










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.48 n.1 San José Mar. 2000
Quintana Roo, México
José Rolando Bastida-Zavala Aurora U. Beltrán-Torres Martha Angélica Gutiérrez-Aguirre y Gabriela de la Fuente-Betancourt 1
Abstract
The Majagual patch reef was surveyed 13 days after the indirect effects of hurricane Mitch with 1 m2 quadrants and rapid assesment techniques. In comparison with an earlier survey in the Xahuayxol reef, live coral cover was low (11.5% ± 5.2), whereas the cover of filamentous seaweed and old dead coral was high (53.7% ± 27.7 in bath of them). The coral community disturbance may have began several years ago, and low algal turf growth suggests indirect effects of hurricane Mitch, because the surge suspended sediments. Frequency of Favia fragum (Esper) and encrusting coralline algae encounters was higher than for other coral/alga pairs; this suggest a possible effect specificity.
Key words
Benthos, hurricane Mitch, patch reefs, rapid assessment, tourism.
Los arrecifes coralinos del Atlántico presentan un fenómeno en común: se están transformando en arrecifes algales (Lapointe 1989); empero, aún se mantiene la discusión de los agentes causales. Se esgrimen múltiples explicaciones para ello; entre las más importantes se encuentra la drástica disminución, en tiempos recientes (ca. 500 años), de los macroherbívoros, causadas por la sobre-explotación histórica de estos recursos o por mortalidades masivas de origen desconocido (Jackson 1997). Igualmente, ha influido el aumento de las descargas de sedimentos y materia orgánica en los sitios costeros utilizados por el hombre, que ha elevado la cantidad de nutrimentos disponibles para las algas, como en los cayos de Florida, Jamaica, Veracruz, Cancún (Lang et al. 1998). Otras causas que se esgrimen, aunque con menor evidencia, incluyen al cambio climático global y la elevación del nivel medio del mar, aunque otros autores consideran que algunos de estos eventos causarían a los arrecifes coralinos más beneficios que perjuicios (Grigg 1992).
Aún no se tiene un extenso conocimiento de los ecosistemas arrecifales de Quintana Roo y ya existe un fuerte deterioro en muchos de ellos (Lang et al. 1998). El estado estaba prácticamente despoblador hace apenas un siglo; para 1950 había un reducido número de poblaciones costeras (entre las que se encontraba Majagual), y apenas hace 20 años surgió un enorme desarrollo turístico en Cancún, que se ha extendido a la región norte del Caribe mexicano, conocida actualmente como Riviera Maya. En los últimos años, la inversión en infraestructura turística se ha ido incrementado en el sur del estado.
Majagual se encuentra en un acelerado proceso de transformación desde hace por lo menos diez años. Actualmente el arrecife frente a esta localidad, que forma parte de la Barrera Arrecifal Mesoamericana, representa un atractivo menor a los visitantes del que pudo haber tenido hace 10-12 años, como ha sido constatado por uno de nosotros (Beltrán-Torres). Incluso, es posible que lo que actualmente existe en el arrecife sea una caricatura de lo que fue hace algunos cientos de años (sensu Jackson 1997). Como respuesta al imparable desarrollo humano han surgido diversas técnicas de evaluación rápida, que permiten obtener cierta estimación del estado de salud de los ecosistemas arrecifales; sin embargo, no debe perderse de vista que los estudios de largo plazo proporcionan mayor información y predictibilidad (Aronson et al. 1994). El objetivo del presente estudio es evaluar con técnicas rápidas a los arrecifes parche de Majagual.
Materiales y Métodos
Majagual se localiza al sur de Quintana Roo, 18º44N y 87º42W. Es un pequeño poblado con 500 habitantes permanentes y que recibe afluencia turística con regularidad. Existen varios hoteles y cabañas a lo largo de la costa, incluyendo una instalación militar y varios comercios. No existe tratamiento alguno para las aguas residuales generadas por esta población.
