











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
La motivación y la activación sexual son dos procesos semi-independientes necesarios para la interacción copulatoria (Pfaus, Kippin, & Coria-Avila, 2003). La motivación sexual es un término que se refiere al ímpetu que surge de la estimulación interna y/o externa para buscar establecer la interacción sexual y es medida por la disposición de los sujetos para gastar energía y tener acceso a la potencial pareja sexual (Sachs, 2000). Para que ocurra este proceso, es necesario que los machos cumplan con dos condiciones: por una parte, que presenten un medio hormonal interno propicio; por otra, que se lleve a cabo un adecuado procesamiento de los estímulos sexuales procedentes de la potencial pareja (Hernández-González & Guevara, 2009). Este estado “sexualmente motivado” lo inducirá a realizar una serie de conductas propias de la interacción sexual (acercamiento a la potencial pareja, estimulación corporal y genital) que redundarán en un estado de activación sexual asociado al proceso de erección peneana; de esta manera, los machos se encontrarán en un estado fisiológico adecuado para iniciar la fase consumatoria de la conducta sexual (Masters & Johnson, 1966;Ågmo, 2011).
Sachs (2000)propone que la erección peneana es la “medida de oro” de la activación (arousal). Así, la activación sexual ha sido ligada con la respuesta genital, de tal manera que si un macho tiene una erección, se dice que está sexualmente activado. Es posible que un macho realice conductas de cortejo con tal de tener acceso a una potencial pareja sin presentar erección genital y, por tanto, los eventos de motivación y activación sexual aparecen desfasados.Bancroft (2005)considera la activación sexual como “un estado motivado hacia la experiencia de placer sexual”, involucrando procesamiento de estímulos relevantes, activación generalizada, motivación de incentivo y respuesta genital. Si bien se han reportado múltiples definiciones para estos conceptos, a la fecha aún no se ha logrado dilucidar si son procesos independientes o si estos guardan una interrelación temporal que es necesaria para lograr una interacción sexual adecuada.
Por otra parte, los modelos animales se han utilizado para tener un acercamiento a diferentes fenómenos fisiológicos y poder estudiar mecanismos biológicos en el humano, por ejemplo, la depresión (Sequeira-Cordero & Fornaguera-Trías, 2014), la ansiedad y el miedo (Mora-Gallegos & Salas-Castillo, 2014). Dichos modelos pretenden cumplir con criterios de homología con las conductas que se estudian en el humano; en este sentido, las conductas de la interacción sexual de la rata, como modelo, cumplen con los criterios de correlación y de homología con la conducta sexual del humano (Ågmo, 2017). Por ejemplo, el hombre realiza conductas de aproximación a una mujer que le resulta sexualmente atrayente con la intención de conseguir la cópula con esta; efectivamente, la rata macho, despliega también conductas de acercamiento hacia una hembra receptiva previas a la realización de montas, intromisiones y eyaculaciones que vienen acompañadas de acicalamiento genital (AG, auto-acicalamiento de testículos y pene) (Meisel & Sachs, 1994). El tiempo que tarda el macho en presentar las respuestas de AG, la primera monta y la primera intromisión, junto con las conductas de atención a la hembra han sido consideradas como índices de motivación sexual (Paredes, Highland, & Karam, 1993; Meisel & Sachs, 1994;Hernández-González, 2000;Retana-Márquez, Bonilla-Jaime, Vázquez-Palacios, Martínez-García, & Velázquez-Moctezuma, 2003).
A nivel del laboratorio, se han podido diferenciar de manera precisa las conductas indicadoras de motivación y de activación sexual utilizando paradigmas que restringen el contacto físico con la rata hembra receptiva al colocar entre ambos sujetos una división de acrílico transparente con orificios. En este paradigma, el AG y los toques de narina (TN, cuando el macho coloca las narinas en los orificios del panel de acrílico) que el macho despliega al percibir los estímulos visuales, olfatorios y auditivos de la hembra inaccesible (Paredes, 2009), constituyen los parámetros de motivación sexual, en tanto que las erecciones peneanas sin contacto (EPsC), definidas como aquellas erecciones que ocurren fuera del contexto de la cópula y que son generadas por la percepción de estímulos olfativos, odoríferos y visuales emitidos por la hembra receptiva inaccesible, constituyen la conducta indicadora de activación sexual del macho (Sachs, 1997).
