











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkLas vesículas son sistemas coloidales esféricos constituidos generalmente por una bicapa lipídica, los cuales pueden contener sustancias polares en su interior y, a la vez, estar dispersos en medios acuosos. La forma, el tamaño y la distribución de las vesículas varían dependiendo de las características físicas de los fosfolípidos, así como del método de preparación que se utilice. Estos sistemas coloidales son de particular interés debido a sus características estructurales, las cuales se pueden clasificar como unilamelares o multilamelares (Evans y Wennerströn, 1999). En este contexto, las vesículas pueden ser aprovechadas en aplicaciones como liberación de drogas (van der Meel et al., 2014), transporte de genes (Balazs y Godbey, 2010), diseño de sensores (Damhorst et al., 2013), formulaciones de vacunas (Schwendener, Ludewig, Cerny y Engler, 2010) y aplicaciones en el control microbiano (Yang et al., 2009).
Entre los compuestos más comunes utilizados en la preparación de liposomas se encuentran los fosfolípidos tales como: la fosfatidilserina, la fosfatidiletanolamina, el dipalmitoilfosfatidilglicerol, el diacetilfosfato, la lecitina, entre otros. La fosfatidilcolina es el componente principal de la lecitina, y consiste en una molécula anfífílica que posee residuos de fosfato y colina, unida por un residuo de glicerol a una cadena hidrofóbica, formada por dos residuos de ácidos palmítico y oleico. Esta estructura le permite a la fosfatidilcolina poseer un carácter polar y no polar y una simetría que determina su autoensamblaje en solución acuosa (Lasic, 1998).
La formulación de vesículas liposomales es muy variada y puede incluir diferentes etapas como extrusión, preformado y sonicación (Pérez, 2010). Las ondas inducidas por la sonicación rompen las vesículas más grandes en estructuras más pequeñas, resultando una dispersión transparente con tamaños de partículas cercanos a los 100 nm. Sin embargo, las dispersiones vesiculares tienden a ser termodinámicamente inestables e inducen la agregación de las nano estructuras, debido a las interacciones de atracción de van der Waals (Sabín, Prieto, Ruso, Hidalgo-Alvarez y Sarmiento, 2006). Diferentes estrategias han sido propuestas con el fin de favorecer la estabilización de las vesículas liposomales en disolución. Koetz y colaboradores han estudiado mezclas de dodecilsulfato de sodio (SDS) y lecitina en agua. La adición del surfactante aniónico induce una reducción significativa de la viscosidad y una aparente variación de la microestructura multilamelar a una unilamelar o planar. Los autores han atribuido este efecto a las fuerzas de repulsión electrostáticas entre las vesículas negativamente cargadas. La adición de un polímero catiónico, como el cloruro de poli (dialildimetilamonio) (PDADMAC), sobre el sistema lecitina/SDS también fue estudiada. Se debe indicar que la adsorción del polímero catiónico sobre la bicapa negativamente cargada genera el restablecimiento de la microestructura multilamelar inicial (Hellweg, Brûlet, Lapp, Robertson y Koetz, 2002; Robertson, Hellweg, Tiersch y Koetz, 2004). La preparación de catonosomas, es decir, vesículas constituidas por fosfolípidos y surfactantes catiónicos ha sido reportada por varios estudios (Alves et al., 2007; Kim, Han, Jang y Seong, 2014; Koirala et al., 2016; Pascoe y Foley, 2003). Por ejemplo, Koirala y colaboradores han estudiado el sistema fosfatidil colina y colesterol en presencia de bromuro de haxadeciltrimetil amonio (CTAB) y bromuro de dihexadeciltrimetil amonio (DHDAB). En particular, la adición de DHDAB resultó en una disminución del tamaño hidrodinámico y una mayor estabilidad de las vesículas preparadas.
