











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
La fragmentación se refiere a la pérdida de continuidad de la vegetación natural o ecosistemas, dejando remanentes de menor tamaño (parches o fragmentos) lo cual afecta tanto la estructura como la diversidad del paisaje. Los procesos más generales que propician la fragmentación de un paisaje son la actividad ganadera y la agrícola, la urbanización y la creación de infraestructura (Salazar et al., 2017).
Con la finalidad de atenuar los efectos ecológicos negativos de la fragmentación es necesario mantener la conectividad del paisaje conservando las relaciones espaciales entre los elementos que lo conforman. La conectividad estructural se presenta cuando dos o más fragmentos se mantienen conectados por una franja de clase de hábitat similar y su ausencia incrementa la fragmentación del paisaje (Jiménez-Ramírez y López-Arévalo, 2021).
Existen varias razones por las cuales no hay una única fórmula de cálculo universal (Aguilar et al, 2014), entre éstas están la complejidad y diversidad de los sistemas ecológicos , la diversidad de organismos y paisajes dado que cada especie tiene diferentes requerimientos de hábitat y patrones de movimiento; las diferencias en escalas espaciales y temporales ya que las mismas pueden variar significativamente, desde individuos hasta poblaciones y ecosistemas enteros; o debido a las múltiples dimensiones de la conectividad dado que la misma involucra no solo la distancia física entre parches de hábitat, sino también factores como la calidad del hábitat, la presencia de barreras y la interacción entre especies.
Combinar un índice de conectividad estructural como la fragmentación de hábitats con un índice de ocupación de las especies es una estrategia valiosa en ecología, especialmente cuando se busca entender las dinámicas de las poblaciones y su relación con el paisaje considerando que la conectividad funcional mide la capacidad de los organismos de moverse y persistir en un paisaje, considerando factores como la distancia entre parches de hábitat, la calidad del hábitat y las barreras y un Índice de ocupación indica la probabilidad de que una especie esté presente en un sitio específico (Gurrutxaga, 2004).
Esta investigación se desarrolló con el objetivo de brindar una alternativa de medición para la conectividad funcional que los corredores biológicos (CB) están aportando a la conservación de mamíferos silvestres con algún grado de amenaza o vulnerabilidad en Costa Rica.
Materiales y métodos
Consideré como unidad territorial de análisis 51 corredores biológicos de Costa Rica, dentro de los cuales delimité su cobertura forestal según tipos de bosque (SINAC, 2021), información con la que calculé la fragmentación de ecosistemas según Morera y Sandoval (2018), mediante el software Patch Analyst de ArcView 3.3™ de ESRI© (Environment Systems Research Institute Inc.) según Elkie et al. (1999):
IFcb = TLA / CA / (Nump * MNN)
donde,
IFcb= es el índice de fragmentación en un área de estudio (en este estudio corredores biológicos) TLA= superficie total del área de estudio en Ha.
CA= superficie total de tipo de bosque (o ecosistema) NUMP= # manchas
MNN= distancia media en metros desde el borde de una mancha hasta el borde de la mancha más cercana
Determiné las especies por evaluar, lo cual para este estudio lo establecí con el apoyo del departamento de participación ciudadana-gobernanza y el programa nacional de corredores biológicos del SINAC (Jairo Sancho, com. pers.), y definí como fuente de información los registros de las especies de la plataforma oficial del Ministerio de Ambiente y Energía iNaturalist (GBIF) y ciencia ciudadana, bases de datos sin limitación de accesibilidad y descargables para las especies preestablecidas (Hernández et al., 2022; Monge-Velázquez, 2024).
Con la ubicación de avistamientos de las especies priorizadas que provee las bases de datos de MINAE-iNaturalist, georreferencié y calculé la función de distribución de probabilidad Kernel (Silverman, 1998) según criterio de Epanechnikov para determinar los “mapas de calor” de observaciones ciudadanas, utilizando ArcGIS 10.5™ de ESRI© (Environment Systems Research Institute Inc.)