La costa presenta una playa angosta y un humedal compuesto de manglares, con dominancia local de Rhizophora mangle, el cual se extiende a lo largo de la costa caribeña de México. El arrecife coralino se encuentra muy cercano a la costa (ca. 300 m), su laguna arrecifal es somera (1-2 m de profundidad), con lechos de pastos marinos (Thalassia testudinum y Syringodium filiforme) y amplias zonas arenosas donde se ubican varios arrecifes parche. El antearrecife y la cresta arrecifal albergan la mayor diversidad del sitio, mientras que el arrecife frontal está poco desarrollado en comparación con otras áreas del Caribe (Jordan 1979, Tunnell et al. 1993).
El trabajo de campo se realizó el 10 de noviembre de 1998, 13 días después de que la costa quintanarroense recibió los efectos indirectos del huracán Mitch. Se observó gran cantidad de pastos marinos y algas que habían sido arrojados a las playas por el fuerte oleaje, así como signos de erosión en algunos sitios de la playa. Por otro lado, se observaron manchas de hidrocarburos en la superficie del mar cuyo origen no se pudo precisar.
En dos arrecifes parche someros se colocaron cinco cuadrantes de PVC de 1 m2 divididos en cuatro subcuadrantes, manteniendo una distancia mínima de 1 m entre ellos. En cada cuadrante se estimó la cobertura de coral vivo, recientemente muerto y antiguamente muerto, algas, esponjas y otros invertebrados. La identificación de los organismos se hizo in situ. Se consideraron los tres grupos funcionales de algas (filamentosas, arborescentes e incrustantes) establecidos por Littler et al. (1983) y Steneck y Dethier (1994) y se registró la altura promedio del tapete algal. Para evaluar la complejidad estructural de los arrecifes parche se usó el índice de complejidad topográfica (C), que consiste en tomar dos distancias entre los vértices opuestos de los cuadrantes, siguiendo el contorno del sustrato; se calculó a partir de la fórmula:
donde:d= distancia rectilínea entre ambos vértices,
Se asume que los disturbios que producen una mortalidad completa o parcial disminuyen el índice de complejidad (Aronson et al. 1994, Aronson y Precht 1995). Asimismo, se utilizó el análisis de la vitalidad coralina, comparando la cobertura de coral vivo con la de coral reciente y antiguamente muerto (Ginsburg et al. 1996, Steneck et al. 1997). Los datos obtenidos se compararon con los resultados gráficos de un trabajo previo realizado en Xahuayxol (18º30N y 87º45W), sitio ubicado 20 km al sur de Majagual (Carricart-Ganivet et al. en prep.) y que carece de todo tipo de desarrollo.
Por último, se realizó un censo visual de los encuentros entre algas y corales, presumiendo competencia entre ambos taxa. El censo se realizó sobre los parches coralinos en un área aproximada de 2 500 m2. El esfuerzo total fue de 1.5 horas/hombre, tomando nota de las especies de corales y algas que se encontraron en relación estrecha, asumiendo ésta como el contacto directo entre alga y coral o los contactos eventuales cuando el talo está en contacto con el coral debido al movimiento provocado por el oleaje. La desviación estándar se abrevió como ± y la notación de aproximadamente como ~.
Resultados
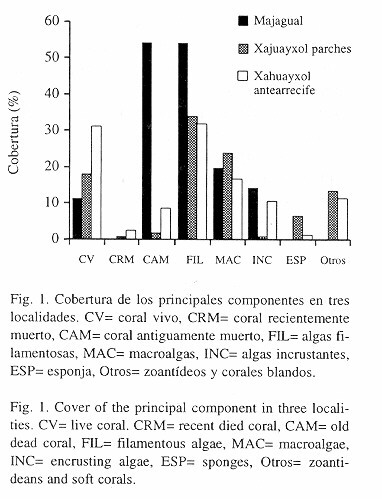
En Majagual la mayor cobertura la ocupó el coral antiguamente muerto (53.7% ±27.7), el cual se encontró cubierto por algas filamentosas (Fig. 1). Las diferencias entre Majagual y Xahuayxol son notables, tanto en cobertura del coral vivo (11.5% ±5.2 vs. ~18%), como la del coral antiguamente muerto (53.7% vs. ~2%), además de que en Majagual no se observó coral recientemente muerto y en Xahuayxol sí (Fig. 1).