El uso de polímeros tanto sintéticos (Hristova, Kenworthy y McIntosh, 1995; Hristova y Needham, 1995; Kenworthy, Simon y McIntosh, 1995; Robertson et al., 2004; Sou, Endo, Takeoka y Tsuchida, 2000) como naturales (Madrigal-Carballo et al., 2010) ha sido también reportado en la bibliografía. La estabilización mediante la adsorción de sus cadenas iónicas o neutras sobre la superficie de las vesículas genera una barrera estérica o electro estérica que evita la colisión entre las partículas. Sou y colaboradores, han reportado el uso de poli(etilenglicol) (PEG) sobre vesículas de fosfatidiletanolamina. La incorporación del polímero y su interacción sobre la superficie fosfolipídica fue estudiada por medio de calorimetría de titulación isotérmica. Los resultados demostraron una reducción considerable de la disociación polímero-vesícula, la cual se explicó en términos de una mayor interacción entre los residuos hidrofóbicos de PEG-lípidos y como consecuencia, una menor agregación de las vesículas (Sou et al., 2000). El uso de biopolímeros ha sido propuesto por su reconocido interés en sus propiedades biocompatibles y biodegradables. El quitosano es un polímero obtenido de la desacetilación de quitina. Este polímero presenta en soluciones acuosas ligeramente ácidas, grupos aminos regularmente protonados. Esto podría conducir a una interacción de tipo electrostática con los fosfolípidos regularmente cargados negativamente. El aprovechamiento de esta interacción permite la obtención de liposomas recubiertos con una menor tendencia a la agregación por periodos de tiempo mayores (Mertins y Dimova, 2011).
El presente trabajo se enfocó en la preparación de vesículas liposomales compuestas por fosfatidilcolina y por mezclas de PC/fosfolípidos. El efecto de adicionar diferentes fosfogliceroles sobre el tamaño, la polidispersidad y el potencial zeta fue evaluado. Adicionalmente, la modificación de una mezcla constituida por fosfatidilcolina y dimiristoilfosfoglicerol sal de sodio (DMPG-Na) con quitosano fue estudiada en mayor detalle, por medio de calorimetría de titulación isotérmica. Adicionalmente, las vesículas se caracterizaron mediante técnicas que permiten determinar el efecto de la adsorción del polímero sobre la carga superficial, tamaño y estructura.
Metodología
Los fosfolípidos, L-α-fosfatidilcolina extraída de la soya (PC) (94%), el dimiristoilfosfoglicerol sal de sodio (DMPG-Na) (98%), diestearoilfosfoglicerol sal de sodio (DSPG-Na) (98%) y dipalmitoilfosfoglicerol sal de sodio (DPPG-Na) (98%) fueron donados por la empresa LIPOID (Alemania). El quitosano purificado extraído de cáscaras de camarón fue proporcionado por el Laboratorio de Polímeros de la Universidad Nacional. La masa molecular viscosimétrica del quitosano utilizado fue de 865 kDa y se determinó siguiendo el procedimiento descrito en la norma ASTM D 2857 (ASTM D2857-16). El grado de desacetilación del quitosano usado fue de (77,8 ± 0,6)% y se llevó a cabo basado en el método reportado por Czechowska-Biskup, B. (Czechowska-Biskup, Jarosińska, Rokita, Ulański, y Rosiak, 2012) Todas las disoluciones fueron preparadas utilizando agua desionizada.
El proceso de preparación de las vesículas consistió en dispersar 0,02 g de PC y 0,02 g de DMPG-Na en 20 mL de agua desionizada. La dispersión de agitó a 150 rpm durante 72 horas a temperatura ambiente. Transcurrido este tiempo, la dispersión se transvasó a una botella de 250 mL y se le añadió 20 mL más de agua desionizada. Esta se agitó durante 72 horas más, para lograr una dispersión con concentración final de 1 mg/mL de fosfolípidos. Este proceso se repitió para obtener vesículas de igual concentración para PC y las mezclas PC/DPPG-Na y PC/DSPG-Na. Al finalizar el proceso de hinchamiento las dispersiones se almacenaron a 4°C.
Los sistemas vesiculares a base de fosfolípidos se recubrieron con quitosano, siguiendo el procedimiento descrito por Quemeneur, Rinaudo, Maret, y Pépin-Donat ( 2010). Se tomaron 2 mL de las dispersiones de PC/DMPG-Na 1 mg/mL previamente sonicada y se adicionaron alícuotas de 50 μL de una disolución de quitosano de 1 mg/mL. Después de cada adición se agitó por 5 minutos y se determinó el tamaño y la carga superficial de las vesículas.
