
Discusión
La riqueza de especies en la dieta del venado cola blanca encontrada en este estudio (42 especies) con las tres técnicas utilizadas, resultó mucho menor, comparada con la obtenida en estudios realizados en Venezuela (125 especies, Granado 1989) y México (82 especies, Arceo et al. 2005) en áreas con bosque tropical, mezclado con áreas de cultivo y bosque tropical caducifolio, respectivamente. Así también, fue menor que en un hábitat con bosque de pinoencino, en el estado de Durango, México (135 especies, Gallina 1993). Sin embargo, resultó similar a las encontradas en un bosque de coníferas en Canadá (23 especies, Skinner & Telfer 1974), en un chaparral mezclado con pastizal en EEUU (39 especies, Kie et al. 1980), y un bosque tropical caducifolio en Costa Rica (45 especies, DiMare 1994).
Si bien es cierto que el venado cola blanca puede incluir en su dieta más de 100 especies vegetales, el hecho de que en algunas regiones distribuidas en diferentes latitudes, incluyendo este estudio, se haya encontrado una riqueza menor, sugiere que más que la latitud y el tamaño de la especie (Arceo et al. 2005), es la calidad del hábitat y la estacionalidad lo que determina la calidad de la dieta.
El historial y las técnicas de aprovechamiento forestal llevadas a cabo en el área de estudio en años anteriores al desarrollo del trabajo, influyeron significativamente en la baja riqueza y abundancia de especies vegetales en el sotobosque, independientemente del tipo de vegetación. Las técnicas utilizadas consistieron en talar todos los árboles maderables con los mejores atributos y extraerlos del área mediante el arrastre, lo que elimina toda la cubierta vegetal existente en el sotobosque. Esto se reflejó en los resultados obtenidos sobre la estimación de la biomasa vegetal, que resultó baja (58kg/ha) (Ortiz-Martínez et al. 2005), comparada con lo estimado por Gallina (1993) en bosques similares (297kg/ha). Otro factor que pudo influir en el resultado fue que el periodo de muestreo se caracterizó por ser atípico, debido a la sequía más drástica comparada con años anteriores, lo que pudo afectar la disponibilidad de especies sensibles a la falta de humedad y reflejarse en la baja productividad de la biomasa vegetal (Ortiz-Martínez et al. 2005).
Por otro lado, el bajo número de grupos fecales encontrados (28), comparado con estudios del mismo tipo, está relacionado con la densidad poblacional de venado cola blanca encontrada en la misma área de estudio. El resultado fue de 1.13 venados por km2 (Ortiz- Martínez et al. 2005), mientras que en otras regiones del país y del mismo estado de Oaxaca se han encontrado densidades por arriba de los 5.0 venados por km2 (Mandujano 1992, Gallina 1993, Duarte 2004).
Con respecto a la riqueza de especies vegetales y su abundancia relativa entre estaciones, ambas fueron similares. De acuerdo con el índice de similitud, el 56% de las especies que consumió en la temporada seca las incluyó también en la temporada lluviosa. Esto posiblemente se debe a que las plantas consumidas en ambas temporadas presentan densidades constantes a lo largo del año.
Las herbáceas fueron las mejor representadas en la dieta del venado, comparado con las demás formas de vida de las plantas. Sin embargo, dentro de las especies preferidas estuvieron incluidas tanto las hierbas como los arbustos. El índice de preferencia mostró que las especies consumidas en la estación seca dejaron de ser importantes para el venado durante la estación húmeda con excepción de la especie S. macrostema. Esta última especie se caracteriza por la presencia de antioxidantes con varias propiedades curativas (Alonso 2009) y S. dendroideum es una especie de rápido crecimiento, con alto contenido de nutrientes y agua, y se adapta a condiciones extremas como falta de humedad. Probablemente la preferencia por estas especies tuvo que ver con la cantidad de nutrientes y la proporción de agua en una temporada tan crítica como la sequia que ocurrió durante el año de muestreo. M. xalapensis y Bromus sp., tienen un alto contenido nutricional, esta última especie es utilizada para la engorda y producción de leche en ovinos en algunas regiones del país y en Sudamérica (Siebald 2002).
