











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
Los cambios en el uso del suelo de bosques nativos a tierras usadas en agricultura, ganadería y plantaciones forestales, afectan las comunidades acuáticas, ya que la pérdida de vegetación ribereña altera directa e indirectamente la calidad del agua, el tipo de sustrato y afecta las condiciones físicas del hábitat acuático y las comunidades bióticas asociadas (e.g. Giraldo, Chará, Zúñiga, Chará-Serna & Pedraza, 2014; Galeano-Rendón, Monsalve-Cortes, & Mancera-Rodríguez, 2017), disminuyendo el material alóctono que ingresa al cuerpo hídrico y la disponibilidad de sustratos aportados por los bosques adyacentes (e.g. Bücker, Sondermann, Frede, & Breuer, 2010; Giraldo et al., 2014), y alterando la estructura y la función trófica de las comunidades de insectos acuáticos (e.g., Lorion & Kennedy, 2009; Ferrú & Fierro, 2015; Iñiguez-Armijos, Hampel, & Breuer, 2018).
Para evaluar los impactos antrópicos en los ecosistemas acuáticos, muchos estudios utilizan indicadores biológicos que proporcionan información sobre la condición ecológica y la evaluación de ecosistemas fluviales, en consecuencia los ensamblajes de macroinvertebrados acuáticos son excelentes indicadores de la calidad del agua debido a la variedad de sensibilidad a la contaminación orgánica y tolerancia a diferentes alteraciones de su hábitat (Figueroa, Palma, Ruiz, & Niell, 2007; Barba, Lanza, Ramos, & González, 2013).
Diversos estudios han encontrado que los cambios en el uso de la tierra y por consiguiente en las coberturas vegetales de los márgenes de las quebradas, tienen efectos sobre los ensamblajes de macroinvertebrados acuáticos, con variaciones en la composición taxonómica y de grupos funcionales y una disminución de la diversidad en quebradas con coberturas de bosques nativos, con relación a quebradas con coberturas de plantaciones de bosques exóticos, y de pastos para actividades agrícolas y ganaderas (e.g. Lorion & Kennedy, 2009; Fierro et al., 2017; Iñiguez-Armijos et al., 2018).
En Ecuador se han desarrollado estudios con macroinvertebrados acuáticos como indicadores biológicos que proporcionan información sobre la condición ecológica de diversos cuerpos hídricos (e.g. Giacometti & Bersosa, 2006; Arroyo & Encalada, 2009; Terneus, Hernández, & Racines, 2012; Álvarez-Mieles et al., 2013; Villamarín, Rieradevall, Paul, Barbour, & Prat, 2013; Damanik-Ambarita et al., 2016; Liñero et al., 2016), sobre la diversidad taxonómica y funcional en relación con los regímenes de caudal y disturbios hidrológicos o como respuesta a factores ambientales y condiciones fisicoquímicas (e.g., Jacobsen & Encalada, 1998; Carrera & Gunkel, 2003; Ríos-Touma, Encalada, & Prat, 2009, 2011), existiendo poca información acerca de los efectos de los cambios de la vegetación riparia sobre las comunidades de macroinvertebrados acuáticos (e.g.; Bojsen & Jacobsen, 2003; Bücker et al., 2010; Iñiguez-Armijos, Leiva, Frede, Hampel & Breuer, 2014; Iñiguez-Armijos et al., 2016; Iñiguez-Armijos et al., 2018).
Dado el incremento de pérdida de bosques por cambios en el uso de suelo con el consecuente aumento de afectaciones por actividades antrópicas y la falta de conocimiento de la respuesta del ensamblaje de macroinvertebrados a dichas alteraciones, el presente estudio espera proporcionar información sobre los efectos de las variaciones de la vegetación de ribera asociada a tres quebradas tropicales con diferentes coberturas de vegetación riparia (bosque nativo, pastos/cultivos y plantación forestal) sobre las comunidades de macroinvertebrados en dos períodos hidrológicos (seca vs. lluviosa) contrastantes en el bosque protector Murocomba, Provincia de Los Ríos, Ecuador. Se plantea como hipótesis de investigación que la quebrada con márgenes riparios con uso de bosque nativo presentará mejor condición ecológica, por lo tanto, la diversidad taxonómica (estructura y composición) a nivel de familias será mayor que en zonas con coberturas en pastos para uso agrícola y ganadero y de plantación forestal.
La información de este estudio servirá como base ecológica para futuros estudios de calidad de agua mediante el empleo de macroinvertebrados acuáticos como índices biológicos, contribuirá a la generación de información sobre las respuestas del ensamblaje de macroinvertebrados acuáticos al cambio en la cobertura ribereña de quebradas, útil para la formulación de estrategias efectivas de conservación, restauración y manejo de bosques de ribera en el área de bosque protector Murocomba y a la mitigación de los efectos negativos de las actividades agropecuarias sobre las zonas de margen de quebradas y cuerpos hídricos.
Materiales y Métodos
Área de estudio: El trabajo se realizó en tres quebradas de la zona del Bosque Protector Murocomba, en la provincia Los Ríos, Ecuador, con diferentes tipos de cobertura vegetal en sus márgenes. El rango altitudinal de los sitios de estudio se encuentra entre los 415 y 613 m, y de acuerdo a Holdridge (1982) corresponde a una zona de bosque húmedo premontano (bh-PM), con una temperatura que oscila entre 18 y 23 °C.