Se ha descrito que la conducta sexual es afectada por diferentes factores, tales como cambios hormonales (Manzo, Cruz, Hernández, Pacheco, & Sachs, 1999), estrés (Hernández-González et al., 2017) y el consumo de diversas sustancias psicoactivas (SPA); entre ellas, el alcohol (Pfaus & Pinel, 1989). El alcohol es una SPA de abuso legal de alto consumo, cuyos efectos a nivel fisiológico dependen en gran medida de la dosis, tipo de alcohol y tiempo de consumo, así como de la edad y género de quien lo consume (McKim, 1996). En el humano, existen discrepancias respecto al efecto del alcohol sobre la conducta sexual. Por una parte, se ha descrito que su consumo va asociado con una desinhibición conductual (puede ser en ambientes sociales o no, igual ocurrirá), acompañada de una mayor activación hacia estímulos sexuales (Wilson, 1981); mientras que, otros reportes muestran que la concentración de alcohol en sangre es inversamente proporcional al grado de tumescencia peneana (Farkas & Rosen, 1976). Tales discrepancias pudieran deberse a que la mayoría de estudios han sido efectuados en personas alcohólicas crónicas en las cuales es difícil controlar variables tales como la dosis, temporalidad de consumo y la edad de los participantes (Wilson, Lawson, & Abrams, 1978;George & Stoner, 2000).
Considerando que estas variables (dosis, temporalidad y edad) pueden ser controladas en los modelos animales, se ha reportado que la administración crónica de dosis altas de alcohol en ratas desencadena anormalidades anatómicas de estructuras reproductivas, como una reducción en el músculo liso peneano (Saito et al., 1994;Gan, Shi, Liu, An, & Wang, 2007), disminución en los niveles hormonales de la testosterona, deficiencias en los parámetros espermáticos (Cicero et al., 1990) y una deficiente ejecución sexual, manifestada por mayores latencias de monta y eyaculación (Pfaus & Pinel, 1989). En otro estudio,Hernández-González et al. (2004)reportaron un adelanto en la pubertad, manifestado por un pico de ocurrencia más temprano de AG y EPsC en ratas macho prepúberes que consumieron alcohol de manera crónica desde el destete hasta la pubertad.
Tomando en cuenta los antecedentes anteriores, en este trabajo se utilizó a la rata como modelo animal para evaluar los estados de motivación y activación sexual, así como su sensibilidad a los efectos de diferentes dosis de alcohol. El modelo animal utilizado en este estudio permitió brindar información más objetiva del efecto del alcohol sobre los componentes apetitivos de la conducta sexual, teniendo dos principales ventajas; una, poder controlar las dosis, la vía de administración y la temporalidad de administración del alcohol; y dos, utilizar un paradigma que permite evaluar de manera independiente los parámetros indicadores de motivación y activación sexual en la rata macho.
Metodología
Animales
Se utilizaron ratas machos sexualmente expertas (n= 56) de la cepa Wistar (250 - 300 g de peso), tomadas del bioterio del Instituto de Neurociencias, de la Universidad de Guadalajara, los cuales fueron hospedados 4 por caja-habitación (jaula estándar de laboratorio), en un cuarto con luz controlada en un ciclo de 12:12 horas luz: oscuridad (la luz se encendía a las 8:00 p.m.), con temperatura mantenida en un rango entre 22-23 °C. La comida y el agua estuvieron disponiblesad libitum. El cuidado de los animales, así como todos los procedimientos, fueron aprobados por nuestro Comité Institucional de cuidado y manejo animal de acuerdo con la guía del Comité Institucional del Instituto Nacional de abuso de drogas, el Instituto Nacional de salud, y la guía para el cuidado y uso de animales de laboratorio (Institute of Laboratory Animal Resources, Commission on Life Science, National Research Council, 1996).