Por medio de la técnica de dispersión de luz (DLS, siglas en inglés) se determinó el tamaño de partículas y la distribución de tamaño de las dispersiones vesiculares preparadas. Estas se realizaron en un Zetasizer (Malven Instruments, Nano-ZS90) con un ángulo de 90°, equipado con un láser He-Ne (λ = 633 nm, 4 mW) a una temperatura controlada de 25 ºC. Cada determinación se realizó diez veces y con un mínimo de diez corridas, cada una con una duración de 10 s. Adicionalmente, la carga superficial de las partículas en dispersión se determinó mediante la medición del potencial Z. El análisis se llevó a cabo utilizando el mismo instrumento. Cada determinación se realizó diez veces y con un mínimo de diez corridas por cada determinación. La caracterización anterior fue complementada mediante la técnica de microscopía electrónica de barrido criogénico (Cryo-SEM, en sus siglas en inglés). Las muestras fueron congeladas utilizando nitrógeno líquido y fracturadas a -180 °C, el grabado se realizó durante 60 s a -98 °C, en la cámara de preparación cryo GATAN Alto 2500. Por último, la muestra se transfirió al Cryo-SEM S-4800 Hitachi. La energía involucrada, producto de las interacciones entre el quitosano y las vesículas de fosfolípidos, se determinó mediante la técnica de calorimetría de titulación isotérmica (ITC), mediante el procedimiento descrito por Dimova (Mertins & Dimova, 2011). Las vesículas utilizadas fueron dispersadas en un tampón de ácido acético y acetato de sodio con una concentración total 80 mM a pH 4,5. El instrumento empleado fue un NanoITC (TA instruments). La celda de trabajo con capacidad de 1 mL se llenó con la dispersión fosfolipídica y la celda de referencia con agua. La primera inyección fue de 2 uL seguida por 24 inyecciones de 10 uL de disolución de quitosano, el cual se encontraba disuelto en el mismo tampón que la dispersión liposomal a un pH de 4,5. Cada inyección fue realizada en intervalos de 200 s, con una velocidad de agitación de 300 rpm a una temperatura de 25 °C. Seguidamente, se realizaron los experimentos de referencia correspondientes, la titulación del quitosano sobre la dispersión de vesículas a base de PC 1 mg/mL y la titulación de la solución de quitosano sobre la disolución tampón. Para evitar la presencia de burbujas, todas las muestras se desgasificaron con un baño ultrasónico durante 20 min antes de realizar las mediciones. Los análisis de datos se llevaron a cabo con el software de Launch ITCRun proporcionado por TA Instruments. El rango de concentraciones de quitosano que se trabajó fue entre 8,6x10-3 y 0,2 mg/mL. Las disoluciones de quitosano fueron previamente filtradas usando una membrana de celulosa de 300 micras de tamaño de poro.
Resultados y discusión
Con el propósito de evaluar el efecto de adicionar fosfolípidos cargados con diferentes longitudes de cadena sobre el fenómeno de autoensamblaje, se utilizó PC y mezclas de este con diferentes fosfogliceroles, tales como: DMPG-Na, DPPG-Na y DSPG-Na. Las dispersiones fueron preparadas y caracterizadas siguiendo el procedimiento descrito anteriormente. En la tabla 1 se observan los resultados obtenidos.
Tabla 1 Potencial Z y tamaño de partícula en función de la composición de las vesiculas.

Notas: Fuente propia del estudio. a Tamaño de partícula determinado por DLS usando el valor promedio (100- 99 %) obtenido por el análisis en porcentaje de número de partículas. Desviación estándar (σ).
