










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.48 n.2-3 San José Jun. 2000
y distribución geográfica de Euspondylus acutirostris y descripción
de un nuevo Euspondylus del nordeste de Venezuela
Abraham Mijares-Urrutia1 J. Celsa Señaris2 & Alexis Arends1
Recibido 28-IV-1999. Corregido 21-I-2000. Aceptado 4-II-2000.
Abstract
Only two species of the microteiid lizard genus Euspondylus (E. acutirostris and E. phelpsorum) had been reported from Venezuela. New records of the poorly known gymnophthalmid lizard Euspondylus acutirostris are reported extending its known range along the Coastal Range and Sierra de Aroa (north-central Venezuela) and Sierra de San Luis (northwestern Venezuela), all records occurring at the cloud forest above 1000 m. Seven body measurements were included and morphological variation is described based on at least 17 new specimens. Sexual dimorphism is determined in, at least, eight characteristics of size and squamation. Specimens were found in epiphytic bromeliads and the ground. The populations from Sierra de Aroa and Sierra de San Luis (only one specimen known from each locality), differ from those of the Coastal Range (supposedly the nominal population) in some morphological and coloration features, suggesting that the former two could represent different taxonomical entities. A new species of Euspondylus is described based on an female (taken within a bromeliad) from Cerro El Humo, Sucre, northeastern Venezuela. It differs from congeners mainly by having keeled scales on the limbs and a very acute snout.
Key words
Euspondylus acutirostris, Euspondylus sp., Teiidae, Taxonomy, Squamata, Venezuela.
En Venezuela se conocen 34 especies entre 13 géneros (La Marca 1997) de lagartijos microtéidos (Familia Teiidae [Gymnophthalmidae, auctorum], Myers & Donnelly 1996). La mayoría se conocen por escasos ejemplares representativos, un hecho que dificulta la resolución de las relaciones taxonómicas (La Marca & García-Pérez 1990) y la identificación de nuevas especies. Un caso de estos, Euspondylus, es caracterizado en la clave genérica de Peters & Donoso-Barros (1970) por tener las escamas nasales anteriores separadas por las escamas rostral y frontonasal, abertura auricular presente, párpados móviles, disco palpebral inferior transparente, pólex normal (uña presente), escamas dorsales no granuliformes, prefrontales presentes, escamas occipitales presentes (pero los bordes posteriores de las parietales e interparietal no son parejamente transversos ni están alineados), escamas laterales de tamaño claramente reducido, escamas laterales del cuello no granuliformes y palpebral inferior no en forma de espejuelo circular. La especie tipo es Euspondylus maculatus de Perú y el sur de Ecuador. Las especies venezolanas de Euspondylus se ajustan adecuadamente a esta clave diagnóstica, sin embargo, es cuestionable el que éstas características impliquen que exista afinidad entre las ocho especies actualmente asignadas al género (Uzzell 1973, Oftedal 1974).
Dos especies de Euspondylus son endémicas de Venezuela Euspondylus acutirostris Peters (1862) de la Cordillera de la Costa, incluyendo el Parque Nacional Henri Pittier, una de las regiones de Venezuela mejor estudiada faunísticamente; E. phelpsorum Lancini (1968, como E. phelpsi, corregido por Myers & Donnelly 1996), descrita con base en un único ejemplar (en esta especie incluimos, tentativamente, a E. goeleti, considerado un sinónimo de E. phelpsorum por Gorzula & Señaris 1998), cuya distribución abarca las cimas de dos remotos y poco explorados tepuyes al sur de Venezuela, Cerro Jaua y Cerro Yaví, respectivamente. La situación de la nomenclatura y taxonomía de ésta última especie es incierta y requiere de una urgente revisión, aunque su discusión está fuera del alcance de este artículo.
Peters (1862) describió Euspondylus acutirostris con base en un sólo ejemplar. Un ejemplar adicional fue registrado por Test et al. (1966). Para este estudio localizamos 17 ejemplares adicionales, los cuales permitirán hacer una primera aproximación de su variabilidad morfológica, distribución geográfica e historia natural. También, se describe una nueva especie de Euspondylus de las alturas intermedias de la región montañosa del extremo nororiental de Venezuela. Aunque éste último taxon se describe a partir de un único ejemplar, confiamos en que exhibe características suficientemente distintivas como para garantizar su propia identidad.