Todos los sujetos fueron considerados sexualmente expertos cuando eyacularon en menos de 15 minutos en al menos 2 de 3 pruebas de conducta sexual. Dichas pruebas se realizaron una cada tercer día y consistían en permitirle al macho tener interacción con una hembra receptiva. En cada prueba, los machos eran colocados en una arena de prueba plexiglás (50 cm x 25 cm x 30 cm) por cinco minutos antes de introducir a una hembra receptiva que servía como estímulo sexual. Una vez que la hembra estaba con el macho en la arena, se le permitía tener interacción sexual hasta la eyaculación. La prueba se dio por terminada si el macho no conseguía la intromisión en 15 minutos o bien, si no conseguía la eyaculación en 15 minutos posterior a la primera intromisión. Las hembras fueron inducidas a la receptividad sexual con la administración secuencial de valerianato de estradiol (5 mg/0.05 ml, 44 horas antes de la prueba) seguido de progesterona (1 mg/0.05 ml, 6 horas antes de la prueba).
Administración de tratamientos
Los machos sexualmente expertos fueron divididos en 4 grupos (n= 14/grupo) dependiendo del tratamiento al que fueron asignados: SAL (solución salina); 0.75 g/kg etanol; 1.0 g/kg etanol; 1.25 g/kg etanol. Las diferentes dosis de etanol (99.8%, MERK, Germany) fueron administradas por vía intraperitoneal (i.p.) en un volumen de (0.5 ml) como una solución en salina fisiológica para minimizar las diferencias inducidas por la concentración en la tasa de absorción del alcohol, así como la irritación del tejido en el sitio de inyección. Posterior a la inyección, todos los animales fueron devueltos a su caja-habitación y, después de 15 minutos, se colocaron en la caja de motivación y activación sexual para evaluar la latencia, frecuencia y duración del acicalamiento genital (AG), de los toques de narina (TN) y de las erecciones peneanas sin contacto (EPsC).
Procedimiento
Las evaluaciones se realizaron en la caja de motivación y activación sexual, la cual está construida con plexiglás (60 cm x 30 cm x 50 cm) y dividida en dos compartimientos iguales por un panel de acrílico transparente con orificios de 1 cm de diámetro. La hembra se debe colocar en un compartimiento (cada compartimiento medía 30 cm x 30 cm x 50 cm) y el macho se coloca en el compartimiento contiguo, de tal manera que este puede ver, oler y escuchar a la hembra receptiva, pero no tiene acceso físico a ella. Para poder observar apropiadamente la parte ventral de los sujetos, la caja de registro posee un espejo por debajo del piso transparente colocado con un ángulo de 45°.
Quince minutos después de la administración de los diferentes tratamientos, los machos fueron colocados en su respectivo compartimiento durante 5 minutos para su habituación sin ningún estímulo y, posterior a este periodo de habituación, se colocó una hembra en el mismo compartimiento del macho, al cual se le permitió tener una intromisión con esta para inducirlo a un estado sexualmente motivado. Inmediatamente después, la misma hembra se pasó al compartimiento adyacente y se realizó el registro conductual, durante 15 minutos. Los parámetros considerados fueron: a) latencia de AG, medido en segundos, desde la intromisión con la hembra receptiva hasta la ocurrencia del primer AG; b) frecuencia de AG, número de veces que el macho realizó AG, durante los 15 minutos de registro; c) duración de AG, tiempo total en segundos en que el macho realizó AG; a) latencia de TN, tiempo en segundos desde la intromisión con la hembra receptiva hasta que el macho presentó el primer TN; e) frecuencia de TN, número de veces que el macho realizó TN durante 15 minutos de registro; f) duración de TN, tiempo total en segundos en que el macho realizó TN; g) latencia de EPsC, tiempo en segundos desde la intromisión con la hembra receptiva hasta que el macho presentó la primera EPsC; h) frecuencia de EPsC, número de veces que el macho realizó EPS durante 15 minutos de registro; i) duración de EPsC, tiempo total en segundos en que el macho realizó EPsC.