La quebrada La Victoria (Q1-CBN) (0º38’18.1’’ S & 79º08’53.7’’ W, 613 m.s.n.m.) presenta cobertura de bosque nativo secundario en sus dos márgenes en buen estado de conservación; tiene una amplia franja de vegetación ribereña y un dosel que alcanza más de 15 m de altura, con presencia de vegetación arbórea nativa y especies como Guachapelí (Albizia guachapeli Kunth), Naranjillo (Trichanthera gigantea Humb. & Bonpl.), Tillo (Brosimum acutifolium Huber), Clavellín (Brownea herthae Harms), Caucho (Ficus elastica Hornem), Guarumo (Cecropia peltata L.), Cedrillo (Terminalia sp.), Laurel blanco (Cordia alliodona Johnst.), Guabo (Inga edulis Mart.), Pambil (Iriartea sp.), Tomate de árbol de montaña (Solanum betaceum Cav.), Moral Fino (Chlorophora tinctoria L. Gaudich), Balsa (Ochroma pyramidale Cav. ex Lam. Urb.), entre otras. La quebrada El Congo (Q2-CAG) (0º37’32.9’’ S - 79º09’ 07.7’’W, 533 m.s.n.m.) corresponde a la zona de cobertura en pastos con actividades agrícolas y ganaderas; sus márgenes están cubiertos principalmente por pastos y algunos cultivos de maíz, naranjilla, plátano y cacao. La quebrada La Damita (Q3-CPF) (0º38’38.2’’ S & 79º10’46.7’’ W, 415 m.s.n.m.) corresponde a la zona de cobertura de plantaciones forestales de especies no nativas de la zona como Melina (Gmelina arborea Roxb.) y Pachaco (Schizolobium parahybum Blake) de más de 5 ha de extensión con árboles de cerca de 7 m de altura.
Variables fisicoquímicas e hidrológicas: Se midieron las siguientes variables in situ: oxígeno disuelto (mg/L), pH, conductividad (μS/cm), temperatura superficial del agua (°C), y sólidos totales disueltos STD (mg/L) con un medidor Hanna HI-9829. El ancho y la profundidad fueron medidos con flexómetro, y una vara graduada en cm. El caudal (Q) se determinó a través del método del objeto flotante, siendo Q = (Fc) x (A) x (V), donde Fc representa el factor de corrección (0.70) para canal en tierra con profundidad > 15 cm, A representa el área y V la velocidad. La velocidad de la corriente se midió con la ayuda de un objeto flotador en cuatro puntos del cauce, midiendo el tiempo que demoró en recorrer cinco metros de distancia. El área de la sección transversal se calculó midiendo el ancho del río en cuatro puntos distintos de las márgenes y sobre esta referencia se midió la profundidad en puntos consecutivos ubicados cada 20 cm.
Recolecta e identificación de macroinvertebrados acuáticos: La recolección de macroinvertebrados acuáticos se realizó en dos períodos hidrológicamente distintos: (i) temporada seca entre junio y noviembre de 2014 y (ii) temporada lluviosa entre diciembre de 2014 y abril de 2015. En cada período hidrológico se realizaron cuatro muestreos con un intervalo de 45 días entre cada muestreo. Los macroinvertebrados acuáticos se recolectaron usando una red D-net de 405 cm2 y con malla de 500 µm ubicada en sentido contrario de la corriente, para lo cual se seleccionaron 20 puntos al azar a lo largo del tramo de 100 m en cada quebrada, removiendo hojarascas y sedimento in situ en el cauce de las quebradas. El material recolectado fue etiquetado y almacenado en recipientes con alcohol al 70 %. La identificación taxonómica de los especímenes se realizó hasta el nivel de familia utilizando un estereoscopio y claves taxonómicas especializadas (Roldán, 2003; Domínguez & Fernández, 2009) y las muestras se encuentran depositadas en el laboratorio de Microbiología y Entomología de la Universidad Técnica Estatal de Quevedo.
Índices biológicos: Para determinar la condición ecológica se calcularon los valores del Índice Biótico de Familias (IBF), el Índice de porcentaje de individuos de Ephemeroptera, Plecoptera y Trichoptera (EPT) y el Índice Biological Monitoring Working Party (BMWP/Col), para cada estación de muestreo y período climático.
Análisis de datos: Para establecer si existieron diferencias significativas en los valores de las variables fisicoquímicas y los valores de caudal, así como de los índices IBF, EPT y BMWP/Col entre períodos hidrológicos y entre quebradas con diferente tipo de cobertura, se aplicaron análisis de varianza unidireccionales (ANOVA) a cada variable, previa validación de los supuestos de normalidad (Kolmogorov-Smirnov) y homocedasticidad (prueba de Levene) y se realizaron comparaciones evaluadas mediante la prueba de Tukey. Cuando los datos no cumplieron con los supuestos de normalidad y homocedasticidad se utilizó la prueba no paramétrica de Kruskall-Wallis.
Se determinó el número de familias e individuos, y se calcularon los índices de Shannon-Weaver, Dominancia, Equidad de Simpson, número de taxones raros (abundancia inferior al 3 %), para cada estación de muestreo y período hidrológico, utilizando el programa estadístico PAST (Hammer, Harper, & Ryan, 2001). Los valores de los índices de diversidad Shannon-Weaver y dominancia de Simpson entre estaciones y períodos se compararon mediante pruebas de t de Hutcheson (Zar, 1999).
Se definieron los patrones de variación en la composición de familias del ensamblaje de macroinvertebrados entre quebradas con diferentes coberturas con respecto a las variables ambientales y a los valores de los Índices IBF, EPT y BMWP-Col, mediante análisis discriminante canónico (ADC), que se realizó sobre datos de abundancia de los taxones que presentaron una frecuencia mayor al 5 % y una abundancia superior a 3 % (Gauch, 1982; Ríos-Pulgarín, Barletta, Arango-Jaramillo, & Mancera-Rodríguez, 2016a). En este análisis, los taxones que ayudan a explicar las diferencias se representan como vectores cuya longitud y dirección es proporcional a su contribución para explicar la variabilidad (Ríos-Pulgarín, Barletta, & Mancera-Rodríguez, 2016b). Todos los análisis se desarrollaron en versiones R Software 3.1.1 (R Core Team, 2012) y RWizard (Guisande, 2015).
Resultados
Variables fisicoquímicas e hidrológicas: La quebrada La Victoria (Q1-CBN) presentó un sustrato pedregoso y rocoso, con pendiente inclinada y con caudal medio de 0.292 m3/s en período seco y de 0.590 m3/s en período de lluvias. Presentó un ancho medio de 4.25 m en el período seco y de 4.50 m en el período de lluvias y una profundidad entre 25 y 30 cm. La quebrada El Congo (Q2- CAG), presentó un sustrato pedregoso, con pendiente plana y con caudal medio de 0.108 m3/s en período seco y de 0.815 m3/s en período de lluvias. Presentó un ancho medio de 3.85 m en el período seco y de 5.70 m en el período de lluvias y una profundidad entre 14 y 24 cm. Por último, la quebrada La Damita (Q3-CPF) posee un sustrato arenoso con pendiente plana y con caudal medio de 0.365 m3/s en período seco y de 0.829 m3/s en período de lluvias. Presentó un ancho medio de 5.70 m en el período seco y de 6.30 m en el período de lluvias y una profundidad media entre 15 y 22 cm (Tabla 1).