Con dicho cálculo de probabilidad de distribución, reclasifiqué el ráster en presencia / ausencia de la especie en estudio para vectorizar en anillos de presencia dichas áreas para cada especie. Con dichas áreas de presencia probable de la especie, extraje las áreas coincidentes con los distintos corredores biológicos de Costa Rica.
Contando con estas áreas de ocupación de las especies, y con las áreas totales de los corredores biológicos calculé un Índice de Ocupación de especies, diseñando la siguiente fórmula para este estudio:
IOsp = Aosp / At
donde,
IOsp= es el índice de ocupación de la especie en un área de estudio (en este estudio corredores biológicos)
Aosp= es el área ocupada teórica o real por la especie en un área de estudio
At= es el área total de estudio, sea un área protegida, corredor biológico, cantón, país, etc.
A partir del cálculo de la fragmentación de los tipos de bosques en los corredores biológicos, y la ocupación de las especies en los mismos, procedí a definir la conectividad funcional real de las especies en estudio.
Ésta es la relación entre la fragmentación y su ocupación real mediante sus índices estandarizados, cuyo valor dividido entre 100 tiene un rango de 0 a 1, siendo 0 una conectividad funcional relativa (fortuita o esporádica), y 1 una conectividad funcional totalmente activa para la especie en el corredor biológico. Como alternativa de medición de esta conectividad funcional, considerando los anteriores criterios, diseñé la siguiente fórmula de cálculo:
ICFRsp = IFcb / IOsp / 100
donde,
ICFRsp= es el índice de conectividad funcional real de la especie
IFcb= es el índice de fragmentación en un área de estudio (en este estudio corredores biológicos) IOsp= es el índice de ocupación de la especie en un área de estudio (en este estudio corredores biológicos)
Realicé análisis estadísticos de correlación de las variables en estudio mediante el coeficiente de correlación de rangos de Spearman, y un análisis PCoA con la matriz triangular de disimilitudes Bray-Curtis presencia-ausencia de las especies y su relación con las variables ecológicas (fragmentación, ocupación y conectividad) como método de ordenación, utilicé para esto el software Past 4.04 (Hammer et al., 2001).
Resultados
Los mayores índices de fragmentación se sugieren para cuatro corredores biológicos interurbanos (María Aguilar, Cobri-Surac, Nambiral y Cubujuquí), por su parte los corredores biológicos con mínimos efectos en su fragmentación de la cobertura forestal están alejados de zonas urbanas, como Río Nosara, Acuíferos, Los Santos y Central-Talamanca (Tabla 1).
Tabla 1 Índice de fragmentación en los corredores biológicos del SINAC
| N | Corredor Biológico | CA_Ha | TLA_Ha | NumP | MNN(m) | IF |
| 1 | Interurbano María Aguilar | 192,80 | 3 873,63 | 95,00 | 182,11 | 38,51 |
| 2 | Cobri-Surac | 2 770,23 | 16 766,24 | 609,00 | 350,58 | 3,48 |
| 3 | Nambiral | 2 716,24 | 10 830,95 | 1 983,00 | 1 455,12 | 2,93 |
| 4 | Interurbano Cubujuquí | 4 323,96 | 26 159,94 | 1 428,00 | 318,74 | 1,35 |
| 5 | Pará | 941,43 | 2 198,18 | 292,00 | 76,51 | 0,61 |
| 6 | Achiote | 1 583,20 | 5 491,27 | 693,00 | 120,66 | 0,60 |
| 7 | OSREO | 10 286,84 | 26 032,70 | 3 291,00 | 771,94 | 0,59 |
| 8 | Garcimuñoz | 9 541,02 | 29 798,99 | 1 958,00 | 325,01 | 0,52 |
| 9 | Colorado-Tortuguero | 15 357,99 | 36 279,65 | 5 617,00 | 1 207,73 | 0,51 |