Análisis estadístico
Se comparó el número de sujetos por grupo que presentó cada conducta utilizando la prueba de x 2 y se consideró unap≤ .05 como significativa. Además, se utilizó un análisis de varianza (ANDEVA) de una vía para comparar el efecto de los tratamientos sobre la latencia, frecuencia y duración de AG, TN y EPsC (se realizó un análisis por cada parámetro evaluado), seguido de la prueba de Tukey como pruebapost hoc. Se consideró unap≤ .05 como significativa. En el caso de las latencias, solo los machos que presentaron la conducta fueron considerados para el análisis.
Resultados
Proporción de sujetos que presentaron cada conducta
Todos los sujetos de los cuatro grupos presentaron las conductas de AG y TN. Solo en el caso de las EPsC, un menor número de sujetos del grupo que recibió la dosis de 1.25 g/kg (3 de 14 sujetos, 21.4%) presentaron EPsC respecto al grupo 0.75 g/kg (14 de 14 sujetos, 100%) [x 2 = 18.12;p≤ .00002] y SAL (9 de 14 sujetos, 64.3%) [x 2 = 5.25; p ≤ .02195]. Similarmente, un menor número de sujetos del grupo 1.0 g/kg (7 de 14 sujetos, 50%) presentaron EPsC respecto al grupo 0.75 g/kg [x 2 = 9.33;p≤ 0.00225]. Contrariamente, un mayor número de sujetos del grupo 0.75 g/kg exhibieron EPsC comparado con SAL [x 2 = 6.09;p≤ .01362]. No se encontraron diferencias significativas en las comparaciones entre SAL y 1.0 g/kg [x 2 = .58;p≤ .44501], así como entre 1.0 g/kg y 1.25 g/kg [x 2 = 2.49;p≤ .11465].
Acicalamiento genital (AG)
El análisis de ANDEVA reflejó diferencias significativas entre grupos [F(3.52)= 3.0;p≤ .0388] en la latencia a la primer ocurrencia de AG. El análisispost hocmostró que el grupo 1.25 g/kg presentó una mayor latencia de AG a comparación con los grupos SAL y 0.75 g/kg (p≤ .05) (Figura 1A). También, se encontraron diferencias significativas entre grupos [F(3.52)= 2.76;p≤ .0512] en la frecuencia de AG, de manera que el análisispost hocmostró que el grupo 1.25 g/kg presentó una menor frecuencia de AG respecto a todos los demás grupos (p≤ 0.05) (Figura 1B). En la duración de este parámetro el análisis de ANDEVA no mostró diferencias significativas entre grupos [F(3.52)= 2.16;p≤ .1035] (Figura 1C).
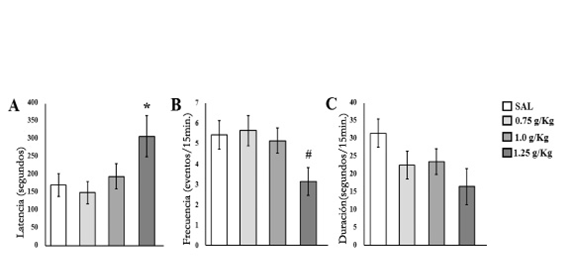
Figura 1 Media ± error estándar de A) latencia, B) frecuencia y C) duración del acicalamiento genital presentado por los sujetos de los diferentes grupos. *p ≤ .05 mayor respecto a SAL y 0.75 g/kg; # p ≤ .05 menor respecto a SAL, 0.75 g/kg y 1.0 g/kg.