Materiales y Métodos
Seguimos la terminología y formato de descripción empleados por Harris (1994). Las medidas se tomaron con un calibrador (precisión ± 0.1 mm) bajo una lupa de disección; las medidas tomadas son las siguientes: longitud rostro-cloacal (desde la punta del hocico hasta el labio anterior de la cloaca, LRC), longitud hocico-axila (desde la punta del hocico hasta por detrás de la inserción de la extremidad anterior, LHA), longitud axila-ingle (desde la axila hasta la ingle de las extremidades del mismo lado, LAI), longitud cefálica (desde la punta del hocico hasta el borde anterior del oído, LCB), ancho cefálico (a nivel de la porción más ancha de la cabeza, ACB), longitud de la extremidad anterior (desde la axila hasta la punta del dedo más largo, sin incluir la uña, LEA), longitud de la extremidad posterior (desde la ingle hasta la punta del dedo más largo, sin incluir la uña, LEP). Para evaluar las diferencias en escamación, tamaño y proporciones entre sexos, empleamos un análisis t-de Student; para propósitos estadísticos, comparamos el número total de escamas en los caracteres con conteos bilaterales (suma de los conteos de izquierda y derecha). Las medidas se presentan en el Cuadro 1. El sexo se determinó por disección. Los ejemplares están catalogados en las siguientes colecciones: Estación Biológica de Rancho Grande (EBRG), Ministerio del Ambiente y de los Recursos Naturales Renovables, Maracay; Museo de Historia Natural La Salle (MHNLS), Caracas; Museo del Instituto de Zoología Agrícola, Universidad Central de Venezuela (MIZA-UCV), Maracay; Museum of Natural History, University of Kansas (KU), Lawrence, U.S.A.; Museum of Vertebrate Zoology (MVZ), University of California in Berkeley, U.S.A.; Museum of Zoology, University of Michigan (UMMZ), Ann Arbor, U.S.A. Los datos de las distintas especies de Euspondylus se tomaron de Boulenger (1901, 1911), Burt & Burt (1931), Lancini (1968), Oftedal (1974), Peters (1862), Peters & Donoso-Barros (1970) y Uzzell (1973).
Resultados
Euspondylus acutirostris (Peters 1862)
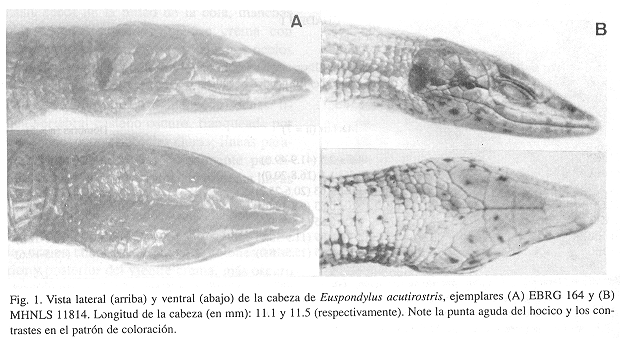
Figs. 1A-B
Descripción: Escamas prefrontales en amplio contacto por la sutura media; supraoculares comúnmente cuatro, rara vez tres; escamas supraciliares normalmente cinco, rara vez cuatro; serie de escamas supraciliares separan completamente las supraoculares de las ciliares; escama interparietal con los bordes laterales casi paralelos, divergiendo ligeramente hacia el extremo posterior; todas las escamas del dorso de la cabeza entre lisas y ligeramente rugosas, sin crestas ni protuberancias; escamas temporales 16-20 (izquierda, 17-21 (derecha); escamas occipitales grandes, pentagonales, situadas paramedialmente; segunda fila postparietal con escamas cuadradas, escamas centrales rectangulares, aproximadamente 1.5 veces más largas que anchas; escama nasal entera, entre 1.5 y 2.5 veces más ancha que alta, estrecha anteriormente, casi triangular; narinas redondeadas, situadas en el centro de la escama nasal; abertura nasal dirigida latero-posteriormente, canal descendente; escama loreal en amplio contacto con segunda supralabial; concavidad ciliar con o sin una fila de diminutas escamas planas; infralabiales agrandadas 5 (6 en EBRG 165); una geneial anterior seguida de 4 pares agrandados; par anterior de geneiales en amplio contacto por la sutura media, otras geneiales separadas; escamas sublabiales bordean la quinta (o sexta) infralabial y cuarta geneial; filas transversales completas de escamas pregulares 1-6, a continuación de 2-4 filas parciales; escamas pregulares no agrandadas, situadas oblicuamente; pliegue gular (de oído a oído) no evidente, excepto cuando la cabeza es empujada hacia abajo las escamas gulares se curvan convexamente hacia afuera mientras las escamas pregulares se mantienen planas; filas transversas de escamas gulares (incluyendo el collar) 6-21; serie paramedial de escamas gulares agrandadas ausentes; escamas del collar 7-10, escamas centrales más estrechas; escamas laterales del cuello aproximadamente 1/4 del tamaño de las escamas de la nuca; pliegue del collar difuso pero obvio, sin gránulos; pliegue gutural débil pero obvio.