Inicialmente se puede notar que las vesículas sin sonicar presentan tamaños en el orden de micras. Después del proceso de sonicación el tamaño disminuyó significativamente. Estos resultados pueden explicarse tomando en cuenta que el proceso de sonicación suministra energía al sistema y hace que las vesículas logren reordenarse en estructuras de menor tamaño y con una mayor uniformidad. Este efecto puede notarse en las vesículas de PC, donde el tamaño cambia significativamente desde 2608 nm hasta 278 nm. Por otro lado, las vesículas de PC experimentan un cambio significativo en el potencial Zeta (ξ), variando desde -43 mV hasta valores de -4 mV sin ningún efecto de agregación aparente, después del proceso de sonicación. El resultado obtenido puede interpretarse de manera similar a lo expuesto anteriormente, donde la sonicación induce a un mayor ordenamiento de las cadenas alifáticas y los residuos polares, cuyo resultado se evidencia en la reducción del tamaño y la polidispersidad de las estructuras vesiculares.
La adición de fosfogliceroles de diferentes longitudes de cadena resultó en una disminución del tamaño de las vesículas y una reducción del potencial Zeta a valores más negativos. Este resultado es de esperarse, considerando que la adición de fosfogliceroles favorece un mayor empaquetamiento por la incorporación de una doble capa fosfolipídica, la cual induce a un aumento de las fuerzas de van der Waals. Por otro lado, la adición de los fosfogliceroles aniónicos resultó en una disminución significativa de ξ a valores de -70 mV, lo que favoreció la repulsión electrostática y aumentó la estabilidad de la estructura a través del tiempo. Los resultados obtenidos son comparables con los reportados por otros estudios (Barenholz y Lasic, 1996; Uchegbu, Schätzlein, Cheng, y Lalatsa, 2013).
Una de las principales limitaciones de las vesículas liposomales es la tendencia al fenómeno de agregación con el tiempo. Este proceso se hace más pronunciado a valores de pH ácidos, donde se disminuyen las cargas superficiales. Una alternativa para evitar la degradación temprana de liposomas implica el recubrimiento de la superficie con una membrana polimérica (Madrigal-Carballo et al., 2010).
Estudios recientes han demostrado la efectividad del quitosano como agente estabilizante de los sistemas liposomales, al aumentar la estabilidad física y la eficiencia de encapsulamiento de estas (Prabhu et al., 2012). Para aumentar la estabilidad de las vesículas de L-α-fosfatidilcolina, se adicionó una disolución diluida de quitosano. La capacidad de recubrimiento del quitosano sobre las vesículas se monitoreó mediante la determinación de la carga eléctrica superficial y el tamaño de partícula. La figura 1 muestra la variación del potencial Zeta y del tamaño en función de la adición de una disolución diluida de quitosano al 0,86 mg/mL. El tamaño de partícula fue determinado por DLS utilizando el análisis en porcentaje de la intensidad.
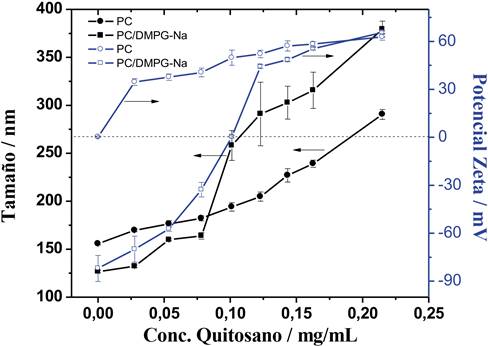
Nota: Fuente propia del estudio
Figura 1 Dependencia de ζ y el tamaño de partícula de las vesículas liposomales (1,38 mM) modificadas con una disolución de quitosano 0,86 mg/mL.