Toques de narinas (TN)
En la latencia a la primer ocurrencia de TN, el análisis de ANDEVA no mostró diferencias significativas entre grupos [F(3.52)= 0.82;p≤ .4933] (Figura 2A). En contraste, claras diferencias significativas entre grupos fueron detectadas en la frecuencia [F(3.52)= 15.97;p≤ .0001]. Los sujetos del grupo 1.25 g/kg presentaron un menor número de TN respecto a SAL y 0.75 g/kg (p≤ 0.01), además el grupo 1.0 g/kg mostró una menor frecuencia de esta conducta comparado con SAL (p≤ .05) (Figura 2B). En la duración también se encontraron diferencias significativas entre grupos [F(3.52)= 24.44;p≤ .0001]; el análisispost hocmostró que 1.25 g/kg presentó una menor duración de TN comparado con los otros tres grupos (p≤ 0.01), además 1.0 g/kg mostró una menor duración de esta conducta comparada con SAL (p≤ .01) (Figura 2C).
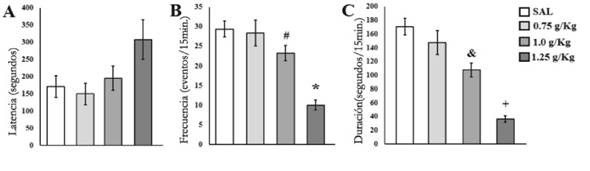
Figura 2 Media ± error estándar de A) latencia, B) frecuencia y C) duración de los toques de narina presentados por los sujetos de los diferentes grupos. *p ≤ .01 menor respecto a SAL y 0.75 g/kg; #p ≤ .05 menor respecto a SAL; +p ≤ .01 menor respecto a SAL, 0.75 g/kg y 1.0 g/kg; & p ≤ .01 menor respecto a SAL.
Erecciones peneanas sin contacto (EPsC)
Para la latencia de la ocurrencia de la primer EPsC, el análisis de ANDEVA se realizó solo con los sujetos que presentaron EPsC (SAL = 9, 0.75 g/kg=14, 1.0 g/kg = 7, 1.25 g/kg = 3) y no se reflejaron diferencias significativas entre grupos [F(3.29)= 0.82;p≤ .4933] (Figura 3A). En la frecuencia de este parámetro sí se encontraron diferencias significativas entre grupos [F(3.52)= 4.46;p≤ .0073]; el análisis post hoc mostró que los sujetos del grupo 1.25 g/kg presentaron una menor frecuencia de EPsC respecto a los del grupo SAL (p≤ .05) y 0.75 kg/g (p≤ .01), también 1.0 g/kg exhibió un menor número de EPsC comparado con 0.75 g/kg (p≤ .01) (Figura 3B). En la duración de este parámetro, el análisis de ANDEVA mostró diferencias significativas entre grupos [F(3.52)= 5.76;p≤ .0018], el análisispost hocindicó que 1.25 g/kg presentó una menor duración de EPsC comparado con SAL (p≤ .05) y con 0.75 g/kg (p≤ 0.01), también 1.0 g/kg exhibió una menor duración de EPsC comparado con 0.75 mg/kg (p≤ 0.01) (Figura 3C).
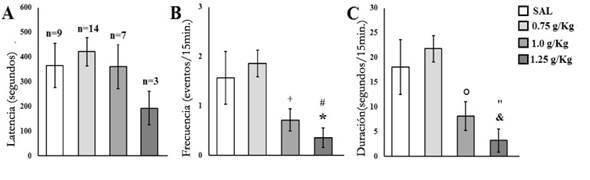
Figura 3 Media ± error estándar de A) latencia, B) frecuencia y C) duración de las erecciones peneanas sin contacto presentadas por los sujetos de los diferentes grupos. En la gráfica A se indica el número de sujetos de cada grupo que presentaron la conducta. *p ≤ .05 menor respecto a SAL; #p ≤ .05 menor respecto a 0.75 g/kg; +p ≤ .01 menor respecto a 1.0 g/kg; &p ≤ .05 menor respecto a SAL; “p ≤ .01 menor respecto a 0.75 g/kg; °p ≤ .01 menor respecto a 0.75 g/kg.