Se encontraron diferencias significativas en los valores de oxígeno disuelto entre períodos hidrológicos para las tres quebradas estudiadas (Kruskal-Wallis, P < 0.05) con mayores valores medios durante el período de lluvia entre 9.4 y 9.7 mg/l y una marcada disminución en el período seco con valores medios entre 4.8 y 5.4 mg/l (Tabla 1). Se encontró también diferencia significativa en la quebrada El Congo (Q2-CAG) que tiene márgenes con uso de suelo agrícola-ganadero para los valores de caudal (Kruskal-Wallis, P = 0.043) que fueron mayores en el período de lluvia. Las demás variables fisicoquímicas no presentaron diferencias significativas entre períodos hidrológicos para cada quebrada, aunque la temperatura superficial del agua fue mayor para las tres quebradas en la temporada seca (Tabla 1).
TABLA 1: TABLE 1: Valores de los índices y características químicas durante las temporadas seca y lluviosa en tres quebradas con diferentes coberturas riparias en el Bosque Protector Murocomba, Ecuador Values of the indexes and chemical characteristics during dry and rainy seasons in three creeks with different riparian cover in the Murocomba Protector Forest, Ecuador
| - | Q1-CBN | - | Q2-CAG | - | Q3-CPF | - |
|---|---|---|---|---|---|---|
| - | Seca | Lluvia | Seca | Lluvia | Seca | Lluvia |
| Número de familias | 36 | 26 | 36 | 25 | 39 | 25 |
| Número de individuos | 885 | 1192 | 768 | 797 | 1261 | 1680 |
| Índice de Shannon (H´) | 2.56 | 2.30 | 2.57 | 2.59 | 2.52 | 1.59 |
| Dominancia (D) | 0.12 | 0.14 | 0.12 | 0.11 | 0.12 | 0.36 |
| Equidad de Simpson (1-D) | 0.88 | 0.86 | 0.88 | 0.89 | 0.88 | 0.64 |
| Número de taxones raros | 29 | 19 | 27 | 16 | 31 | 19 |
| Índice IBF | 4.3 ± 0.2 | 3.9 ± 0.7 | 4.9 ± 0.7 | 5.3 ± 0.9 | 5.0 ± 0.0 | 5.1 ± 0.2 |
| Taxones EPT | 7.3 ± 3.0 | 7.5 ± 0.6 | 7.0 ± 1.4 | 6.8 ± 1.3 | 7.5 ±2.9 | 7.0 ± 0.8 |
| Índice % EPT | 51.4 ± 19.6 | 61.7 ± 9.3 | 60.5 ± 2.7 | 51.4 ± 10.2 | 47.8 ± 19.6 | 71.9 ± 9.4 |
| BMWP/Col | 111.3 ± 4.3 | 120.5 ± 36.8 | 105.5 ± 6.6 | 106.5 ± 25.2 | 98.0 ± 5.5 | 115.8 ± 17.7 |
| O2 (mg/L) | 4.9 ± 0.6 | 9.7 ± 1.3 | 5.4 ± 0.1 | 9.5 ± 1.3 | 4.8 ± 0.5 | 9.4 ± 1.1 |
| pH | 7.2 ± 0.8 | 7.2 ± 0.1 | 7.4 ± 1.0 | 7.3 ± 0.1 | 7.3 ± 1.0 | 7.2 ± 0.1 |
| Conductividad (μS cm−1) | 184.0 ± 60.8 | 197.3 ± 87.3 | 203.0 ± 84.2 | 196.8 ± 55.1 | 174.6 ± 28.6 | 187.5 ± 50.0 |
| Temperatura agua (°C) | 21.8 ± 3.1 | 16.4 ± 6.7 | 23.5 ± 3.0 | 17.2 ± 7.0 | 22.8 ± 1.26 | 17.6 ± 6.0 |
| sólidos totales disueltos (mg/L) | 85.8 ± 29.2 | 83.8 ± 46.0 | 97.0 ± 36.0 | 90.5 ± 27.7 | 90.3 ± 13.5 | 86.8 ± 25.8 |
| Ancho medio (cm) | 425 ± 10 | 450 ± 34.6 | 385 ± 64 | 570 ± 175.1 | 570 ± 154.5 | 630 ± 34.6 |
| Profundidad media (cm) | 25.0 ± 5.4 | 29.2 ± 5.9 | 14.4 ± 2 | 23.8 ± 4.3 | 15.7 ± 4.2 | 21.2 ± 9.6 |
| Caudal medio (m³/s) | 0.292 ± 0.17 | 0.590 ± 0.38 | 0.108 ± 0.16 | 0.815 ± 0.43 | 0.365 ± 0.28 | 0.830 ± 0.77 |
Q1-CBN, bosque nativo; Q2-CAG, pastos para actividades agrícolas y ganaderas; Q3-CPF, plantación forestal.
Q1-CBN, native forest, Q2-CAG, pasture for agricultural and livestock activities; Q3-CPF, forest plantation.
Variabilidad taxonómica de macroinvertebrados acuáticos entre quebradas: En los cuerpos hídricos evaluados se recolectaron un total de 6 583 macroinvertebrados acuáticos pertenecientes a dos filos, tres clases, 13 órdenes y 54 familias. En el período seco se recolectaron 2 914 individuos de 52 familias y en la época lluviosa 3 669 de 34 familias (Tabla 2). De manera general el orden Trichoptera con 2 590 individuos (39.3 %) fue el mejor representado, seguido de los órdenes Coleoptera con 1 547 individuos (23.5 %) y Ephemeroptera con 995 individuos (15.1 %). A nivel de familias la más abundante fue Hydropsychidae con 2 001 individuos (30.40 %), seguida de Elmidae con 1 079 individuos (16.4 %).