Escamas alrededor del medio del cuerpo (a nivel de la fila media de escamas ventrales) 34-40; fila longitudinal de escamas dorsales 12-15; filas laterales 5-8 (izquierda), 5-9 (derecha); filas longitudinales de escamas ventrales 10-11; escamas laterales del cuerpo notablemente más pequeñas que las escamas ventrales y dorsales; filas transversales de escamas dorsales (desde las escamas occipitales hasta el margen posterior de las extremidades posteriores) 36-45; filas transversales de escamas ventrales (desde el pliegue del collar hasta la placa precloacal 21-28, o 23-30 hasta el labio anterior de la cloaca; escamas dorsales aproximadamente entre 1.6 y 2.0 veces más largas que anchas; escamas dorsales con una quilla central, recta, estrecha, con crestas o rugosidades laterales divergentes, normalmente algo difusas; escamas dorsales esencialmente cuadradas con el margen posterior ligeramente convexo, subimbricadas, con filas alternas; escamas medioventrales cuadradas, progresivamente más estrechas hacia los lados, normalmente subimbricadas, rara vez yuxtapuestas, lisas; borde posterior de las escamas ventrales recto; esquinas posteriores de las escamas ventrales angulares; escamas preanales 5-8 en las hembras, 5 en los machos; escamas redondeadas sobre el labio posterior de la cloaca; escamas supracaudales truncadas posteriormente; escamas subcaudales con margen posterior recto.
Escamas sobre las extremidades rugosas; escamas supradigitales del IV dedo manual 9-11; escamas infradigitales del IV dedo manual 13-17 (izquierda), 12-16 (derecha), algunas escamas proximales ligeramente tuberosas; escamas infradigitales mezcladas entre sencillas y divididas centralmente; escamas bajo el pólex de las manos 5-7; escamas de las palmas de manos y pies tuberosas, ninguna cornificada o agrandadas; escamas supradigitales de IV dedo pedal 10-13; escamas infradigitales del IV dedo pedal 17-22 (derecha), 15-23 (izquierda); escamas infradigitales mezcladas entre escamas sencillas y divididas centralmente; uñas largas, aproximadamente tan grande como la escama adyacente; extremo distal de los dígitos ligeramente comprimidos lateralmente; poros situados en escamas enteras; poros precloacales ausentes; poros femorales en machos 9-11 (derecha), 8-11 (izquierda); poros femorales en hembras 3-9 (derecha), 4-8 (izquierda); superposición de las extremidades hacia el centro del cuerpo entre -6 y +3 escamas; dígitos en orden decreciente de extensión : 4 > 3 2 = 5 > 1.
Melanización peritoneal presente; superficie dorsal de la lengua cubierta de filas oblicuas de papilas en forma de escamas; pliegues estrechos infralinguales, oblicuos; dientes bi o tricuspidados, unos pocos dientes frontales cónicos, 15-21 a lo largo de la mandíbula superior, 14-20 a lo largo de la mandíbula inferior; paladar hendido en el centro.

Coloración en preservativo: Dorso castaño con diminuto punteado oscuro anteriormente, progresivamente más grandes hacia el extremo final del cuerpo; puntos grandes y numerosos sobre 1/3 anterior de la cola; línea dorsolateral oscura presente (ligeramente más oscura que el dorso) desde el borde posterior del ojo, recorre por encima de los hombros cubriendo completamente los flancos, terminando en la ingle; ocelos crema blancuzcos, bordeados de negro, 8(izquierda)/8(derecha) en EBRG 159, 5/4 en EBRG 164, 1/1 en MHNLS 12675; ocelos redondeados, situados en la parte baja en los flancos; ocelos ausentes en MIZA-UCV 15-026, EBRG 165 y 840; región ventral posterior gris oscuro, anteriormente crema amarilloso pálido (incluyendo la garganta, mentón y superficie ventral de las extremidades y cola). Cabeza castaña, ligeramente más oscura que el dorso, sin marcas.
