











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
El clima a escala mundial ha cambiado desde siempre de forma natural (1); sin embargo, el deterioro de los reservorios naturales de carbono, principalmente por el uso de combustibles fósiles y la deforestación, han ocasionado el incremento de la concentración de los Gases de Efecto Invernadero (GEI) en la atmósfera (2). Producto de lo anterior han aumentado los efectos del cambio climático, fenómeno que está alterando el comportamiento de los distintos ecosistemas a nivel global (3), lo que para muchos científicos es el principal problema que enfrenta la sociedad actual (4)- (5). Se estima que de no existir un cambio positivo en las actividades humanas que provocan el aumento de uno de los GEI, la temperatura podría incrementarse en promedio entre 1,8 °C y 4 °C para el 2100 (6).
La respuesta de los países para enfrentar este fenómeno ha sido diversa; no obstante, se reconoce a nivel mundial, los esfuerzos de Costa Rica en el afán de colaborar en la búsqueda de soluciones, entre ellas, la política de carbono neutralidad fundamentada en la reducción de emisiones y en la compensación por el aumento en los reservorios a través actividades forestales (1). Al respecto, la forestación y reforestación son reconocidas por el Protocolo de Kioto, como una de las medidas más efectivas para aminorar el cambio climático (7).
Se estima que las plantaciones en el mundo tienen un potencial de secuestro de carbono equivalente al 52 % de las emisiones de carbono proyectadas para el 2100 en los sectores energía e industria (8). A pesar de ello, James, Phua, Awang y Mokhtar (9) apuntan diferencias en la acumulación de biomasa, carbono, en los diferentes arreglos para la producción forestal; e inclusive a nivel de distintas condiciones edáficas (10). Se debe realizar investigación que abarque desde los sistemas más complejos hasta los más simples en términos de biodiversidad, como son las plantaciones puras de especies como teca y melina, objeto del presente estudio.
Tectona grandis (teca) y Gmelina arborea (melina), son especies exóticas en Costa Rica, de rápido crecimiento y las de mayor superficie plantada en Costa Rica, ambas suman 59 215 ha (11. La madera de teca es de excelentes características y se utiliza en la fabricación de productos de alto valor en el mercado mundial (12). Por su parte, la melina registra una madera de menor precio y tiene muchos usos. En Costa Rica se utiliza para construcción en general y, en un alto porcentaje, para la fabricación de tarimas para el embalaje, principalmente de productos agrícolas (13)- (14). Ambas especies, por el área plantada y por su velocidad de crecimiento, pueden aportar de manera significativa al objetivo nacional de carbono neutralidad. Por lo antes expuesto y la importancia que tiene el cálculo de secuestro de carbono se desarrolló la investigación con el objetivo de desarrollar modelos matemáticos que faciliten calcular la producción de biomasa por hectárea para teca y melina, mediante los cuales se genere información que permita establecer indicadores nacionales sobre el potencial que tienen ambas especies para mitigar el cambio climático.
Materiales y métodos
Área de estudio. El área de estudio comprendió la zona norte, pacífico norte y sur de Costa Rica, en sitios con plantaciones de teca y melina establecidas con material clonal. En el Cuadro 1 y la Figura 1 se muestra la localización de los sitios de muestreo. En la zona norte se trabajó en los cantones Guatuso, San Carlos y Upala, que poseen un clima lluvioso todo el año, la precipitación media anual es de 3 056 mm y la temperatura media de 24 °C. En el pacífico norte se trabajó en los cantones Nicoya, Nandayure, Santa Cruz y La Cruz (Guanacaste), además de Lepanto y Puntarenas (Puntarenas), la precipitación promedio es de 1 888 mm al año y la temperatura media de 27 °C. En la zona sur, en los distritos Puerto Jiménez (Golfito) y Piedras Blancas (Osa), los que registran una precipitación promedio de 3 650 mm al año y la temperatura media 26 °C (15).
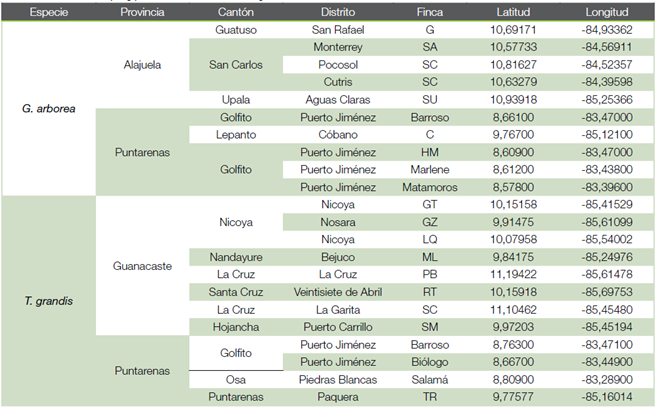
Figure 1. Location of the sampling points for melina and teak.