TABLA 2: TABLE 2: Número total de macroinvertebrados capturados durante las temporadas seca y lluviosa en tres quebradas con diferentes coberturas riparias en el Bosque Protector de Murocomba, Ecuador Total number of macroinvertebrates caught during dry and rainy seasons in three creeks with different riparian cover in the Murocomba Protector Forest, Ecuador
| - | - | Q. La Victoria | - | Q. El Congo | - | Q. La Damita | - |
| - | - | Q1-CBN | - | Q2-CAG | - | Q3-CPF | - |
| Clase/Orden | Familia | Lluvia | Seca | Lluvia | Seca | Lluvia | Seca |
| Blattodea | Blattidae | 0 | 15 | 0 | 4 | 0 | 1 |
| Coleoptera | Dryopidae | 0 | 4 | 0 | 0 | 7 | 10 |
| - | Elmidae | 265 | 165 | 87 | 61 | 227 | 274 |
| - | Hydrochidae | 0 | 3 | 0 | 0 | 0 | 13 |
| - | Hydrophilidae | 0 | 28 | 0 | 5 | 0 | 24 |
| - | Limnichidae | 0 | 0 | 0 | 1 | 0 | 1 |
| - | Lutrochidae | 2 | 2 | 0 | 2 | 0 | 3 |
| - | Psephenidae | 2 | 17 | 53 | 27 | 9 | 10 |
| - | Ptilodactylidae | 87 | 71 | 25 | 11 | 6 | 15 |
| - | Scirtidae | 0 | 0 | 0 | 0 | 0 | 1 |
| - | Staphylinidae | 7 | 14 | 0 | 1 | 5 | 2 |
| Diptera | Blephariceridae | 0 | 0 | 0 | 1 | 0 | 0 |
| - | Ceratopogonidae | 0 | 1 | 0 | 0 | 0 | 0 |
| - | Chironomidae | 0 | 3 | 58 | 60 | 2 | 111 |
| - | Dixidae | 0 | 0 | 0 | 1 | 0 | 3 |
| - | Dolichopodidae | 0 | 2 | 0 | 0 | 0 | 1 |
| - | Empididae | 0 | 0 | 0 | 0 | 0 | 1 |
| - | Muscidae | 0 | 1 | 0 | 0 | 0 | 1 |
| - | Sarcophagidae | 0 | 0 | 0 | 0 | 0 | 1 |
| - | Simuliidae | 4 | 0 | 11 | 56 | 3 | 102 |
| - | Stratiomyidae | 0 | 0 | 0 | 0 | 0 | 1 |
| - | Tabanidae | 0 | 0 | 11 | 0 | 0 | 0 |
| - | Thaumaleidae | 0 | 1 | 0 | 0 | 0 | 0 |
| - | Tipulidae | 0 | 9 | 1 | 10 | 2 | 1 |
| Ephemeroptera | Baetidae | 37 | 6 | 25 | 26 | 47 | 39 |
| - | Leptohyphidae | 23 | 11 | 13 | 68 | 52 | 58 |
| - | Leptophlebiidae | 111 | 44 | 133 | 88 | 165 | 40 |
| - | Oligoneuriidae | 0 | 0 | 0 | 9 | 0 | 0 |
| Hemiptera | Gerridae | 26 | 23 | 23 | 8 | 4 | 1 |
| - | Hebridae | 0 | 0 | 0 | 1 | 0 | 0 |
| - | Hydrometridae | 0 | 0 | 0 | 1 | 0 | 5 |
| - | Naucoridae | 10 | 8 | 19 | 20 | 6 | 8 |
| - | Veliidae | 13 | 2 | 40 | 3 | 7 | 3 |
| Lepidoptera | Crambidae | 0 | 0 | 0 | 0 | 0 | 7 |
| Megaloptera | Corydalidae | 1 | 3 | 12 | 15 | 69 | 25 |
| Odonata | Calopterygidae | 7 | 6 | 0 | 0 | 0 | 0 |
| - | Coenagrionidae | 11 | 8 | 13 | 6 | 8 | 3 |
| - | Libellulidae | 0 | 2 | 17 | 6 | 11 | 9 |
| - | Gomphidae | 12 | 0 | 6 | 0 | 2 | 0 |
| - | Polythoridae | 18 | 11 | 4 | 0 | 0 | 0 |
| Plecoptera | Perlidae | 81 | 99 | 46 | 43 | 8 | 153 |
| Trichoptera | Calamoceratidae | 3 | 13 | 0 | 1 | 0 | 0 |
| - | Glossosomatidae | 0 | 1 | 0 | 3 | 0 | 0 |
| - | Helicopsychidae | 5 | 6 | 10 | 6 | 0 | 0 |
| - | Hydrobiosidae | 13 | 2 | 0 | 6 | 0 | 23 |
| - | Hydropsychidae | 229 | 167 | 175 | 212 | 965 | 253 |
| - | Hydroptilidae | 0 | 0 | 2 | 0 | 0 | 3 |
| - | Leptoceridae | 217 | 129 | 6 | 1 | 6 | 24 |
| - | Odontoceridae | 0 | 0 | 6 | 2 | 2 | 1 |
| - | Philopotamidae | 2 | 2 | 0 | 0 | 62 | 29 |
| - | Polycentropodidae | 0 | 0 | 0 | 1 | 1 | 1 |
| Decapoda | Pseudothelphusidae | 3 | 2 | 1 | 1 | 4 | 0 |
| Isopoda | Porcellionidae | 3 | 4 | 0 | 0 | 0 | 0 |
| Gastropoda | Physidae | 0 | 0 | 0 | 1 | 0 | 0 |
Q1-CBN, bosque nativo; Q2-CAG, pastos para actividades agrícolas y ganaderas; Q3-CPF, plantación forestal
Q1-CBN, native forest, Q2-CAG, pasture for agricultural and livestock activities; Q3-CPF, forest plantation
A nivel de órdenes, se presentaron diferencias en sus abundancias entre las tres quebradas con diferente cobertura de vegetación en sus márgenes. El orden más abundante en las tres fue Trichoptera y aunque la diferencia no fue significativa, la quebrada con cobertura de plantación forestal (Q3-CPF) presentó un valor mayor con un 46.6 % de abundancia, seguida de la quebrada con cobertura de bosque nativo (Q1-CBN) con 38.0 % y la quebrada con cobertura agrícola y ganadera (Q2-CAG) que presentó el menor valor con 27.5 % (Fig. 1A). El orden Coleoptera presentó mayor abundancia en la quebrada Q1-CBN con un 32.1 % y fue significativamente diferente a la quebrada Q2-CAG con un 17.4 % (P = 0.017) y no presento diferencia significativa con la quebrada Q3-CPF, mientras que el orden Diptera fue más abundante en la quebrada con cobertura agrícola y ganadera (Q2-CAG) con un 13.4% y significativamente diferente de Q1-CBN que presentó solamente un 1.0 % (P = 0.022). El orden Hemiptera presentó la menor abundancia en la quebrada Q3-CPF y fue significativamente diferente de Q1-CBN (P = 0.036) y de Q2-CAG (P = 0.041), y el orden Megaloptera presentó la menor abundancia en la quebrada Q1-CBN y fue significativamente diferente de Q2-CAG (P = 0.003) y de Q3-CPF (P = 0.004) (Fig. 1A).