| 10 | Parismina | 8 162,41 | 31 489,64 | 6 092,00 | 447,22 | 0,28 |
| 11 | Miravalles-Rincón de la Vieja | 1 145,19 | 4 348,82 | 1 852,00 | 105,98 | 0,22 |
| 12 | Montes del Aguacate | 17 994,44 | 55 216,67 | 4 186,00 | 280,63 | 0,21 |
| 13 | Morocochas | 26 036,05 | 51 672,52 | 15 814,00 | 1 621,44 | 0,20 |
| 14 | Osa | 31 024,26 | 49 322,24 | 9 585,00 | 1 096,30 | 0,18 |
| 15 | Bolsón-Ortega | 1 677,47 | 9 217,45 | 6 324,00 | 173,60 | 0,15 |
| 16 | Fuente de Vida-Amistad | 4 168,67 | 10 435,73 | 1 486,00 | 88,02 | 0,15 |
| 17 | Mono Aullador | 37 867,29 | 98 520,37 | 24 229,00 | 1 359,78 | 0,15 |
| 18 | Playa Hermosa | 7 883,10 | 13 104,15 | 1 920,00 | 159,34 | 0,14 |
| 19 | Las Camelias | 6 499,93 | 22 893,87 | 4 176,00 | 161,29 | 0,14 |
| 20 | Alexander Skutch | 2 853,60 | 6 010,76 | 1 796,00 | 93,63 | 0,11 |
| 21 | Río Naranjo | 8 614,65 | 18 990,32 | 3 178,00 | 157,85 | 0,11 |
| 22 | Moín-Tortuguero | 9 955,10 | 13 622,42 | 5 673,00 | 380,22 | 0,09 |
| 23 | Diriá | 64 272,03 | 102 082,13 | 59 169,00 | 3 245,25 | 0,09 |
| 24 | Peninsular | 44 826,46 | 64 028,41 | 27 765,00 | 1 652,04 | 0,08 |
| 25 | Pájaro Campana | 35 758,58 | 78 318,08 | 19 719,00 | 730,29 | 0,08 |
| 26 | Río Cañas | 3 199,34 | 5 251,70 | 2 061,00 | 77,67 | 0,06 |
| 27 | Rincón-Cacao | 2 579,39 | 2 798,75 | 1 184,00 | 65,34 | 0,06 |
| 28 | Paso de las Lapas | 21 228,73 | 38 949,96 | 14 215,00 | 444,70 | 0,06 |
| 29 | Cordillera | 10 463,81 | 13 897,81 | 7 976,00 | 342,19 | 0,06 |
| 30 | Jesús | 10 646,93 | 19 359,27 | 9 361,00 | 279,42 | 0,05 |
| 31 | Baulas | 7 947,59 | 14 081,64 | 6 635,00 | 194,72 | 0,05 |
| 32 | Barra Honda-Rosario | 8 316,88 | 18 192,56 | 13 078,00 | 283,41 | 0,05 |
| 33 | Tres Colinas | 1 467,62 | 1 912,19 | 1 364,00 | 43,64 | 0,04 |
| 34 | Ruta de los Maleku | 53 296,15 | 136 632,78 | 28 318,00 | 449,91 | 0,04 |
| 35 | Arenal-Tenorio | 13 139,55 | 34 093,17 | 12 311,00 | 179,14 | 0,04 |
| 36 | San Juan-La Selva | 83 018,89 | 174 025,58 | 48 557,00 | 754,46 | 0,03 |
| 37 | Amistosa | 48 245,86 | 91 737,24 | 17 027,00 | 288,51 | 0,03 |
| 38 | Tenorio-Miravalles | 6 498,60 | 12 695,82 | 4 956,00 | 79,16 | 0,03 |
| 39 | Bosque Agua | 10 813,17 | 15 850,33 | 4 972,00 | 104,22 | 0,03 |
| 40 | Paso de las Nubes | 17 602,14 | 40 078,42 | 13 582,00 | 153,15 | 0,03 |
| 41 | Langusiana | 19 240,26 | 32 916,14 | 9 758,00 | 146,16 | 0,03 |
| 42 | Hojancha | 31 080,80 | 41 798,27 | 15 215,00 | 260,64 | 0,02 |
| 43 | Chirripó-Savegre | 17 637,36 | 34 515,74 | 14 415,00 | 154,98 | 0,02 |
| 44 | Paso de la Danta | 59 130,02 | 79 353,97 | 29 072,00 | 437,14 | 0,02 |
| 45 | Rain Forest | 13 718,76 | 26 116,65 | 12 738,00 | 109,88 | 0,02 |
| 46 | Talamanca-Caribe | 23 772,68 | 27 056,55 | 19 013,00 | 266,06 | 0,02 |
| 47 | Potrero-Caimital | 13 868,03 | 19 952,09 | 9 658,00 | 103,31 | 0,02 |
| 48 | Río Nosara | 21 312,65 | 28 561,36 | 12 633,00 | 132,90 | 0,01 |
| 49 | Acuíferos | 15 471,59 | 25 897,62 | 14 864,00 | 83,33 | 0,01 |
| 50 | Los Santos | 93 266,40 | 156 687,64 | 53 051,00 | 284,79 | 0,01 |
| 51 | Central-Talamanca | 47 936,26 | 90 730,70 | 47 117,00 | 144,32 | 0,01 |
* CA-Ha= Superficie total de tipo de bosque (o ecosistema); TLA-Ha= superficie total del área de estudio en Ha.; NumP= # manchas; MNN (m)= distancia media en metros desde el borde de una mancha hasta el borde de la mancha más cercana; IF= índice de fragmentación.