Discusión
En este trabajo, aprovechando las bondades de la rata como modelo de la conducta sexual y utilizando un paradigma experimentalad hoc, es que fue posible analizar de manera independiente la sensibilidad a diferentes dosis de alcohol que presentan los parámetros indicadores de motivación sexual; el AG como respuesta de estimulación genital ante los estímulos de la hembra inaccesible y los TN como signo de atención y aproximación a la hembra receptiva. También, se analizó el parámetro indicador por excelencia de activación sexual, las EPsC.
Uno de los aspectos a controlar en este estudio fue descartar que los resultados pudieran deberse a la inhibición psicomotora inducida por el efecto del alcohol, por lo que en un estudio piloto se evaluaron los efectos de las dosis utilizadas sobre la actividad motora en campo abierto (15 minutos después de la administración de alcohol i.p., se colocaron a las ratas en una caja de plástico de 55 cm x 55 cm x 60 cm, con el piso dividido en 25 cuadros de 11 cm por lado, cada uno). En este piloto no se encontraron diferencias en el número de cuadros cruzados por los sujetos de los diferentes grupos (p= .94, datos no mostrados). Estos resultados coinciden con otros estudios en los que se administraron 0.5 g/kg (Scott et al., 1994), 0.75 g/kg (Varlinskaya & Spear, 2002) y 1 g/kg de alcohol via i.p. en ratas (Hart, 1969;Scott et al., 1994) y no observaron alteraciones en la coordinación motora. Por el contrario, se han descrito incrementos en la actividad social (por ende, de actividad motora) con dosis de 0.5 y 0.75 g/kg vía i.g. (Varlinskaya et al., 2001;Varlinskaya & Spear, 2002). Dado lo anterior, es improbable que los cambios encontrados en el presente estudio puedan resultar de afectaciones motoras inducidas por el alcohol.
En términos generales, se encontró un efecto diferencial dosis-dependiente del alcohol sobre la motivación y activación sexual de la rata macho, dando evidencia de la diferente sensibilidad al alcohol que tienen estos procesos. Las dosis alta (1.25 g/kg) y moderada (1.0 g/kg) de alcohol afectaron drásticamente ambos procesos fisiológicos, mientras que la dosis baja (0.75 g/kg), por el contrario, tuvo un efecto facilitador solo sobre la activación sexual sin alterar la motivación sexual.
Los machos tratados con la dosis más alta (1.25 g/kg) mostraron una mayor latencia y menor frecuencia de AG, así como una menor frecuencia y duración de TN. Estos resultados son similares a los reportados en otros estudios donde se evaluó el efecto del alcohol sobre la conducta sexual, que indicaba que las ratas tratadas con dosis mayores a 1 g/kg tardaron más en presentar las respuestas copulatorias, mientras que dosis bajas no alteraron tales parámetros (Hart, 1969;Pfaus & Pinel, 1989).
La motivación y activación sexual dependen del adecuado funcionamiento de estructuras del sistema dopaminérgico mesocorticolímbico (Hull, Muschamp, & Sato, 2004), tales como la corteza prefrontal (CPF) (Ågmo, Villalpando, Picker, & Fernández, 1995) y el núcleo accumbens (NAcc) (Liu, Sachs, & Salomone, 1998), entre otros. Efectivamente, una prevalencia de frecuencias electroencefalográficas (EEG) de 4 a 13 Hz (banda theta) en estas estructuras durante el estado sexualmente motivado de ratas macho ha sido reportado (Hernández-González, Guevara, & Ågmo, 2014). De manera interesante, se mostró que la administración i.p. de 0.75 g/kg de alcohol indujo una menor sincronización de la banda theta entre los NAcc izquierdo y derecho durante el estado vigilia-quieto, lo cual indica el efecto del etanol sobre el funcionamiento de estas estructuras que participan en el adecuado procesamiento del valor incentivo de los estímulos (Hernández-González, Martínez-Pelayo, Arteaga-Silva, Bonilla-Jaime, & Guevara, 2009).