MHNLS 11943 y MVZ 116532 exhiben una línea dorsolateral prominente, blanca y gris, respectivamente, con bordes negros, bien definidos, entre el margen posterior del ojo; las líneas se fragmentan un poco por detrás de los hombros; algo anterior a la ingle y hasta la porción anterior de la cola se observaron numerosas manchas claro-oscuras, dispersas. MHNLS 11943 y MVZ 116532 presentan 6/8 y 5/5 ocelos blancos con bordes negros sobre la parte inferior de los flancos, respectivamente; vientre entre crema blancuzco y crema amarilloso con punteado oscuro en el centro de cada escama. El ejemplar MHNLS 11942 es muy similar a MHNLS 11943, excepto que las líneas blancas bordeadas de negro dorsolaterales se observan bien definidas a lo largo de todo el cuerpo desde el borde anterior del ojo hasta cerca de la mitad de la cola, manchas claro-oscuras ausentes, vientre crema con punteado oscuro inconspícuo, algunos ocelos pequeños sobre el codo, ingle y rodilla.
Ejemplar EBRG 3248 muestra el dorso con cinco líneas longitudinales, estrechas: línea vertebral castaño oscuro, flanqueada por dos líneas paravertebrales claras; líneas paravertebrales flanqueadas lateralmente por líneas oscuras; dorsolateralmente presenta líneas claras; cerca de ½ del cuerpo todas las líneas se fragmentan en grandes manchas claro-oscuras a lo largo del resto del dorso hasta la cola; ocelos laterales ausentes; porciones anterior y posterior del vientre crema, más oscuro en el centro; extremidades dorsalmente castaño oscuro con ocelos crema con bordes negros; dígitos dorsalmente bicoloreados, alternando escamas claras y oscuras; una línea oscura sobre otra blanca cremosa, más gruesa, desde el borde posterior del ojo hasta el borde anterior del oído; una línea clara bordeada por dos líneas oscuras entre el borde posterior del oído hasta algo por delante de los hombros, línea oscura inferior algo difusa anteriormente, fragmentada, progresivamente deviene más definida posteriormente.
En el ejemplar MHNLS 11814 dorso castaño grisáceo pálido; flancos castaño, más pálido que el dorso, con 6/5 ocelos, claros, diminutos, algo difusos; extremidades igual a los flancos, excepto por presentar ocelos claros con bordes oscuros; vientre blanco cremoso con una mancha oscura en el centro de cada escama; escamas supralabiales crema; dos líneas, una oscura y estrecha por encima de otra ligeramente más ancha, blanca cremosa, entre el borde posterior del ojo y el borde anterior del oído; entre el borde posterior del oído y algo por delante de los hombros una línea blanca ancha es bordeada por dos líneas oscuras, estrechas.
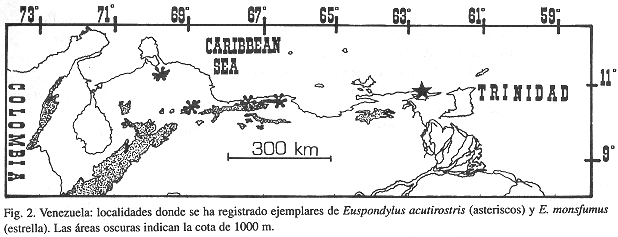
Distribución y ecología: Los ejemplares estudiados provienen de localidades situadas entre 1093 y 2760 m.s.n.m., habitando una variedad de zonas de vida que van desde Bosque Ombrófilo Montano (Bosque Nublado Costero) hasta el Matorral Subparamero Costero según Huber & Alarcón (1988). Esta especie parece estar restringida a elevaciones por encima de 1000 m., desde la porción central de la Cordillera de la Costa en el centro norte del país, la Sierra de Aroa y la Sierra de San Luis en la región noroccidental de Venezuela (ver Fig. 2).
Ocho ejemplares disponían de datos sobre microhábitat: tres ejemplares (EBRG 159, 165 y 3248) fueron encontrados dentro de bromelias aéreas, un ejemplar (MHNLS 11814) fue recolectado entre la hojarasca del suelo dentro del bosque primario (Elías León, comunicación personal, 1995) y otros cuatro se recolectaron dentro o en los alrededores del edificio de la Estación Biológica de Rancho Grande (EBRG 164, 840, 841, MIZA-UCV 15-026, rodeado por bosque primario.
Todas las hembras tenían huevos en los oviductos, cuatro de ellas con uno o dos huevos, maduros, grandes, de color crema anaranjado pálido; sólo un ejemplar contiene varios huevos blancos, pequeños e inmaduros. Las hembras con huevos maduros fueron recolectadas entre los meses de agosto y noviembre; la única hembra con huevos inmaduros fue recolectada en abril. En el Parque Nacional Henri Pittier (porción central de la Cordillera de la Costa) la temporada lluviosa ocurre entre los meses de mayo y diciembre (Huber 1986), al igual que en la Sierra de San Luis, Falcón (Pla et al. 1978). Esta coincidencia sugiere que la oviposición en Euspondylus acutirostris ocurriría al final de la estación lluviosa.