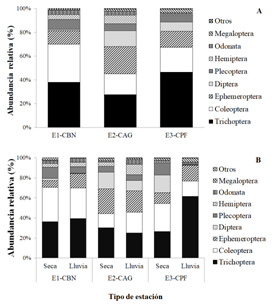
Fig. 1. Fig. 1 Abundancia relativa de órdenes de macroinvertebrados (A) para cada período hidrológico (B) en tres quebradas con diferentes coberturas riparias (Q1-CBN, bosque nativo; Q2-CAG, pastos para actividades agrícolas y ganaderas; Q3-CPF, plantación forestal) en el Bosque Protector de Murocomba, Valencia, Ecuador. Relative abundance of macroinvertebrate orders (A), for hydrological periods (B) in three creeks with different riparian cover (Q1-CBN, native forest, Q2-CAG, pasture for agricultural and livestock activities; E3-CPF, forest plantation) in the Murocomba Protector Forest, Valencia, Ecuador.
Las familias Hydropsychidae, Elmidae, Leptophlebiidae y Perlidae representaron el 62.1 % de la abundancia relativa para el total de individuos colectados en las tres quebradas. El análisis canónico discriminante basado en la abundancia de familias de macroinvertebrados y las variables ambientales separó la quebrada Q2-CAG con uso de suelo agrícola y ganadero y cobertura de pastos en sus márgenes, de las quebradas Q1-CBN con uso de bosque nativo y la quebrada Q3-CPF de plantación forestal y representó el 91.7 % de la varianza en el primer eje canónico. Para la quebrada Q2-CAG, las familias que contribuyeron a la variación fueron principalmente Psephenidae, Chironomidae, Leptophlebiidae, Veliidae, Simuliidae, Baetidae y Leptohyphidae, que se asociaron con los mayores valores de sólidos disueltos totales y pH, así como a mayores valores de IBF y menores valores de los índices EPT y BMWP y de profundidad del cauce (Fig. 2). En las quebradas Q1-CBN y Q3-CPF las familias que más contribuyeron a la variación fueron Leptoceridae, Elmidae, Ptilodactylidae y Philopotamidae, esta última, principalmente en Q3-CPF y se asociaron con mayores valores de los índices EPT y BMWP y con mayor profundidad del cauce (Fig. 2).
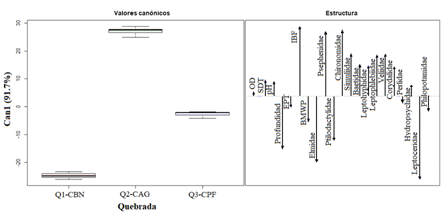
Fig. 2. Fig. 2 Análisis discriminante canónico basado en la abundancia de familias de macroinvertebrados y variables ambientales en tres quebradas con diferentes coberturas riparias (Q1-CBN, bosque nativo; Q2-CAG, pastos para actividades agrícolas y ganaderas; Q3-CPF, plantación forestal) en el Bosque Protector de Murocomba, Valencia, Ecuador. OD, oxígeno disuelto; S.D.T., solidos disueltos totales; EPT, Índice Ephemeroptera, Plecoptera y Trichoptera; IBF, Índice biótico de familias; BWMP, Índice Biological Monitoring Working Party. Discriminant canonical analysis based on macroinvertebrates families abundance and environmental variables in three creeks with different riparian cover (Q1-CBN, native forest, Q2-CAG, pasture for agricultural and livestock activities; Q3-CPF, forest plantation) in the Murocomba Protector Forest, Valencia, Ecuador. OD, dissolved oxygen; S.D.T., total dissolved solids; EPT, Ephemeroptera, Plecoptera and Trichoptera Index; IBF, Biotic Family Index; BWMP, Biological Monitoring Working Party Index.
Variabilidad taxonómica de macroinvertebrados acuáticos entre períodos Hidrológicos: En la Quebrada La Victoria (Q1-CBN) se identificaron 38 familias y se recolectaron un total de 2 077 individuos de macroinvertebrados, de los cuales 885 individuos de 36 familias fueron recolectados en el período seco y 1 192 individuos de 26 familias en período de lluvias (Tabla 1). Los órdenes más abundantes tanto en época seca como en lluvias fueron Trichoptera (36.2 y 39.3 %, respectivamente) y Coleoptera (34.4 y 30.5 %, respectivamente). Se observó un aumento significativo del orden Ephemeroptera en el período de lluvia con un 14.3 %, con relación a la época seca con un 6.9 % (P = 0.031) (Fig. 1B). En época seca las familias más abundantes fueron Hydropsychidae con un 18.9 %, Elmidae con 18.6 % y Leptoceridae con 14.6 %, y en época lluviosa la familia Elmidae fue la más abundante con un 22.2 %, seguida por Hydropsychidae con 19.2 % y Leptoceridae con 18.2 % de individuos (Tabla 2).