Un proceso que es necesario para el despliegue de las conductas motivadas es que los sujetos presenten niveles adecuados de atención a los estímulos incentivos; en este sentido, se ha reportado que dosis altas de etanol (mayores a 0.75 g/kg) alteran la ejecución de pruebas de detección visual en ratas, las cuales presentan un menor porcentaje de ensayos correctos, demostrando que el alcohol altera la atención sostenida (Rezvani & Levin, 2003). Particularmente, en el contexto de la conducta sexual, se ha descrito que la CPF y la corteza parietal podrían participar en la atención e identificación de la hembra como un incentivo sexual.Guevara et al. (2014)reportaron que, tras la administración intraperitoneal de etanol, las ratas macho presentaron una mayor latencia de monta e intromisión, lo que se asoció con una menor correlación EEG de frecuencias rápidas (14-30 Hz) entre áreas prefronto-parietales.
Se ha reportado una prevalencia de frecuencias rápidas durante procesos atencionales en respuesta a estimulación sensorial (Murthy & Fetz, 1992). Por tanto, en este estudio, es probable que las dosis moderada (1.0 g/kg) y alta (1.25 g/kg) hayan afectado la funcionalidad de la CPF, de tal manera que disminuyó la atención a la hembra receptiva. Esta aseveración es apoyada por el hecho de que las ratas tratadas con estas dosis de alcohol presentaron una menor duración y frecuencia de TN (indicadores de atención y aproximación a la hembra receptiva); así, es probable que, al no prestar la suficiente atención a la hembra receptiva, no lograron alcanzar una adecuada motivación sexual.
Por otra parte, se ha descrito que las hormonas esteroideas, como la testosterona y sus metabolitos, regulan tanto el AG (Moore, 1986) como a las EPsC (Manzo et al., 1999). Existen varios trabajos en los que se ha demostrado que el alcohol afecta los niveles de testosterona en ratas; por ejemplo,Badr, Bartke, Dalterio y Bulger (1977)reportaron que el alcohol administrado i.p. a dosis de 1.24 g/kg en ratones adultos, provocó un decremento de testosterona. La dosis que reportaron estos autores (1.24 g/kg) es semejante a la utilizada en el presente trabajo (1.25 g/kg), donde se encontró que las ratas tratadas con dosis altas de alcohol presentaron una menor frecuencia de AG y se vieron suprimidas las EPsC (solo 3 de 14 sujetos las presentaron). Por lo tanto, es probable que el alcohol a esta alta dosis ejerciera un efecto inhibitorio sobre la liberación de testosterona, afectando así la ocurrencia del AG y las EPsC.
Si bien, las ratas tratadas con la dosis baja de alcohol no mostraron diferencias significativas respecto al grupo tratado con solución salina (solo se observó una tendencia a presentar mayor frecuencia y duración de EPsC respecto al grupo salina), si exhibieron una mayor frecuencia y duración de EPsC respecto a los otros grupos tratados con alcohol; además fue el grupo 0.75 g/kg el que mostró un mayor número de sujetos que presentaron EPsC (14 de 14 sujetos lo presentaron). Estos datos sugieren que, a dosis bajas el alcohol, produce un efecto facilitador sobre las EPsC, dato que es apoyado por otro estudio en el que ratas tratadas con dosis bajas de alcohol (0.5 g/kg), un mayor porcentaje de sujetos presentaron monta y eyaculación aun cuando las hembras no estaban receptivas a comparación del grupo control y tratados con dosis moderadas (1.0 g/kg), los cuales no presentaron dichas conductas con hembras no receptivas (Pfaus & Pinel, 1989). Estos hallazgos tomados en conjunto muestran que dosis bajas y altas de alcohol ejercen un efecto diferencial sobre la activación sexual; es decir, dosis bajas asociándose a un efecto facilitador sobre la inducción de EPsC y dosis altas ejerciendo un efecto supresor sobre la inducción de este parámetro indicador de activación sexual.