Como se observa en la figura 1, la adición de una concentración de quitosano menor a 0,025 mg/mL induce un aumento en la carga eléctrica superficial de las vesículas, hasta alcanzar un valor cercano a + 30 mV. Considerando que el polímero está disuelto en una disolución ácido acético (1% v/v), la adición de 0,025 mg/mL de quitosano sobre la dispersión fosfolípídica a base de PC induce un cambio en el pH de la dispersión de 7 a 4,5. Esta variación provoca que la superficie fosfolipídica adquiera una carga positiva debido a la protonación del grupo colina del PC y como resultado una interacción de tipo repulsiva con las cadenas poliméricas de quitosano. Lo anterior sugiere que las cargas positivas del polímero se neutralizan a muy bajas concentraciones con las cargas negativas disponibles de las vesículas, provocando que el recubrimiento sea parcial. Consecuentemente, el tamaño de las microestructuras incrementa ligeramente con la adición de quitosano hasta alcanzar un valor máximo de partícula de 275 nm a una concentración de 0,2 mg/mL de quitosano, sin registrarse una agregación aparente. Por el contrario, la interacción del quitosano con mezclas de PC/DMPG-Na experimenta un efecto más significativo en la medición del potencial Zeta y en el tamaño. Inicialmente, los resultados indican que las vesículas presentan un ζ de -72 ± 14 mV y un tamaño de partícula alrededor de los 125 nm a un pH cercano a 7. La adición de 0,1 mg/mL de polímero provoca el aumento del ζ hasta valores cercanos a 0 mV y un aumento de la microestructura a valores de 250 nm. Sin embargo, en las condiciones experimentales, la dispersión no presentó una sedimentación inmediata. El aumento en el potencial Zeta puede interpretarse como la incorporación de las cadenas de polielectrolito catiónico sobre la superficie negativa de las vesículas, debido a las fuerzas electrostáticas existentes. No obstante, la adición de mayor cantidad de polímero no induce un cambio aparente en la carga negativa del grupo fosfato del DMPG-Na. Esto implica la disponibilidad de una superficie permanente, negativamente cargada para la interacción con el polielectrolíto catiónico. La adición de concentraciones superiores a 0,1 mg /mL de quitosano resulta en estructuras de mayor tamaño, lo cual induce a la sedimentación de las vesículas. Al adicionar más polímero, el pH disminuye hasta valores de 4,5, lo que provoca que los grupos aminos estén totalmente protonados y sean adsorbidos con mayor intensidad por las vesículas vecinas induciendo la agregación del sistema. Resultados similares han sido reportados por Quemeneur et al., al utilizar liposomas unilamelares a base de 1,2-dioleoil-sn-glicero-3-fosfatidilcolina (DOPC), modificados con un quitosano de un peso molecular de 225 kDa y un grado de desacetilación de 95%. Quemeneur et al. sugieren que la incorporación del quitosano sobre la membrana de las vesículas se debe a una interacción eficiente de origen electrostático, producida entre los grupos amino cargados positivamente del polelectrolito y los fosfatos cargados negativamente en la superficie del liposoma (Quemeneur, Rammal, Rinuaudo, y Pépin-Donat, 2007).
Con el objetivo de visualizar las estructuras formadas a partir de las diferentes formulaciones de vesículas y la adición de quitosano, se realizaron experimentos de microscopía electrónica de barrido criogénico (Cryo-SEM). La figura 2 muestra las micrografías Cryo-SEM de las vesículas a base de L-α-fosfatidilcolina (PC), L-α-fosfatidilcolina y dimiristoilfosfoglicerol sal de sodio (PC/DMPG-Na) y L-α-fosfatidilcolina- dimiristoilfosfoglicerol sal de sodio modificada con quitosano (PC/DMPG-Na/Quitosano), después del proceso de sonicación.
En la figura 2A, se pueden observar estructuras esféricas con tamaños entre 50 y 200 nm para el sistema a base de PC. La adición de DMPG-Na sobre PC se muestra en la figura 2B. Esta imagen sugiere que la incorporación de un fosfoglicerol induce una mayor uniformidad en las dimensiones de las vesículas encontradas y una relativa reducción del tamaño. Los resultados obtenidos refuerzan valores determinados mediante dispersión de luz dinámica.
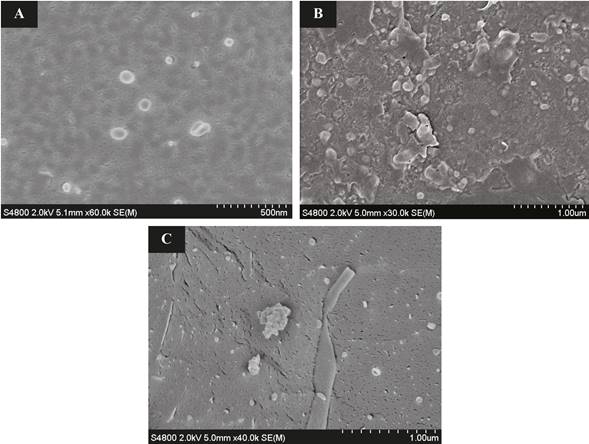
Figura 2 Micrografía Cryo-SEM de vesículas A) a base de PC, B) a PC/DMPG-Na y C) PC/DMPG-Na/Quitosano. Nota: Fuente propia del estudio.