Dimorfismo sexual: Encontramos dimorfismo sexual en al menos ocho caracteres (ver Cuadro 2): tres de escamación (número de escamas precloacales, número de filas transversales de escamas dorsales y número de poros femorales) y cinco de tamaño (longitud rostro-cloacal, longitud axila-ingle, relación longitud cefálica/longitud rostro-cloacal, relación longitud cefálica/longitud axila-ingle, relación ancho cefálico/longitud rostro-cloacal).
No hay diferencias apreciables en la coloración corporal entre ambos sexos. Sin embargo, la mayoría de los machos (excepto EBRG 840, 841) y una sola hembra (MHNLS 11814) presentan ocelos en los flancos.
Aunque el sexo del holotipo no se define en la descripción original, creemos que este es un macho por tener 42 filas transversales de escamas dorsales, cinco escamas precloacales, 10-11 poros femorales y se ajusta bastante bien a las restantes proporciones del cuerpo y cabeza determinadas como dimórficas (ver Cuadro 2).
Comentarios: El segundo ejemplar conocido de Euspondylus acutirostris fue registrado por Test et al. (1966). Este ejemplar (UMMZ 125769) ya no existe; posiblemente fue destruido durante el proceso de transparentación y teñido para estudio del esqueleto (Greg Schneider in litt., 20 de julio de 1994). Lancini (1968) sólo resumió los datos de E. acutirostris de fuentes bibliográficas, pero Uzzell (1973) lo citó como si hubiese registrado ejemplares adicionales.
Test et al. (1966) sugieren que Euspondylus acutirostris habita en la hojarasca epífita (la que queda retenida entre las ramas de los árboles); Uzzell (1973) destaca que la estructura de las escamas infradigitales es similar a las de las especies arbóreas de Anadia. Esta arborealidad parece confirmarse en que tres individuos se obtuvieron de bromelias, sin embargo, cinco fueron encontrados en el suelo dentro del bosque primario o en construcciones humanas lo cual sugiere que E. acutirostris emplea indistintamente microhábitats verticales y horizontales.

Euspondylus acutirostris habita en el bosque nublado entre los 1093 y 2760 metros de altura, entonces, los ejemplares disponibles probablemente representen tres poblaciones separadas. La población presumiblemente nominal, la de la porción central de la Cordillera de la Costa, es la mejor representada. Las poblaciones de la Sierra de San Luis (estado Falcón) y de la Sierra de Aroa (estado Yaracuy), referidas aquí a E. acutirostris, cada una representada por un único individuo, muestran algunas diferencias. El macho de la Sierra de Aroa tiene varias listas mediodorsales y dorsolaterales claro-oscuras, una línea postocular clara con bordes negros y ocelos claros con bordes negros en las extremidades. La hembra de la Sierra de San Luis tiene una línea postocular clara con bordes negros en las extremidades, ocelos claros sin bordes oscuros en los flancos, listas dorsales y dorsolaterales ausentes y un bajo número de dientes en la mandíbula superior (15) y dentario (16). Serían necesarios ejemplares adicionales de estas poblaciones para establecer si ellas requieren un reconocimiento taxonómico distinto.
Euspondylus monsfumus sp. nov.
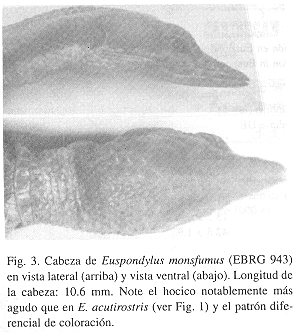
Fig. 3
Holotipo: EBRG 943, hembra, del Cerro El Humo, estado Sucre, Venezuela, aproximadamente 800 m. (?), (aprox. 10°42N, 62°37W), recolectado por S. Gorzula, el 15 de abril, 1978.
Diagnosis: El hocico extremadamente agudo de esta especie, 24° en vista dorsal, es su caracter más distintivo. Euspondylus acutirostris (30°-36°) y E. guentheri (35°, fide Harris, in litt. 1999) son las especies que más se acercan a esta condición. Las otras especies asignadas a Euspondylus tienen hocicos más cortos y obtusos. El elevado número de filas de escamas ventrales transversales distingue E. monsfumus (28) y E. acutirostris (26-33) de las otras especies de Euspondylus (19-24, colectivamente). Cuatro especies, E. monsfumus, E. acutirostris, E. guentheri y E. rahmi, poseen cuatro escamas supraoculares sobre cada ojo. Adicionalmente, E. monsfumus se distingue de E. acutirostris por el menor número de escamas temporales (16-22, contra 24-29 por lado), escamas dorsales de las extremidades claramente quilladas contra rugosas, escama nasal rectangular contra triangular, respectivamente.