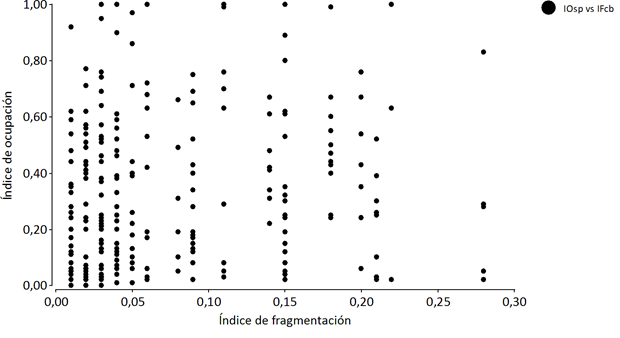
La correlación entre la fragmentación y la ocupación de las 13 especies en los 51 corredores biológicos sugiere que la misma es débil, con significancia estadística (Fig. 1).
Únicamente la especie de felino manigordo u ocelote (Leopardus pardalis) sugiere una conectividad funcional casi total en el corredor biológico Acuíferos (ICFR = 0,98; Fig. 3G), siendo la especie con mayor presencia en los corredores biológicos, en 38 de los 51 corredores evaluados.
Seguida por el mono titi o ardilla (Saimiri oerstedii) en el corredor biológico Los Santos con una conectividad alta (ICFR = 0,69; Fig. 3E), no obstante, a estar presente únicamente en 13 de los corredores biológicos estudiados (Apéndice 1).
r (gl 301) = 0,11; p = 0,05*
El mono congo (A. palliata) fue la segunda especie con más presencia en los corredores biológicos, en 37 de los 51 corredores evaluados, a su vez que registró su mayor conectividad en el CB. Talamanca-Caribe con (ICFR = 0,48; Fig. 3A; Apéndice 1).
La especie con menor presencia en corredores biológicos fue el chancho de monte o pecari (T. pecari) en siete de los 51 corredores evaluados, siendo el CB. Rincón-Cacao el que presentó mayor conectividad (ICFR = 0,17; Fig. 3K; Apéndice 1).
Los corredores biológicos que registraron mayor cantidad de especies fueron CB. Osa y CB. Arenal-Tenorio con 12 especies, mientras que los CB. Las Camelias y CB. Potrero-Caimital únicamente registraron una especie (Apéndice 1).
El análisis de coordenadas principales reveló patrones de asociación, co-ocurrencia o similitud entre algunas especies como S. oertedii, L. tigrinus, L. pardalis, L. wiedii, o T. bairdii, A. palliata y A. geoffroyi. No obstante, la matriz triangular de distancias y su relación con las variables ecológicas sugieren débil diferenciación respecto al espacio de coordenadas y su influencia en la presencia-ausencia de las especies, con excepción de T. bairdii, A. palliata y A. geoffroyi (Fig. 2).