El efecto de la incorporación de quitosano sobre las vesículas a base de PC/DMPG-Na puede visualizarse en la figura 2C. Se puede observar que la adición del polímero induce la agregación de las vesículas (área dentro del círculo), debido a la fuerte interacción electrostática que se ejerce sobre la superficie de las vesículas.
La calorimetría de titulación isotérmica se utilizó para caracterizar la interacción del quitosano con el medio amortiguador y con liposomas ligeramente cargados a base de PC y liposomas cargados a base de mezclas de PC y DMPG-Na. El resultado obtenido de la titulación de la solución de quitosano de concentración 0,86 mg/mL sobre la disolución tampón y sobre la dispersión de PC 1 mg/mL se puede observar en la figura 3.
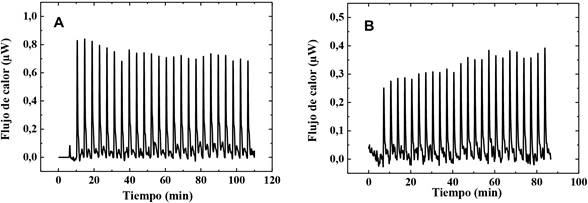
Nota: Fuente propia del estudio
Figura 3 A) Termograma obtenido de la titulación de quitosano 0,86 mg/mL sobre un tampón de HAc/NaAc 80 mM (pH 4,5) y B) sobre liposomas compuestos por PC 1 mg/mL (1,38 mM) a un pH 4,5.
Como se observa en esta figura, cada inyección de la solución de quitosano sobre la solución tampón produce una señal endotérmica, que se podría atribuir al proceso de dilución. El calor global asociado con las inyecciones corresponde a un promedio de 0,74 μW por inyección. En contraste, Mertins y Dimova reportaron flujos de calor diez veces menores, entre 0,07-0,08 μW, que podrían explicarse por el elevado peso molecular del polímero empleado en el presente trabajo. Esto sugiere que el proceso de reorganización de las cadenas de polímero en la disolución diluida del tampón involucra un alto consumo de energía (Mertins y Dimova, 2011, ).
La adición del quitosano sobre los liposomas a base de PC 1 mg/mL ambos disueltos en un tampón de pH 4,5, se representa en la figura 3B. Al pH utilizado, es de esperar que la interacción entre el quitosano y PC genere señales exotérmicas al principio de la adición. Sorpresivamente, los resultados reportados muestran un comportamiento opuesto, es decir, señales endotérmicas, correspondiente a un promedio de 0,33 μW por inyección. Este comportamiento demuestra que el calor de dilución del quitosano solapa el calor de la interacción entre el quitosano y PC debido a las bajas concentraciones de PC estudiadas y a la carga positiva adquirida debido a la protonación de los residuos de colina, que inducen una baja intensidad en la interacción con el polímero.
La caracterización de la interacción del quitosano con membranas cargadas negativamente se realizó utilizando mezclas de PC/DMPG-Na. El termograma obtenido en la figura 4A, muestra que la adición de quitosano sobre la suspensión de liposomas de una concentración de 1,38 mM genera señales altamente exotérmicas en comparación con el resultado obtenido en la titulación de vesículas de PC. Nótese que en la tercera inyección se alcanza la máxima liberación de calor, disminuyendo su intensidad conforme se adiciona más polímero, induciendo a que la carga negativa neta de la membrana se vuelva cada vez menos negativa. El aumento de la concentración de liposomas a 3,82 mM de fosfolípidos resulta en un aumento de las señales exotérmicas derivadas de la fuerte interacción electrostática polímero-vesícula.