Descripción: Escamas prefrontales en amplio contacto por sutura media; escamas supraoculares 4; escamas supraciliares 5 por cada lado; serie supraciliar completamente separa las escamas supraoculares de las escamas ciliares; escama interparietal con los bordes laterales casi paralelos, divergiendo ligeramente hacia el extremo posterior; todas las escamas del dorso de la cabeza rugosas, sin crestas o protuberancias; escamas temporales 29 (izquierda), 24 (derecha); escamas occipitales grandes, pentagonales, paramediales; segunda fila postparietal no agrandadas transversalmente, escama central rectangular, aproximadamente 1.5 veces más larga que ancha; escama nasal entera, 2.5 veces más ancha que alta, rectangular; narinas redondeadas, situada en el centro de la nasal; abertura narinal dirigida latero-posteriormente, canal descendiente; escama loreal en amplio contacto con segunda escama supralabial; concavidad ciliar con una fila de escamas diminutas, planas; palpebrales superiores no evidentes; palpebrales inferiores 3 por lado; palpebrales inferiores translúcidas, difusamente manchadas; una fila de gránulos bajo las palpebrales inferiores; 4 escamas suboculares en contacto con labiales; una fila de gránulos en el borde anterior de la abertura auricular; gránulos preauriculares 6 (izquierda), 8 (derecha), anchos; escamas supralabiales 7 (5 a nivel de la inflexión del labio), tan largas como anchas; infralabiales agrandadas 5; serie de geneiales dispuestas en una anterior entera seguida de 4 pares; par anterior de geneiales en completo contacto por sutura media, las restantes separadas; escamas sublabiales bordean quinta infralabial y cuarta geneial; filas transversas completas de pregulares 6, por detrás de 2 filas parciales; pregulares no agrandadas, dispuestas oblicuamente; pliegue gular no evidente, excepto cuando la cabeza se empuja hacia abajo que las escamas gulares se curvan convexamente hacia afuera mientras las pregulares se mantienen planas; fila transversal de gulares (incluyendo el collar) 14; serie paramedial de gulares agrandadas ausentes, gulares ½ tan largas como anchas, separadas por una escama central estrecha; escamas del collar 8, escamas centrales más estrechas; escamas laterales del cuello aproximadamente 1/3 del tamaño de las escamas de la nuca; pliegue del collar ligero pero obvio, no tiene gránulos; pliegue gutural débil pero obvio.
Escamas alrededor del medio del cuerpo (a nivel de la mitad de las filas transversales ventrales) 36; filas longitudinales de escamas dorsales 14; filas de escamas laterales 6 por lado; filas longitudinales de escamas ventrales 10; las escamas laterales son claramente más pequeñas que las escamas dorsales y ventrales; filas transversales de dorsales entre la escama occipital hasta el margen posterior 45; filas transversales de escamas ventrales 28 hasta la placa precloacal, 30 hasta el labio anterior de la cloaca; escamas dorsales aproximadamente 2.0 veces más largas que anchas; escamas dorsales con una quilla central, recta, estrecha, resto de la superficie de la escama lisa; dorsales esencialmente cuadradas con el borde posterior ligeramente convexo, subimbricadas, en filas alternadas; escamas medioventrales cuadradas, estrechándose progresivamente hacia los lados, normalmente subimbricadas, lisas; borde posterior de las escamas ventrales recto; esquinas posteriores de las escamas ventrales angulosas; escamas precloacales 7; escamas sobre el labio posterior de la cloaca redondeados; escamas supracaudales truncadas posteriormente; escamas subcaudales con borde posterior recto.
Escamas dorsales de las extremidades quilladas; escamas supradigitales de IV dedo manual 10 por lado; escamas infradigitales del IV dedo manual 17 (izquierda), 15 (derecha), con algunos tubérculos proximales; escamas infradigitales mezcladas entre escamas sencillas y divididas longitudinalmente por el centro; escamas bajo el pólex de la mano 6 (izquierda), 5 (derecha); escamas de las palmas manuales y pedales tuberosas, no cornificadas ni agrandadas; escamas supradigitales del IV dedo pedal 10 (izquierda), 11 (derecha); escamas infradigitales del IV dedo pedal 20 (izquierda), 21 (derecha); escamas infradigitales divididas longitudinalmente por el centro; uñas grandes, aproximadamente tan grandes como la escama adyacente; punta de los dígitos con una ligera compresión lateral; escamas poríferas enteras; poros femorales 9 (izquierda), 10 (derecha); extremidades estiradas hacia el centro del cuerpo con + 1 escama de superposición; dígitos en orden decreciente de longitud 4 > 3 > 2 = 5 > 1.