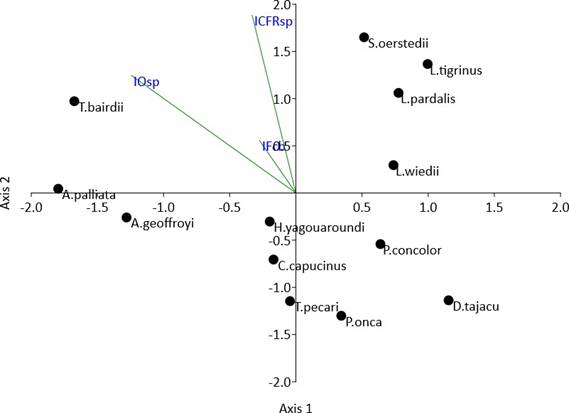
Fig. 2 PCoA matriz triangular Bray-Curtis disimilitud presencia-ausencia especies y relación de fragmentación, ocupación e índice de conectividad funcional real.
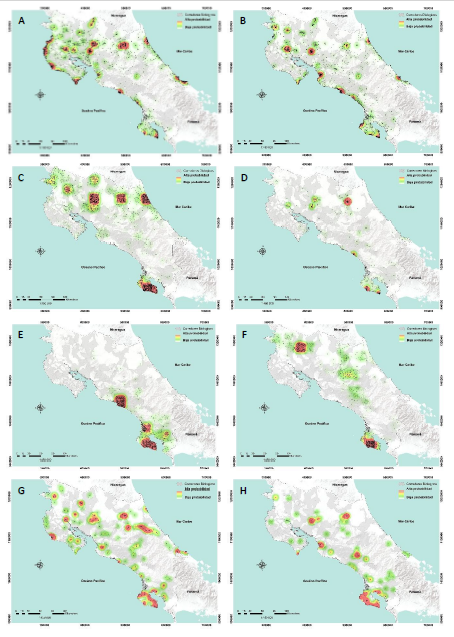
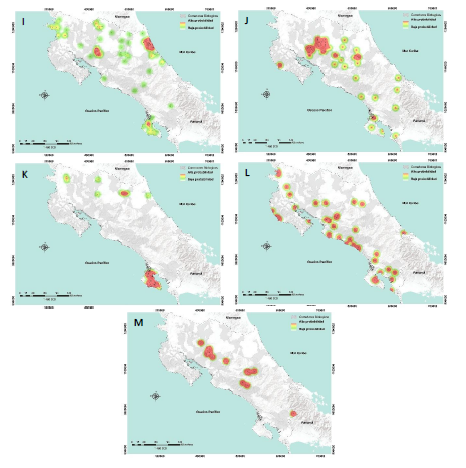
*Nota: Las versiones en alta resolución de estas imágenes están disponibles para su descarga y visualización detallada en el siguiente enlace: Mapas HQ
Fig. 3. Distribución de probabilidad mediante Epanechnikov-Kernel por especie: A) Alouatta palliata; B) Cebus capucinus; C) Ateles geoffroyi; D) Dasyprocta tajacu; E) Saimiri oerstedii; F) Tapirus bairdii; G) Leopardus pardalis; H) Puma concolor; I) Panthera onca; J) Leopardus wiedii; K) Tayassu pecari; L) Herpailurus yagouaroundi; M) Leopardus tigrinus.
Discusión
La conectividad de y entre los ecosistemas tiene muchas alternativas de medición, en este estudio se utilizó la fragmentación de los tipos de bosque en Costa Rica y la ocupación de las especies priorizadas y su relación como cálculo matemático para mostrar esa conectividad. Este criterio de ocupación es sugerido por Leija y Mendoza (2021) para establecer la conectividad funcional real para especies en estudio en determinadas áreas de análisis.
A partir de la definición de todos los polígonos de tipos de bosques, se sugiere que la mayoría de los corredores biológicos presentó bajos índices de fragmentación, lo cual es congruente con el aumento gradual de las últimas décadas del crecimiento de bosques, particularmente secundarios (MINAE et al., 2018)
De las 13 especies de mamíferos evaluadas, cuatro (mono Titi, el tigrillo, mono araña y el mono congo) sugieren que utilizan los corredores biológicos como parte obligada de su ámbito de hogar, con excepción del felino ocelote o manigordo, el cual en el corredor biológico Acuíferos indica una conectividad funcional casi total (ICFR = 0,98), siendo 0 una conectividad nula, y 1 conectividad total del corredor para la especie.