Varios estudios han demostrado que el venado consume un número mínimo de pastos, ya que no tolera una lenta digestión de fibras, por presentar un pequeño rumen con relación al tamaño corporal y necesita compensar su menor capacidad rumino-reticular con alimentos de alta calidad (Clemente 1984, Gallina 1993, Arceo et al. 2005, Galindo-Leal & Weber 1998). En este caso solo incluyó una especie de gramínea, pero su consumo fue frecuente y preferido (Ip) durante la estación seca del año. Indudablemente, estas especies fueron las más importantes en la dieta del venado cola blanca en ambas o en alguna estación del año; sin embargo, un análisis bromatológico permitiría fundamentar con mayor claridad el por qué de su preferencia.
Por otra parte, entre las especies que, aunque no fueron preferidas pero que fueron frecuentes en alguna temporada del año, estuvo Quercus rugosa cuyo consumo pudo estar relacionado con el aporte energético y vitamínico que le provee tanto el follaje como las bellotas, éstas últimas a través del consumo de sus aceites. En este sentido, las características de las hojas de los encinos son una buena defensa contra la herbivoría (Clemente et al. 2005). Otro caso fue el de Arctostaphylus glandulosa y de Geranium repens, dos especies características en la dieta del venado cola blanca en los bosques templados de otras regiones del país, seleccionadas probablemente por su alta digestibilidad y contenido de proteína cruda. La escogencia de plantas ricas en nutrientes de rápida fermentación y fácilmente digeribles son las ideales para la especie.
Por último, la frecuencia de especies del género Pinus fue considerable, estas especies poseen terpenos que son sustancias aparentemente desagradables, indigeribles y tóxicas para la mayoría de los animales, por lo que muy pocas intentan consumir el follaje o las ramas tiernas de esos árboles. Es probable, que el venado haya consumido las hojas secas, para así evitar los efectos negativos y mejorar la digestión por el alto contenido de fibra (Clemente et al. 2005). La frecuencia de esta especie fue mayor en la temporada de lluvias, lo que podría indicar la necesidad de consumo de fibra ante la gran cantidad de alimento fresco.
Los resultados reflejan que al menos durante el periodo en que se realizó el estudio, el sitio presentó posiblemente condiciones de estrés para los venados. El hecho de que hayan consumido las hojas de pinos, especies fagodisuasivas y de consumo poco frecuente en otras áreas, fue quizá por la ausencia de plantas con mayor valor nutricional, aunado a la baja productividad en la biomasa. Aunque actualmente el aprovechamiento forestal en el área de estudio se lleva a cabo mediante un manejo regulado, los efectos de empobrecimiento de la cobertura vegetal continúan presentes, y es posiblemente una causa importante de las bajas densidades de esta especie.
Agradecimientos
A las autoridades municipales de San Miguel Amatlán, Santa Catarina Lachatao, San Antonio Cuajimoloyas y San Isidro Llano Grande, por el permiso para trabajar en sus
terrenos comunales. A F.J. Vargas, por colaborar activamente en el trabajo de campo. A.G. Sánchez- Rojas y G. Ramos-Fernández por aportar sugerencias para mejorar las diversas versiones del documento. La CGEPI del Instituto Politécnico Nacional (978031), el Fondo Mexicano para la Conservación de la Naturaleza (A1- 98/006) y el Sistema de Investigación Benito Juárez, CONACyT (20000506023) financiaron parcialmente la realización del proyecto. La COFAA del IPN, apoya económicamente a los autores. MB-S agradece a EDI del IPN y al Sistema Nacional de Investigadores por su apoyo.