Para la Quebrada El Congo de cobertura en pastos y uso agrícola y ganadero (Q2-CAG) se recolectaron 1 565 individuos pertenecientes a 40 familias, de los cuales 768 individuos de 36 familias fueron recolectados en el período seco y 797 individuos de 25 familias en período de lluvias (Tabla 1). El orden más abundante tanto en época seca como en lluvias fue Trichoptera (30.2 y 25.0 %, respectivamente). Se observó una mayor abundancia del orden Ephemeroptera y una menor abundancia de Coleoptera en los dos períodos, con relación a las quebradas Q1-CBN y Q2-CAG. La abundancia del orden Hemiptera que fue significativamente mayor en el período de lluvias (Fig. 1B). En época seca las familias más abundantes fueron Hydropsychidae con un 27.6 %, Leptophlebiidae con 11.5 % y Leptohyphidae con 8.9 %, y en época lluviosa la familia Hydropsychidae fue la más abundante con un 22.0 seguida por Leptophlebiidae con 16.7 % y Elmidae con 10.9 % de los individuos (Tabla 2).
Para la quebrada La Damita (Q3-CPF) se recolectaron 2 941 individuos pertenecientes a 41 familias de macroinvertebrados, de los cuales 1 261 individuos de 39 familias fueron recolectados en el período seco y 1 680 individuos de 25 familias en período de lluvias (Tabla 1). Se destacó el incremento de Trichoptera en la época de lluvia con un 61.7 % de los individuos, presentando diferencia significativa respecto a la época seca en la misma quebrada (P = 0.031) y con los dos períodos hidrológicos de las quebradas Q1-CBN y Q2-CAG (Fig. 1B). Igualmente, se presentó un incremento significativo en el período seco de los órdenes Diptera (P = 0.031) y Plecoptera (P = 0.030) (Fig. 1B). En época seca las familias más abundantes fueron Hydropsychidae con un 20.1 %, Elmidae con 21.7 % y Perlidae con 12.1 %, y en época lluviosa la familia Hydropsychidae fue la más abundante con un 57.4 % seguida por Elmidae con 13.5 % de los individuos (Tabla 2).
El análisis canónico discriminante (ACD) basado en la abundancia de familias de macroinvertebrados y las variables ambientales separó para los dos períodos evaluados (temporada seca y lluviosa) a la quebrada con uso de suelo agrícola, ganadero y pastos en sus márgenes (Q2-CAG), de las quebradas con uso de bosque nativo (Q1-CBN) y de plantación forestal (Q3-CPF). En época seca el ACD representó el 100.0 % de la varianza en el primer eje canónico y para la quebrada Q2-CAG, las familias que contribuyeron a la variación fueron Leptophlebiidae, Chironomidae, Simuliidae, Psephenidae, e Hydropsychidae, que se asociaron con los mayores valores de solidos disueltos totales (SDT), así como a menores valores del índice BMWP (Fig. 3A). En las quebradas Q1-CBN y Q3-CPF las familias que más contribuyeron durante la época seca a la variación fueron Leptoceridae, Ptilodactylidae y Perlidae, y se asociaron con mayores valores del índice BMWP (Fig. 3A). En época de lluvia el ACD representó el 99.0 % de la varianza en el primer eje canónico, separando la quebrada Q2-CAG para la que las familias que contribuyeron a la variación fueron principalmente Psephenidae, Chironomidae, Veliidae, Ptilodactylidae y Leptophlebiidae, que se asociaron con los mayores valores de pH y menores valores de profundidad (Fig. 3B). En las quebradas Q1-CBN y Q3-CPF las familias que más contribuyeron durante la época de lluvia a la variación fueron Hydropsychidae y Corydalidae, principalmente en la quebrada con cobertura de plantación forestal Q3-CPF y Elmidae y Leptoceridae en la quebrada Q1-CBN (Fig. 3B).
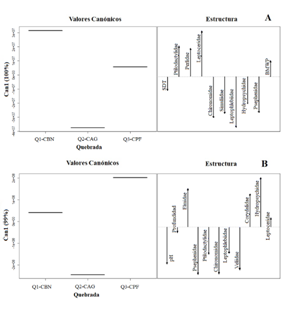
Fig. 3. Fig. 3 Análisis discriminante canónico basado en la abundancia de familias de macroinvertebrados y variables ambientales en el período seco (A) y el período de lluvias (B), en tres quebradas con diferentes coberturas riparias (Q1-CBN, bosque nativo; Q2-CAG, pastos para actividades agrícolas y ganaderas; Q3-CPF, plantación forestal) en el Bosque Protector de Murocomba, Valencia, Ecuador. S.D.T., Solidos Disueltos Totales; BWMP, Índice Biological Monitoring Working Party. Discriminant canonical analysis based on macroinvertebrates families abundance and environmental variables in dry season (A) and rainy season (B), in three creeks with different riparian cover (Q1-CBN, native forest, Q2-CAG, pasture for agricultural and livestock activities; Q3-CPF, forest plantation) in the Murocomba Protector Forest, Valencia, Ecuador. S.D.T., Total Dissolved Solids; BWMP, Biological Monitoring Working Party Index.
Índices biológicos: El índice IBF presentó una calidad de agua muy buena para la quebrada con márgenes con cobertura de bosque nativo (Q1-CBN) y calidad regular a buena para las quebradas con cobertura de pastos (Q2-CAG) y con plantaciones forestales (Q3-CPF) (Fig. 4). Los índices EPT y BMWP presentaron calidad buena para las tres quebradas, aunque la quebrada Q3-CPF presentó mayores valores del Índice EPT y la quebrada Q1-CBN del Índice BMWP (Fig. 4). Por períodos hidrológicos, aunque no se encontró diferencia significativa en el índice EPT, se presentó en el período lluvioso mayor abundancia de estos taxones en las quebradas Q1-CBN y Q3-CPF, mientras que para la quebrada Q2-CAG la abundancia de estos fue menor en dicho período (Tabla 1). Los índices IBF y BMWP tampoco presentaron diferencias significativas entre períodos hidrológicos para cada quebrada.
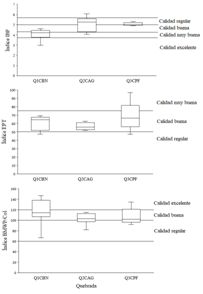
Fig. 4. Fig. 4 Puntuación de IBF, EPT y BWMP para tres quebradas con diferentes coberturas riparias (Q1-CBN, bosque nativo; Q2-CAG, pastos para actividades agrícolas y ganaderas; Q3-CPF, plantación forestal) en el Bosque Protector de Murocomba, Valencia, Ecuador. Las líneas indican umbrales de categoría IBF, EPT and BMWP. IBF, EPT and BWMP score for three creeks with different riparian cover (Q1-CBN, native forest, Q2-CAG, pasture for agricultural and livestock activities; Q3-CPF, forest plantation) in the Murocomba Protector Forest, Valencia, Ecuador. Lines indicate IBF, EPT and BMWP category thresholds.