Melanización peritoneal ausente; superficie dorsal de la lengua cubierta de papilas en forma de escamas; pliegues infralinguales estrechos, oblicuos; dientes bicúspides, unos pocos al frente cónicos, 20 a lo largo de la mandíbula superior, 17 a lo largo de la mandíbula inferior; paladar hendido.
Medidas (en mm): SVL 46.0, SAL 19.1, AGL 24.1, HL 10.6, HW 5.6, FLL 11.2, HDL 16.6.
Distribución y ecología:Euspondylus monsfumus se conoce sólo para la localidad tipo en alturas intermedias del Cerro El Humo, nordeste de Venezuela (Fig. 2). Esta localidad se encuentra en la zona de vida descrita como Bosque Nublado Costero por Huber & Alarcón (1988). El ejemplar fue encontrado dentro de una bromelia arbórea (S. Gorzula, in litt. 1995). El holotipo es una hembra adulta, con dos huevos grandes en la cavidad abdominal, maduros, color crema.
Comentarios: Debido a las dificultades en hallar características que definan objetivamente al género Euspondylus, asignamos esta nueva especie a dicho género tentativamente como una cuestión de conveniencia; además, a primera vista, E. monsfumus recuerda a (y puede ser fácilmente confundido con) E. acutirostris.
Euspondylus monsfumus exhibe la punta del hocico mucho más puntiaguda (en vista dorsal y lateral) que Euspondylus acutirostris. Además, al examinar la mandíbula superior e inferior reveló la aparente ausencia de dientes en el área premaxilar. Es decir, no pudimos observar (bajo la lupa estereoscópica) o sentir (tocando con una aguja de disección) ningún diente o estructura similar en la punta de las mandíbulas. No podemos determinar si la pérdida de los dientes premaxilares fue después de la recolecta del ejemplar (e. g., por manejo descuidado), o si su ausencia es una condición natural. De comprobarse esto último, probablemente E. monsfumus tendría una dieta muy especializada.
Etimología: El epíteto específico monsfumus deriva del latín mons (= montaña) y fumus (=humo). Hace referencia al nombre de la localidad de donde proviene el holotipo, Cerro El Humo. Este es un nombre en aposición en el caso genitivo singular.
Agradecimientos
Gracias a E. León (FUDENA), L. F. Navarrete (MCNC) y G. Schneider (UMMZ) por la valiosa información acerca de algunos ejemplares de Euspondylus acutirostris. También, agradecemos a Stefan Gorzula y Robert Wingfield, Richard A. Siegel y un árbitro anónimo por revisar críticamente una versión preliminar del manuscrito. Tenemos una gran deuda con Dennis M. Harris quien revisó varias versiones del manuscrito haciéndole valiosas sugerencias y extensas modificaciones, generosamente compartió bibliografía, numerosos datos inéditos y figuras del ejemplar MVZ 116532. Francisco Bisbal y Ramón Rivero (EBRG), John Simmons (KU) y Jesús Manzanilla (MIZA-UCV), amablemente prestaron ejemplares depositados en las colecciones bajo su cuidado. Este trabajo es parte del proyecto S1-9112-030, financiado parcialmente por Fundacite-Falcón (para AAR y AMU); igualmente, se presentó en forma de cartel en la XIII Reunión de Comunicaciones Herpetológicas, Santa Fe, Argentina, gracias a la subvención del Consejo Nacional de Investigaciones Científicas y Tecnológicas (expediente N° S2-98001514 para AMU).