Por su parte, la cobertura de algas filamentosas es mucho mayor en Majagual (53.7% ±27.7) que en cualquiera de los dos sitios de Xahuayxol (Fig. 1). Las macroalgas presentan una cobertura similar en todos los sitios (Majagual: 20.3% ±8.2), mientras que las algas incrustantes presentan mayor parecido entre Majagual (14.4% ±20.9) y el antearrecife de Xahuayxol (Fig. 1). La cobertura por otros invertebrados es mínima en Majagual mientras que en Xahuayxol cubren más del 10% (Fig. 1).
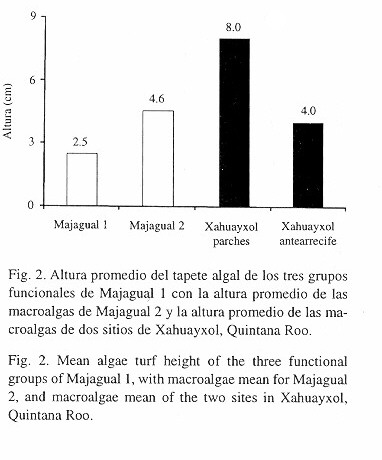
En Majagual 1 la altura promedio de las algas es menor (2.7 cm ±2.9) que en Xahuayxol (>4 cm), aunque para Majagual 1 incluye la altura de los tres grupos de algas (Fig. 2); mientras que en el trabajo de Carricart-Ganivet et al. (en prep.), la altura del tapete algal se refiere sólo a la máxima altura de las macroalgas. Para Majagual 2 sólo se tomó en cuenta la altura promedio de las macroalgas; de esta forma la altura de las macroalgas en Majagual 2 (4.6 cm ±1.1) se asemeja más al del antearrecife de Xahuayxol (~4 cm) que a los arrecifes parche (~8 cm) de ese mismo sitio (Fig. 2).
Por su parte, el valor del índice de complejidad topográfica para Majagual fue de 0.23 (±0.1), un valor muy cercano al 0.27 obtenido por Carricart-Ganivet et al. (en prep.) para Xahuayxol.
Los corales registrados en los cuadrantes de Majagual incluyen siete especies, dominando principalmente Montastrea annularis y Porites astreoides; salvo un cuadrante, donde se presentaron cuatro especies de corales, ningún otro tuvo más de 2-3 especies. Mientras tanto, las algas observadas en los cuadrantes y en el censo totalizan 11 especies, siete rodofitas (más las incrustantes no identificadas) y cuatro clorofitas (Cuadro 1).
Los encuentros alga/coral suman 31, siendo el encuentro más frecuente el del coral Favia fragum y las algas coralinas incrustantes (nueve encuentros), seguida por M. annularis y Centroceras sp. (tres encuentros). El resto de las asociaciones fueron registradas sólo una o dos veces (Cuadro 1).
Lista de algas, invertebrados y asociaciones coral/alga registrados en los arrecifes parche de Majagual, Quintana Roo
| Algas: | Corales: |
| Chlorophyta | |
| | |
| | |
| | |
| | |
| Rhodophyta | |
| | |
| | |
| | |
| | |
| | |
| | |
| | |
| | |
| Relación coral/alga (s) | |
| | |
| | |
| | |
| | |
| | |
| | |
| | |
| | |
| | |
| | |
| | |
| | |
| | |
| | |
| | |
| | |
| | |
| | |
| | |
| | |
| | |
| | |
Discusión
La altura de las macroalgas en Majagual no parece representar un buen indicador de disturbio al compararlo con los datos de Xahuayxol. Por su parte, la reducida altura promedio del tapete algal, con los tres grupos de algas incluidos, puede representar un alto ramoneo o herbivoría; sin embargo, los organismos herbívoros estaban prácticamente ausentes, salvo tres erizos registrados en dos de los cuadrantes (Echinometra? sp.) y muy pocos peces herbívoros observados, entre los que se encontraba un acantúrido y varios pomacéntridos (observaciones in situ).
Por el contrario, los pomacéntridos también promueven la dominancia de algas filamentosas y aumentan la mortalidad en los corales e, indirectamente, incrementan la vulnerabilidad del sustrato coralino debida a la bioerosión ocasionada por los invertebrados perforadores (e.g. Cliona spp., sipuncúlidos) que están estrechamente relacionados con los tapetes algales (Risk y Sammarco 1982). Sin embargo, harían falta censos estacionales de los organismos herbívoros para comprobarlo.