Este registro para el ocelote es similar al reportado por Hernández-Pérez et al. (2024) en condiciones mayormente fragmentadas en la Sierra Norte de Puebla en el caribe mejicano (entre 0,97 y 0,99 utilizando un índice de éxito de conectividad por cambios en la cobertura forestal y presencia de la especie), conclusión a la que llegan estos autores con apenas 13 registros de la especie, registros limitados en comparación con las 253 observaciones utilizadas en la presente investigación para Costa Rica. Considerando que el CB. Acuíferos en Costa Rica presentó bajo índice de fragmentación (0,01) es coincidente con Hernández-Pérez et al. (2024) y Galindo-Aguilar et al. (2019) en que esta especie de felino prefiere regiones con poca deforestación, siendo por consiguiente los bosques los hábitats que utiliza como corredor para su desplazamiento.
Otra especie de felino silvestre que permite comparación con otras regiones de Latinoamérica al contar con estudios sobre su conectividad estructural es la oncilla o tigrillo (L. tigrinus), especie que en Colombia, en parte y cercanías de la Reserva Forestal Protectora Productora Cuenca Alta del Río Bogotá, registró índices de conectividad en ambientes con distintos grados de fragmentación entre 0,19 y 0,36, siendo más frecuente en ambientes menos fragmentados (Jiménez-Ramírez y López-Arévalo, 2021), mientras que en Costa Rica, de manera muy similar en comportamiento, en el presente estudio la especie sugiere alta conectividad en corredores biológicos con baja fragmentación (0,01) como Central-Talamanca (ICFR = 0,63); CB. Los Santos (ICFR = 0,27) y Arenal-Tenorio (ICFR = 0,16).
Sugiero que la métrica propuesta es adecuada y sensible para calcular la conectividad para especies arbóreas como el mono congo (A. palliata) en comparación con otras métricas de conectividad más complejas como el modelo de Montilla-Rivas et al. (2025), estudio en el cual la conectividad probable para esta especie es de 0,0058, parámetro establecido en áreas de gran cobertura forestal. Otras especies que mostraron sensibilidad a la métrica fueron T. bairdii, A. geoffroyi y S. oerstedii. En el presente trabajo y con la métrica de relación fragmentación y ocupación de las especies, el mono congo registró conectividad funcional de 0,48 en un corredor con baja fragmentación como Talamanca-Caribe; 0,30 en el CB. Tenorio-Miravalles; y 0,24 en el CB. Arenal-Tenorio.
Los resultados sugieren el uso de los corredores biológicos por parte de las especies como parte de un ámbito hogareño mucho mayor, donde los corredores biológicos sólo forman parte de un ensamblaje ecosistémico heterogéneo, no obstante, los índices de fragmentación muestran que son los corredores biológicos interurbanos (e.g. Cobri-Surac, Nambiral, Apéndice 1) los más fragmentados, pero usados homogéneamente por las especies. Se evidencia como la disimilitud de presencia-ausencia de registros para las especies ocurren en espacios de coordenadas no necesariamente relacionados con su ocupación en los corredores biológicos, y poco influenciados por la fragmentación. (Fig. 2)
El análisis podría complementarse con el uso de esta metodología propuesta aplicada a los territorios de áreas silvestres protegidas, y comparar con los corredores biológicos, sugiriéndose la verdadera complementariedad entre ambos sistemas de gestión de la cobertura forestal del país.
Ética, conflicto de intereses y declaración de financiamiento
Declaro haber cumplido con todos los requisitos éticos y legales pertinentes, tanto durante el estudio como en la preparación de este documento; que no hay conflictos de interés de ningún tipo, y que todas las fuentes financieras se detallan plena y claramente en la sección de agradecimientos. Asimismo, estoy de acuerdo con la versión editada final de esta publicación. El respectivo documento legal firmado se encuentra en los archivos de la revista.
La declaración de contribución de cada autor es la siguiente: J.C.V.A: Diseño del estudio, recolección y análisis de datos, preparación y aprobación final del manuscrito.