Referencias
Boulenger, G. A. 1901. Further descriptions of new reptiles collected by Mr. P. O. Simons in Peru and Bolivia. Ann. Mag. Nat. Hist. VII(42): 546-549. [ Links ]
Boulenger, G. A. 1911. Descriptions of new reptiles from the Andes of South America, preserved in the British Museum. Ann. Mag. Nat. Hist. VII(37): 19-25. [ Links ]
Burt, C. E. & Burt, M. D. 1931. South American lizards in the collection of the American Museum of Natural History. Bull. Amer. Mus. Nat. Hist. 61: 227-395. [ Links ]
Gorzula, S. & Señaris, J. C. 1998. Contribution to the herpetofauna of the Venezuelan Guayana. I. A data base. Scientia Guayanae 8: 1-270. [ Links ]
Harris, D. M. 1994. Review of the teiid lizard genus Ptychoglossus. Herp Monogr. 8: 226-275. [ Links ]
Huber, O. 1986. El clima, p. 17-29. In O. Huber (ed.). La Selva Nublada de Rancho Grande Parque Nacional Henri Pittier, El Ambiente Físico, Ecología Vegetal y Anatomía Vegetal. Fondo Editorial Acta Científica Venezolana, Caracas. [ Links ]
Huber, O. & Alarcón, C. 1988. Mapa de Vegetación de Venezuela. IUCN-Ministerio del Ambiente y los Recursos Naturales Renovables,.Armitano, Caracas. [ Links ]
La Marca, E. 1997. Lista actualizada de los reptiles de Venezuela, p. 123-142. En: E. La Marca (ed.). Vertebrados Actuales y Fósiles de Venezuela. Serie Catálogo de Vertebrados de Venezuela, Vol. 1. Museo de Ciencias y Tecnología de Mérida, Venezuela. [ Links ]
La Marca, E. & García-Pérez, J. E. 1990. Prionodactylus ampuedai (Sauria: Teiidae): A new lizard record for Colombia, with taxonomic comments on the species. Bull. Maryland Herpetol. Soc. 26: 111-115. [ Links ]
Lancini, A. R. 1968. El género Euspondylus (Sauria: Teiidae) en Venezuela. Publ. Ocas. Mus. Cienc. Nat. Caracas, Zool. 12: 1-6. [ Links ]
Myers, C. W. & Donnelly, M. A. 1996. A new herpetofauna from Cerro Yaví, Venezuela: First results of the Robert G. Goelet American Museum-Terramar Expedition to the northwestern Tepuis. Amer. Mus. Novitates 3172: 1-56. [ Links ]
Oftedal, O. T. 1974. A revision of the genus Anadia (Sauria: Teiidae). Arq. Zool., S. Paulo 25: 203-265. [ Links ]
Peters, J. A. & Donoso-Barros, R. 1970. Catalogue of the neotropical squamata. Part II. Lizards and amphisbaenians. U. S. Natnl. Mus. Bull. 297: 1-293. [ Links ]
Peters, W. 1862. Über Cercosaura und die mit dieser gattung erwandten eidechsen aus Südamerika. Phys. Abh. der K. Ak. d. Wiss. 1862: 164-225. [ Links ]
Pla, L., Matteucci, S. & Colma, A. 1978. El clima de Falcón. Análisis regional de la vegetación y el ambiente del estado Falcón. Instituto Universitario de Tecnología de Coro, Ofic. Reproducción, Coro. 64 p. [ Links ]
Test, F. H., Sexton, O. J. & Heatwole, H. 1966. Reptiles of Rancho rande and vecinity, Estado Aragua, Venezuela. Misc. Publ. Mus. Zool. Univ. Michigan 128: 1-63. [ Links ]
Uzzell, T. 1973. A revision of lizards of the genus Prionodactylus, with a new genus for P. leucostictus and notes on the genus Euspondylus (Sauria, Teiidae). Postilla 159: 1-67. [ Links ]
Apéndices
Ejemplares examinados: Todos los ejemplares de la siguiente lista provienen de localidades en Venezuela. El ejemplar con un "?" no fue posible confirmar su sexo.
Euspondylus acutirostris: ARAGUA MHNLS 8652, hembra, Estación Biológica de Rancho Grande; MIZA-UCV 15-016, hembra, Parque Nacional Henri Pittier, cerca de la Estación Biológica de Rancho Grande, 1100 m EBRG 840-41, 164, machos, Estación Biológica de Rancho Grande (edif.); EBRG 159, 165 (machos), 171 (hembras), El Portachuelo, Rancho Grande; MVZ 116532, macho, Rancho Grande, aproximadamente 1100m.- DISTRITO FEDERAL: MHNLS 11942 (hembra), MHNLS 11943 (macho), Pico Niguatá, Parque Nacional El Avila, 2750 m.- FALCÓN: MHNLS 11814 ,hembra, Parque Nacional Juan Crisóstomo Falcón, Cerro El Danto (Fila de Úrea), Sierra de San Luis, 1400m.- MIRANDA: MHNLS 5997, hembra (?), Pico Naiguatá, El Anfiteatro, 2760 msnm; MHNLS 12678, hembra, Pico Naiguatá, Parque Nacional El Avila.-YARACUY: EBRG 3248, macho, Los Bacos, Sierra de Aroa, 1600m.
1 Centro de Investigaciones en Ecología y Zonas Áridas (CIEZA), Universidad Francisco de Miranda, Apartado 7559, Santa Ana de Coro 4101-A, Venezuela. Fax: +58-68-524922.
Correo electrónico: amijares@funflc.org.ve / aarends@funflc.org.ve
2 Museo de Historia Natural La Salle, Apartado 1930, Caracas 1010-A, Venezuela. Fax: +58-2-7937493. Correo electrónico: celsa.s@usa.net