Al parecer existe cierta especificidad de F. fragum y las algas coralinas incrustantes, indicado por la elevada frecuencia de los encuentros. Esto puede ser explicado por las condiciones del hábitat de los arrecifes parche someros y las características propias de ambos organismos, ya que ambos tienen preferencia por las zonas someras de los parches arrecifales, en lugares de alta energía mecánica provocada por el oleaje. Se ha afirmado que las macroalgas tienen un gran potencial de impacto sobre las comunidades de corales escleractíneos, además de que los encuentros entre ambos grupos se dan aún a niveles de muy baja cobertura algal (Tanner 1995). El crecimiento del alga sobre el coral produce necrosis alelopática de los tejidos del coral; las algas también compiten en manera pasiva, acaparando todo el espacio disponible que pudiera servir para el crecimiento del coral o el asentamiento de sus larvas (Tanner 1995). El registro estandarizado de los encuentros coral/alga puede conformar un enfoque nuevo hacia la comprensión del nivel de especificidad en la competencia entre ambos taxa.
Como en Majagual no se encontró coral recientemente muerto se considera que el sitio ya había sido afectado por otros disturbios anteriores al huracán Mitch y, al parecer, el fuerte oleaje ocasionado por este meteoro no incrementó la mortalidad del ya de por sí escaso coral vivo (Beltrán-Torres observ.). Aunque no se evaluó directamente el efecto del turismo sobre el arrecife, se sospecha que la deforestación del manglar y el turismo ha incrementado la erosión de las playas y afectado los parches coralinos. El balance de los sedimentos en las dunas es muy delicado y, ante la falta de información previa comparativa, se recomienda a los tomadores de decisiones que apliquen el principio de precaución, estimulando la investigación en ese sitio para poder contar con un marco de referencia más preciso del impacto humano y de la capacidad de carga del lugar (Salazar-Vallejo y González 1993).
En conclusión, la extensa cobertura de coral antiguamente muerto nos indica un deterioro del arrecife que tiene ya varios años de haber comenzado (<12 años), mientras que la alta cobertura de algas filamentosas puede ser un indicador del enriquecimiento de nutrimentos producido por contaminación orgánica y/o resuspensión de sedimentos. La reducida altura del tapete algal puede indicar, por un lado, alto herbivorismo, o bien un fuerte disturbio físico (como el provocado por el huracán Mitch). Esto contrasta con la baja resolución de la cobertura de coral recientemente muerto para predecir eventos de corto plazo, al menos en Majagual. Un monitoreo de la cobertura coralina en los arrecifes parche, incluyendo videotransectos, cuadrantes permanentes, análisis de tasas de bioerosión y crecimiento del coral, que permita evaluaciones oportunas antes y después de fenómenos naturales y antropogénicos, arrojaría más luz sobre los patrones de disturbio y las respuestas de largo plazo que tienen los arrecifes parche hacia estos disturbios.
Resumen
Se realizó un estudio sobre el estado de los arrecifes parche de Majagual 13 días después de recibir los efectos indirectos del huracán Mitch. Se utilizaron cuadrantes de 1 m2 y técnicas de evaluación rápida. Los resultados se compararon con un trabajo previo realizado en el arrecife de Xahuayxol y revelaron una baja cobertura de coral vivo (11.5% ±5.2) y una elevada cobertura de algas filamentosas y coral antiguamente muerto (53.7% ±27.7 para ambos). Se infiere que el disturbio sobre los corales ya tiene varios años de haber iniciado, mientras que la reducida altura del tapete algal puede deberse, en parte, a los efectos indirectos del huracán Mitch, pues el oleaje resuspendió el sedimento. Asimismo, se observó una mayor frecuencia de encuentros entre Favia fragum (Esper) y algas coralinas incrustantes, que entre otros corales y algas, infiriendo que existe cierta especificidad entre ambos táxones.