El índice de Shannon-Weaver fue mayor para la quebrada Q2-CAG con valores entre 2.57 en época seca y 2.59 en época lluvia, sin diferencias significativas entre períodos hidrológicos. Por su parte, las quebradas Q1-CBN y Q3-CPF presentaron mayores valores del índice de Shannon-Weaver en el período seco con relación al período de lluvias (P < 0.001), al igual que mayores valores de dominancia (P < 0.001), en especial en la quebrada Q3-CPF donde el Índice de diversidad disminuyó de 2.52 en seca a 1.59 en lluvia y la dominancia aumentó de 0.12 en seca a 0.36 en lluvia (Tabla 1). Hubo diferencias significativas en el índice de Shannon-Weaver entre las épocas, las coberturas y la interacción entre épocas y coberturas (F = 11.87, P = 0.003; F = 7.50, P = 0.004 y F = 8.60, P = 0.002), respectivamente. Los valores de número de familias y número de taxones raros fueron mayores para las tres quebradas en el período seco (Tabla 1).
Discusión
Se determinó una mejor calidad del agua en la quebrada con cobertura de bosque nativo Q1-CBN, con mayores valores de los índices EPT y BMWP, mientras que los valores más bajos se encontraron en la quebrada con márgenes con uso de suelo agrícola y ganadero de cobertura en pastos Q2-CAG. Esto es similar a lo encontrado por Meza, Rubio, Días, & Walteros (2012) en quebradas de la subcuenca alta del río Chinchiná, en la que la estación con cobertura de pastos presentó menores valores de los índices BMWP y EPT, que las ubicadas en márgenes con cobertura de bosque nativo y de plantaciones forestales, y también con lo reportado por Guerrero-Chuez et al. (2016) quienes determinaron un índice BMWP con aguas de calidad excelente en el uso de suelo de bosque natural a diferencia de los usos de suelo agrícola y de pastos con aguas de calidad regular, en la microcuenca El Sapanal en Cotopaxi, Ecuador. Al respecto, Iñiguez-Armijos et al. (2014) encontraron que la mayoría de las métricas de macroinvertebrados indican una buena calidad del agua cuando el porcentaje de cobertura de vegetación nativa, tanto en la cuenca como en la zona ribereña inmediata, es superior al 70 %, y Galeano-Rendón y Mancera-Rodríguez (2018) encontraron que quebradas con márgenes cubiertos por vegetación nativa presentaron mayores valores de índice BMWP y también mayor promedio de taxones EPT, debido a que dichos ordenes son sensibles a las perturbaciones antrópicas, por lo que suelen estar más asociados a quebradas con zonas ribereñas protegidas. Las quebradas con cobertura de vegetación nativa en sus márgenes presentan ensamblajes más heterogéneos de macroinvertebrados, debido a la abundancia y variedad de hábitats, con una mayor diversidad de taxones intolerantes al estrés ambiental (Bojsen & Jacobsen, 2003; Death & Collier, 2010; Bücker et al., 2010).
En quebradas con mayor disturbio, se espera una disminución de los taxones sensibles como los EPT y un aumento en la importancia de los taxones tolerantes, reduciendo la diversidad (Bispo & Oliveira, 2007). En el presente estudio no se observó una disminución marcada en la riqueza y diversidad entre las tres quebradas, pero si una menor abundancia de taxones EPT en la quebrada con márgenes de cobertura en pastos (Q2-CAG), lo cual se debe a que dichos taxones están formados por un grupo diverso de especies que son sensibles a los cambios ambientales, como la contaminación y la fragmentación del hábitat (Rosenberg & Resh, 1993).
Por su parte, a nivel temporal se observó una diminución del índice de Shannon-Weaver, de la riqueza de familias, así como del número de taxones raros durante el período lluvioso. Durante dicho período, la diversidad disminuyó marcadamente en la quebrada con cobertura de plantaciones forestales Q3-CPF, lo cual se asoció a un incremento del orden Trichoptera, en especial por la dominancia de Hydropsychidae. Igualmente se observó una mayor abundancia de taxones EPT durante el período lluvioso en las quebradas Q1-CBN y Q3-CPF, mientras que para la quebrada Q2-CAG, la abundancia de éstos fue menor. Este incremento de taxones puede estar asociado con la mayor cantidad de refugios y entornos heterogéneos que se presentan en quebradas con márgenes con coberturas de bosque, en las que los aumentos en el caudal pueden ser menos drásticos, y en los que muchos macroinvertebrados pueden protegerse contra el efecto de arrastre de la corriente (Scarsbrook & Townsend, 1993; Townsend, Dolédec, & Scarsbrook, 1997) y también a una mejora en la calidad del agua en el período lluvioso, que se evidenció en un mayor valor de los índices EPT y BMWP, debido a que el aumento de caudal permite una mayor reducción de la concentración de materia orgánica. Esto último, coincide con lo reportado por Bispo, Oliveira, Bini, y Sousa (2006) quienes encontraron un pico de abundancia de macroinvertebrados al comienzo de la temporada de lluvias asociada con una mejora de calidad de agua. Estos autores destacan que probablemente, los efectos positivos de la dilución de contaminantes observados se pierden con mayores tasas de precipitación e incremento de caudal, lo cual también puede explicar la disminución en taxones EPT durante el período lluvioso en la quebrada Q2-CAG ya que ésta, fue la única en la que se presentó un aumento significativo del caudal. Bispo et al. (2006) destacan que la precipitación explicó una parte importante de la variación temporal en la abundancia de EPT, especialmente en sitios de mayor caudal con una disminución en la abundancia durante la temporada de lluvias.