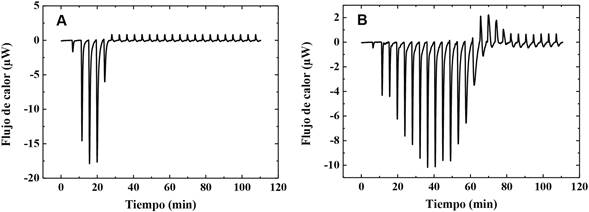
Nota: Fuente propia del estudio
Figura 4 Termogramas asociados a la titulación de A) la suspensión de liposomas 1,38 mM y B) 3,82 mM (52% de moles de DMPG-Na y 48 % PC), con quitosano 0,86 mg/mL.
Como se puede observar, la magnitud de las primeras cuatro señales de la dispersión más diluida es, aproximadamente, el doble que las señales producidas por la dispersión concentrada. Sin embargo, la dispersión más concentrada presenta un mayor consumo del polímero debido al incremento en el número de vesículas. Después de alcanzar la máxima liberación de calor las magnitudes de las señales, disminuyen conforme se adiciona más quitosano hasta alcanzar el calor cercano de dilución del polisacárido.
De acuerdo con los resultados registrados para la dispersión más diluida, se puede observar que después de cinco inyecciones de quitosano sobre las vesículas se alcanza el calor de dilución del polímero; al comparar estos con los resultados de potencial Zeta, es posible notar que a la misma concentración de polímero las vesículas aún presentan un potencial negativo de -28 mV. Estudios similares reportan que no es necesario alcanzar una inversión de la carga inicial del liposoma para asegurar un completo recubrimiento de la vesícula (Mertins & Dimova, 2011). La explicación de este fenómeno sugiere que en las primeras adiciones del polímero, las cadenas del quitosano son absorbidas en la membrana. Al aumentar la concentración del polímero en la dispersión, después de cada inyección, las cadenas del quitosano tenderán a difundir a los espacios libres, que no son lo suficientemente grandes para promover la adsorción de la cadena completa, debido al impedimento estérico. Aparte de este fenómeno, las cadenas recién inyectadas y adsorbidas evitarán la superposición de otras cadenas debido a la repulsión electrostática y a su rigidez. Esto podría conducir a una pobre organización de la macromolécula sobre la vesícula y una rápida saturación de la superficie (Mertins & Dimova, 2011). La adición de mayor concentración de polímero sobre la dispersión vesicular induce finalmente a la agregación de las partículas vesiculares.
Conclusiones
En este trabajo se ha estudiado la influencia de mezclar un fosfolípido zwitteriónico como la fosfatidilcolina (PC) con diferentes fosfogliceroles cargados negativamente; sin embargo, las vesículas de PC/DMPG-Na se estudiaron con mayor detalle. La adición de fosfogliceroles a las dispersiones de PC indujeron una mayor uniformidad en las dimensiones de las vesículas y un potencial Zeta más negativo, favoreciendo la estabilización de la nanopartícula. Usando microscopía electrónica de barrido criogénico (Cryo-SEM, por sus siglas en inglés), se confirmó la presencia de vesículas con una distribución de tamaños similares a los determinados por DLS. Además, la adición de quitosano sobre vesículas ligeramente neutras y negativamente cargadas fue evaluada. Mediante experimentos de calorimetría de titulación isotérmica se demostró una fuerte interacción electrostática del polielectrolito catiónico sobre las superficies cargadas de las vesículas, resultando mucho más intensa con las vesículas de PC/DMPG-Na. Se determinó que no es necesaria la reversión completa de la carga superficial de la vesícula para asegurar la saturación de la superficie del sistema liposomal con el quitosano. Los resultados obtenidos por medio de ITC fueron complementados con experimentos de potencial Zeta. La adición de quitosano a bajas concentraciones resultó en la modificación superficial de las vesículas; sin embargo, el incremento en la concentración del polímero indujo a la saturación y la consecuente agregación de las vesículas, debido al alto peso molecular del polielectrolito. Esto último fue demostrado por medio de Cryo-SEM.
Agradecimientos
Los autores agradecen el financiamiento del MICIT-CONICIT al proyecto “Materiales inteligentes 023930”. A la vez, se agradece a la empresa Lipoid AG de Alemania por la donación de los fosfolípidos y la colaboración del Prof. Joachim Koetz de la Universidad de Potsdam, Alemania, por los análisis de Cryo-SEM.