Agradecimientos
Se agradece a Juan Pablo Carricart-Ganivet y Julio Espinoza-Avalos (ECOSUR-Chetumal) por su ayuda en el diseño del muestreo, la identificación de las algas y corales y por la revisión del manuscrito. Este trabajo surgió a partir de una práctica de campo conjunta de los cursos Arrecifes Coralinos y Vegetación Costera Tropical de ECOSUR. Juan Pablo Carricart-Ganivet proporcionó un trabajo en preparación que permitió la comparación con el arrecife de Xahuayxol. Sergio Salazar-Vallejo comentó sobre una versión previa. Se agradece a tres árbitros anónimos la revisión que mejoró la versión inicial de este trabajo.
Referencias
Aronson, R.B., P.J. Edmunds, W.F. Precht, D.W. Swanson & D.R. Levitan. 1994. Large-scale, long term monitoring of Caribbean coral reefs: simple, quick, inexpensive techniques. Atoll Res. Bull. 421: 1-19. [ Links ]
Aronson, R.B. & W.F. Precht. 1995. Landscape patterns of reef coral diversity: A test of the intermediate disturbance hypothesis. J. Exp. Mar. Biol. Ecol. 192: 1-14. [ Links ]
Ginsburg, R.N., R.P.M. Bak, W.E. Kiene, E. Gischler & V. Kosmynin. 1996. Rapid assessment of reef condition using coral vitality. Reef Encounter 19: 12-14. [ Links ]
Grigg, R.W. 1992. Coral reef environmental science: truth versus the Cassandra syndrome. Coral Reefs 11: 183-186. [ Links ]
Jackson, J.B.C. 1997. Reefs since Columbus. Proc. 8th Int. Coral Reef Sym. 1: 97-106. [ Links ]
Jordan, D.E. 1979. Estructura y composición de los arrecifes coralinos en la región noreste de la Península de Yucatán: estructura comunitaria, un estimador del desarrollo arrecifal. Tesis doctoral, Universidad Nacional Autónoma de México, Ciudad de México, México. [ Links ]
Lang, J., P. Alcolado, J.P. Carricart-Ganivet, M. Chiappone, A. Curran, P. Dustan, G. Gaudian, F. Geraldes, S. Gittings, R. Smith, W. Tunnell & J. Wiener. 1998. Status of coral reefs in the Northern areas of the Wider Caribbean. p. 123-134, In C. Wilkinson (ed.). Status of coral reefs of the World: 1998. Global Coral Reefs Monitoring Network, Queensland. [ Links ]
Lapointe, B.E. 1989. Caribbean coral reefs: are they becoming algal reef? Sea Frontiers p. 84-91. [ Links ]
Littler, M.M., D.S. Littler & P.R. Taylor. 1983. Evolutionary strategies in a tropical barrier reef system: Functional-form groups of marine macroalgae. J. Phycol. 19: 229-237. [ Links ]
Risk, M.J. & P.W. Sammarco. 1982. Bioerosion of corals and the influence of damselfish territoriality: a preliminary study. Oecologia 52: 376-380. [ Links ]
Salazar-Vallejo, S.I. & N.E. González. 1994. Turismo costero y conservación: Competencia o colaboración? Revista Caribe, CIQRO 3: 1-28. [ Links ]
Steneck, R.S. & M.N. Dethier. 1994. A functional group approach to the structure of algal-dominated communities. Oikos 69: 476-498. [ Links ]
Steneck, R.S., J.C. Lang, P.A. Kramer & R.N. Ginsburg. 1997. Atlantic and Gulf Reef Assessment (AGRA) Rapid Assessment Protocol (RAP). Página Web consultada (16/noviembre/1998) en: http://coral.aoml.noaa.gov/agra/agra1.html. [ Links ]
Tanner, J.E. 1995. Competition between scleractinian corals and macroalgae: An experimental investigation of coral growth, survival and reproduction. J. Exp. Mar. Biol. Ecol. 190: 151-168. [ Links ]
Tunnell, J.W.A., A. Rodríguez, R. Lehman & C. Beaver. 1993. An ecological characterization of the southern Quintana Roo coral reef system. Texas A&M University, Texas 36 p. [ Links ]
1 El Colegio de la Frontera Sur, Unidad Chetumal. Dep. de Ecología Acuática. Apdo. Postal 424, Chetumal, C.P. 77000, QROO, México. Fax: (+983) 204-47, rolando@ecosur-qroo.mx