Esta disminución en taxones EPT en la quebrada de cobertura agrícola y ganadera (Q2-CAG) coincide con lo reportado por Schmitt, Siegloch, da Silva, Lisboa, y Petrucio (2016) quienes también encontraron una baja abundancia de EPT durante los períodos de alta precipitación probablemente causada por un aumento en el flujo de agua, lo que produce un consecuente efecto de deriva de los individuos, la remoción de insectos y una reducción de su abundancia local (Flecker & Feifarek, 1994), así como con otros estudios en quebradas Andinas, que también reportan una menor abundancia del ensamblaje de macroinvertebrados en el período lluvioso (e.g. Jacobsen & Encalada, 1998; Ramírez, Pringle, & Douglas, 2006; Ríos-Touma et al., 2011; Ríos-Pulgarín et al., 2016a).
Vásquez-Velásquez (2016), encontró en cuencas de cabecera de los Andes centrales de Colombia, que la cuenca bajo cobertura de bosque natural presentó un régimen de caudales estable y de poca variación con relación a las cuencas con cobertura de plantación forestal y cobertura en pastos, revelando una mayor inestabilidad en el régimen de caudales en éstas últimas. Así mismo la cobertura boscosa ofrece mejores condiciones de regulación de los caudales, por lo que la deforestación tiende a generar caudales máximos y mínimos más extremos (Bruijnzeel, 2004; Licata, Gyenge, Fernández, Schlichter, & Bond, 2008; Vanclay, 2009; Vásquez-Velásquez, 2016), como se observó en la quebrada Q2-CAG, que fue la única con diferencias significativas de caudal entre los períodos seco y lluvioso.
El orden Trichoptera, y la familia Hydropsychidae, presentaron la mayor abundancia relativa para el total de individuos recolectados en las tres quebradas. Este resultado es similar a lo reportado por Jaimes-Contreras y Granados-Martínez (2016), en siete afluentes que drenan de la Sierra Nevada de Santa Marta (Colombia), y coincide también con la mayor abundancia del orden Trichoptera y la familia Hidropsychidae en el Bosque Protector Río Guajalito en Ecuador (Arroyo & Encalada, 2009). La mayor abundancia de Hydropsychidae se puede relacionar con que es una familia muy tolerante, con especies segregadas dentro de diferentes características de calidad del agua a lo largo de los ríos (Bonada, Zamora-Muñoz, Rieradevall, & Prat, 2004), y ha sido reportada como la familia más abundante en las regiones tropicales (Flint, Holzenthal, & Harris, 1999). El amplio rango de distribución de la familia Hidropsychidae, está asociado a su alta tolerancia a cambios ambientales y su capacidad para construir redes que le permiten fijarse en el sustrato y capturar gran cantidad de alimento (López-Delgado, Vásquez-Ramos, & Reinoso-Flórez, 2015).
Se observó una disminución del orden Tricoptera en la quebrada de cobertura de pastos (Q2-CAG), lo que coincide con lo encontrado por Henriques-Oliveira, Nessimian, y Baptista, (2015), quienes observaron una pérdida de diversidad en las quebradas sin vegetación arbórea ribereña en comparación con las quebradas con cobertura de bosque. El orden Trichoptera en sistemas lóticos se ha asociado con una alta cobertura vegetal en sus márgenes, y se ha establecido que el aumento de zonas deforestadas por actividades agropecuarias puede causar cambios ambientales y reducir su diversidad (Benstead, Douglas, & Pringle, 2003; Lorion & Kennedy, 2009; Breda et al., 2018). Esta menor abundancia y diversidad de Trichoptera en sitios deteriorados está relacionada con la pérdida de hábitats por procesos como la deforestación y el cambio de usos de suelo para actividades productivas, con el consecuente aumento de la sedimentación que disminuye la heterogeneidad del hábitat (Allan, Erickson, & Fay, 1997) y la reducción de la entrada de material alóctono al cuerpo hídrico (e.g. Townsend, Dolédec, & Scarsbrook, 1997; Bispo & Oliveira, 2007; Bücker et al., 2010; Giraldo et al., 2014).
Para la quebrada Q2-CAG se observó una menor abundancia del orden Coleoptera y un incremento de los órdenes Diptera (e.g. Chironomidae y Simulidae) y Ephemeroptera (e.g. Leptophlebiidae y Baetidae), mientras que la quebrada Q1-CBN presentó mayores abundancias de Coleoptera (e.g. Elmidae y Ptilodactylidae). Este patrón coincide con lo encontrado por Galeano-Rendón y Mancera-Rodríguez (2018) quienes compararon quebradas con buena cobertura de bosque natural con quebradas sometidas a diferentes afectaciones en su cobertura por procesos de deforestación asociada con actividades agrícolas y ganaderas. Igualmente, una mayor abundancia de Ephemeroptera ha sido reportada en quebradas ubicadas en áreas deforestadas y dominadas por matrices de pastizales por Bücker et al. (2010) en el sur de Ecuador, y en Costa Rica por Lorion y Kennedy (2009), y una mayor abundancia de Díptera fue encontrada por Giraldo et al. (2014) en estaciones con intervención antrópica por ganadería en quebradas en Colombia. La mayor abundancia de Coleoptera, en particular de las familias Elmidae y Ptilodactylidae en la quebrada con cobertura de bosque natural, puede estar asociada a la mayor disponibilidad de materia orgánica gruesa y a la mayor entrada de material alóctono, ya que estas familias se han asociado con la abundancia de detritus de la hojarasca (Bojsen & Jacobsen, 2003).
La quebrada con cobertura de bosque nativo mostró una mayor calidad hídrica y diversidad de macroinvertebrados sensibles a las perturbaciones antrópicas que las quebradas con coberturas de pastos y de plantaciones forestales. Los cambios en la composición y estructura del ensamblaje de macroinvertebrados en estas tres quebradas con diferentes coberturas riparias indican que la pérdida de vegetación nativa representa un importante impacto antrópico con efectos en la estructura de las comunidades acuáticas. En el bosque protector Murocomba la pérdida de la vegetación nativa, las malas prácticas agrícolas y el aumento de la población han modificado los usos de suelo, alterando la cobertura del suelo en los márgenes de las quebradas, la calidad hídrica y la estructura de los ensamblajes de insectos acuáticos. Por lo anterior, se recomienda el mantenimiento y restauración de la vegetación ribereña con bosques nativos, para mitigar los efectos de la deforestación en los patrones y procesos de las comunidades acuáticas.