Referencias
Alonso, C.N. 2009. Actividad antioxidante de Satureja macrostema. Tesis de Maestría, Escuela Nacional de Ciencias Biológicas, Instituto Politécnico Nacional, México, D.F., México. [ Links ]
Anthony, R.G. & N.S. Smith. 1974. Comparison of rumen and fecal analysis to describe deer diets. J. Wildl. Manage. 38: 535-540. [ Links ]
Arceo, G., S. Mandujano & S. Gallina. 2005. Diet diversity of white-tailed deer (Odocoileus virginianus) in a tropical dry forest in Mexico. Mammalia 69: 159-168. [ Links ]
Briones-Salas, M. & C. García. 2005. Estimación de la densidad del venado cola blanca (Odocoileus virginianus oaxacensis) en la Sierra Norte de Oaxaca. Rev. Mex. Mastozool. 9: 128-135. [ Links ]
Briones-Salas, M. & V. Sánchez-Cordero. 2004. Mamíferos, p. 423-447. In A.J. García-Mendoza, M.J. Ordóñez & M. Briones-Salas (eds.). Biodiversidad de Oaxaca. Instituto de Biología, UNAM-Fondo Oaxaqueño para la Conservación de la Naturaleza-World Wildlife Fund, México. [ Links ]
CEMASREN. 1999. Resultados de manejo forestal para el aprovechamiento de recursos forestales maderables en la comunidad de Pueblos Mancomunados de Lachatao, Amatlán, Yavesía y anexos, de los mismos municipios, distrito de Ixtlán, Estado de Oaxaca, resultados del proceso de datos. Vol. II. Oaxaca, Oaxaca, México. [ Links ]
Chapela, F. 1999. Silvicultura comunitaria en la sierra norte de Oaxaca, el caso de la unión zapoteco-chinanteca. Red de Gestión de Recursos Naturales-Fundación Rockefeller, México, D.F., México. [ Links ]
Church, D.C. 1993. El rumiante, fisiología digestiva y nutrición. Acribia, Zaragoza, España. [ Links ]
Clemente, S.F. 1984. Utilización de la vegetación nativa en la alimentación del venado cola blanca (Odocoileus virginianus) en el estado de Aguascalientes. Tesis de Maestría, Colegio de Posgraduados, Instituto de Enseñanza e Investigación en Ciencias Agrícolas,
Chapingo, Estado de México, México. [ Links ]
Clemente, F., E. Riquelme, G.D. Mendoza, G. Bárcena, S. González & R. Ricalde. 2005. Digestibility of forage diets of white-tailed deer (Odocoileus virginianus, Hays) using different ruminal fluid inocula. J. App. Ann. Res. 27: 71-76. [ Links ]
DiMare, M.I. 1994. Hábitos alimentarios del venado cola blanca en la Isla San Lucas, Puntarenas, Costa Rica, p. 73-90. In C. Vaughan & M. Rodríguez (eds.). Ecología y manejo del venado cola blanca en México y Costa Rica. EUNA, Heredia, Costa Rica. [ Links ]
Duarte, M. 2004. Densidad poblacional del venado cola blanca, en el área comunal de Ixtepeji, Sierra Norte, Oaxaca. Tesis de Licenciatura, Instituto Tecnológico Agropecuario de Oaxaca, Oaxaca, México. [ Links ]
Gallina, S. 1993. White-tailed deer and cattle diets at La Michilia, Durango, México. J. Range Manage. 46: 487-492. [ Links ]
Galindo-Leal, C. & M. Weber. 1998. El venado de la Sierra Madre Occidental: Ecología, manejo y conservación. EDICUSA-CONABIO, México, D.F. [ Links ]
Granado, A. 1989. Dieta del venado caramerudo (Odocoileus virginianus gymnotis) en El Socorro, estado Guarico. Tesis de Grado, Universidad Central de Venezuela, Caracas, Venezuela. [ Links ]
Hall, E.R. 1981. The mammals of North America. John Wiley & Sons, NewYork, EEUU. [ Links ]
Hanley, T.A. 1997. A nutritional view of under standing and complexity in the problem of diet selection by deer (Cervidae). Oikos 79: 209-218. [ Links ]
Hubálek, Z. 1982. Coefficients of association and similarity, based on binary (presence and absence) data: an evaluation. Biol. Rev. 87: 669-689. [ Links ]
INEGI. 2007. Carta de suelo y vegetación 1:250 000. Oaxaca, E 14-9. [ Links ]
Johnson, A.S., P.E. Hale, W.M. Ford, J.M. Went-Worth, J.R. French, O.F. Anderson & G.B. Pullen. 1995. White-tailed deer foraging in relation to successional stage, overstory type and management of Southern Appalachian forests. Am. Midl. Nat. 133: 18-35. [ Links ]
Kie, J.G., D.L. Drawe & G. Scott. 1980. Changes in diet and nutrition with increased herd size in Texas whitetailed deer. J. Range Manage. 33: 28-34. [ Links ]