De los pocos estudios que han evaluado el efecto del alcohol sobre las EPsC, destaca el efectuado porHeaton &Varrin (1991).Tales autores reportaron que las ratas tratadas con dosis mayores a 1 g/kg de alcohol mostraron una menor frecuencia de EPsC, mientras que dosis menores no mostraron efectos sobre este parámetro, lo cual coincide con nuestros datos; es decir, dosis altas de alcohol disminuyen la frecuencia y duración de EPsC.
Por otra parte, se ha mostrado la participación del sistema de neurotransmisión dopaminérgico en el inicio y mantenimiento de las erecciones peneanas (Pfaus & Phillips, 1991). Asimismo, se ha descrito que dosis bajas de alcohol producen un aumento en los niveles de dopamina en el NAcc, mientras que dosis altas y el consumo crónico de alcohol producen un decremento de este neurotransmisor (Lewis & June, 1990;Mcbride, Murphy, Lumeng & Li, 1990;Weiss & Porrino, 2002;McBride et al., 1990;Olsen, 2011;Vezali-Costardi et al., 2015). Con base en lo anterior, se podría pensar que en las ratas que recibieron una dosis baja de alcohol (0.75 g/kg) haya ocurrido un aumento de los niveles de dopamina en el NAcc, de tal manera que se vio facilitada la inducción de EPsC; mientras que las ratas tratadas con dosis altas de alcohol pudieron presentar una disminución en los niveles de dopamina, por lo que se perjudicó la inducción de EPsC; sin embargo, estas propuestas requerirán de estudios posteriores.
Son pocos los trabajos en humanos donde se han evaluado los efectos del alcohol sobre la activación sexual. Por ejemplo,George et al. (2006)pidieron a los participantes que imaginaran situaciones eróticas o sexuales que generalmente les permitían experimentar erección peneana (activación sexual instruccional). Mediante un pletismógrafo peneano, midieron el aumento de circunferencia peneana y encontraron que tras la administración de 1.25 ml de etanol/kg de peso, los participantes no fueron capaces de generar una erección peneana mediante la imaginación, a diferencia de los sujetos que no recibieron alcohol. En otro estudio, a 4 diferentes grupos de hombres se les administraron 4 dosis de alcohol (0.08, 0.4, 0.8 y 1.2 g/kg) y fueron expuestos a estimulación visual erótica, observándose un efecto dosis-dependiente sobre las erecciones peneanas (medidas con pletismógrafo peneano). A mayor dosis de alcohol, fue mayor el efecto deletéreo sobre el potencial eréctil (Briddell & Wilson, 1976); es decir, a dosis bajas, los participantes mostraron un mayor potencial eréctil comparado con los participantes tratados con dosis altas. Este resultado es similar a lo encontrado en el presente trabajo, en el que ratas tratadas con dosis bajas presentaron un efecto facilitador sobre el potencial eréctil de la rata macho mientras que a manera dosis-dependiente, las dosis altas deterioraron dicho potencial.
En la presente investigación, el uso de la rata como modelo animal permitió, en primer lugar, el estudio independiente de la motivación y activación sexual (tradicionalmente usados como equivalentes), lo cual representa un enfoque útil y novedoso en el contexto de la conducta sexual. En segundo lugar, permitió un mejor control de variables de la administración del alcohol, ya que en el humano existen variables imposibles de controlar debido a cuestiones éticas, costos, viabilidad de proyectos, entre otros. Así, los resultados de este trabajo muestran que la motivación y activación sexual son dos procesos diferentes, semi-independientes, cuya sensibilidad al alcohol es diferente: dosis altas y moderadas de alcohol provocaron un efecto deletéreo sobre ambos procesos, mientras que dosis bajas facilitaron la activación sexual.
A pesar de las numerosas bondades de utilizar este modelo animal, es importante mencionar algunas limitaciones. En efecto, si bien son ampliamente conocidas las bases neurales y hormonales de la conducta sexual de la rata macho y aun cuando los circuitos neurales en la rata y el hombre son muy similares, hacer la extrapolación al hombre es bastante difícil, considerando aspectos como el diferente metabolismo del alcohol y la importante influencia social y cultural sobre la conducta sexual humana.