Korschgen, J.J. 1980. Procedures for food-habitat analysis, p. 113-128. In S.D. Schemnitz (ed.). Wildlife Management Techniques Manual. The Wildlife Society, Washington, D.C., EEUU. [ Links ]
Mandujano, S. 1992. Estimaciones de la densidad poblacional del venado cola blanca (Odocoileus virginianus) en un bosque tropical de Jalisco. Tesis de Maestría, Universidad Nacional Autónoma de México, México, D.F. [ Links ]
Mandujano, S., S. Gallina, G. Arceo, G. Sánchez-Rojas & G. Silva-Villalobos. 2002. Venado Cola Blanca (Odocoileus virginianus sinaloae J.A. Allen), p. 415-422. In F. Noguera, J.H. Vega-Rivera, A.N. García-Aldrete & M. Quesada-Avendaño (eds.). Historia Natural de Chamela. Instituto de Biología, UNAM, México, D.F. [ Links ]
Middleton, B.A. & R.E. Sánchez. 1994. Microhistological analysis of the food habits of herbivores in the tropics. Vida Sil. Neotr. 13: 41-47. [ Links ]
Ortega del Valle, D., G. Sánchez, C. Solano, M.A. Huerta, V. Meza & C. Galindo. 2010. Áreas de conservación certificadas en el estado de Oaxaca. World Wildlife Fund, Comisión Nacional de Áreas Naturales Protegidas y Secretaría del Medio Ambiente y Recursos Naturales, México. [ Links ]
Ortiz-Martínez, T., S. Gallina, M. Briones-Salas & G. González. 2005. Densidad poblacional y caracterización del hábitat del venado cola blanca (Odocoileus virginianus oaxacensis, Goldman y Kellog, 1940) en un bosque templado de la Sierra Norte de Oaxaca, México. Acta Zool. Mex. (n.s.) 21: 65-78. [ Links ]
Ortiz Pérez, M.A., J.R. Hernández Santana & J.M. Figueroa Mah Eng. 2004. Reconocimiento fisiográfico y geomorfológico, p 43-54. In A.J. García-Mendoza, M.J. Ordoñez & M. Briones-Salas (eds.). Biodiversidad de Oaxaca, Instituto de Biología, UNAM-Fondo Oaxaqueño para la Conservación de la Naturaleza- World Wildlife Fund, México, D.F. [ Links ]
Peña, N.M. & R.P. Habib. 1980. La técnica microhistológica: Un método para determinar la composición botánica de la dieta de herbívoros. Serie Técnico Científica, Departamento de Manejo de Pastizales, INIP-SARH, México, D.F. [ Links ]
Siebald, E. 2002. Forrajeras Nativas y Naturalizadas: Un gran valor. Tierra Adentro 24: 33-35. [ Links ]
Skinner, W.R. & E.S. Telfer. 1974. Spring, summer and fall foods of deer in New Brunswick. J. Wildl. Manage. 38: 210-214. [ Links ]
Tellería, J.L. 1986. Manual para el censo de los vertebrados terrestres. Raíces, Madrid, España. [ Links ]
Villarreal, J. 2000. Venado cola blanca: Manejo y aprovechamiento cinegético. Unión Ganadera Regional de Nuevo León, Monterrey, Nuevo León, México. [ Links ]
Wilson, D.E., F. Rusell, J.D. Nichols, R. Rudran & M.S. Foster (eds.). 1996. Measuring and monitoring biological diversity. Standard methods for mammals. Smithsonian Institution, Washington, D.C., EEUU. [ Links ]
Zar, L.H. 1999. Bioestatistical Analysis. Prentice Hall, Upper Saddle River, Nueva Jersey, EEUU. [ Links ]
*Correspondencia:
Graciela González:Laboratorio de Vertebrados Terrestres (Mastozoología). Centro Interdisciplinario de Investigación para el Desarrollo Integral Regional, Unidad Oaxaca, Instituto Politécnico Nacional. Calle Hornos no. 1003, Sta. Cruz Xoxocotlán, Oaxaca. México, C.P. 71230; gracielagonzalezp@hotmail.com
Miguel Briones-Salas: Laboratorio de Vertebrados Terrestres (Mastozoología). Centro Interdisciplinario de Investigación para el Desarrollo Integral Regional, Unidad Oaxaca, Instituto Politécnico Nacional. Calle Hornos no. 1003, Sta. Cruz Xoxocotlán, Oaxaca. México, C.P. 71230; mbriones@ipn.mx
1. Laboratorio de Vertebrados Terrestres (Mastozoología). Centro Interdisciplinario de Investigación para el Desarrollo Integral Regional, Unidad Oaxaca, Instituto Politécnico Nacional. Calle Hornos no. 1003, Sta. Cruz Xoxocotlán, Oaxaca. México, C.P. 71230; mbriones@ipn.mx, gracielagonzalezp@hotmail.com
Recibido 07-III-2011. Corregido 06-VIII-2011. Aceptado 02-IX-2011.


{kind=link}
{kind=link}
{kind=link}