Muestreo. En las plantaciones evaluadas, de ambas especies, se establecieron parcelas permanentes de muestreo de 500 m2 que fueron remedidas todos los años, el número de mediciones en cada parcela varió según la edad de la plantación, en teca de 1 a 17 años, en melina de 1 a 9 años. También se utilizó las series de mediciones de ensayos clonales de melina desde 1 hasta 5 años.
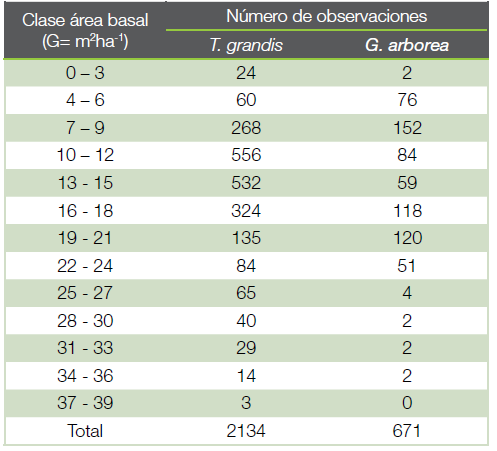
En cada unidad de muestreo se registraron diferentes variables dasométricas como diámetro a la altura de pecho (Dap) medido con cinta diamétrica y altura total medida con clinómetro suunto ; además de la calidad mediante la forma de las trozas, Para efectos de este trabajo solo se consideró el Dap. Con la información de cada unidad de muestreo se confeccionó una base de datos, y una vez revisada para identificar inconsistencias y eliminar los datos que no pertenecen a esa población, se calculó el área basal (m2) por árbol y por hectárea. El tamaño de la muestra en teca fue de 2 134 observaciones y de 671 para melina (Cuadro 2), aseguró la representatividad de las distintas calidades de sitio para el cultivo de ambas especies en Costa Rica. La biomasa por hectárea se calculó al aplicar, a cada individuo de la unidad de muestreo, los modelos alométricos desarrollados por (16) y (17), y luego extrapolada a hectárea.
Ajuste y selección de modelos. Se realizó análisis de regresión simple mediante el método de mínimos cuadrados ordinarios con el programa estadístico Statgrapihcs Centurion XVI. Para el conjunto de datos se ensayaron aproximadamente 10 modelos para predecir la biomasa por hectárea (Mgha-1) del componente hojas (B. hojas), ramas (B. ramas), raíz (B. raíz), fuste (B. fuste), leñoso (ramas+fuste+raíz) (B. leñoso) y árbol completo (B. total). En todos los casos se utilizó como variable independiente el área basal por hectárea (G en m2ha-1). La comprobación de los supuestos del análisis de regresión (normalidad, independencia y homogeneidad de varianzas) se realizó por el método gráfico (18). Además, se probaron transformaciones de las variables para mejorar el ajuste y corregir la heterocedasticidad (18)- (19).
La selección de la ecuación de mejor ajuste se realizó tomando en cuenta: a) la variabilidad del modelo explicado por el coeficiente de determinación (R2) (20), b) la precisión de las estimaciones en función a la raíz del cuadrado medio del error (RCME) (20)- (21)- (22); y c) el comportamiento de la ecuación determinado por el error absoluto medio (EMA) (22).
Para la precisión y validación de los modelos se tomó en consideración: a) el método gráfico de dispersión entre los observados vs predichos (20), y b) el error absoluto promedio porcentual o sesgo: EAPP=1/n(Σ(P-O)/O)100)) (20)-(22)-(23)-(24), donde: P, la biomasa predicha o simulada; O, la biomasa observada y n, el total de datos.
Adicionalmente, se calculó el estadístico Durbin Watson (DW) que mide la independencia de los datos (25), el Índice de Furnival (IF) para comparar modelos lineales con aquellos donde la variable dependiente es transformada (26), el criterio de información de Akaike (AIC) como medida relativa de la calidad del modelo (23) y el error del modelo o sesgo (E%) (19)- (23)- (24), error cuadrático medio (ECM) y la diferencia agregada (DA).
Todos los estadígrafos calculados fueron ranqueados según su magnitud relativa tomando como referencia la metodología expuesta por Segura y Andrade (26), Salas (27) y Bueno et al. (19).
Resultados
Modelos de predicción. En los cuadros 3 y 4, además de las figuras 2 y 3 muestran las estimaciones de los parámetros y las estadísticas de bondad de ajuste para el modelo elegido, por componente de la biomasa y la biomasa total. Las estimaciones de los parámetros fueron significativas (α = 0,05). Todos los modelos presentaron un ajuste satisfactorio y similar (R2 entre 95,3 % y 95,8 %) en teca y de 85,3 % a 97,1 % en melina, con errores o sesgos de estimación inferior a 2,5 %, condición que se observa en la figura 2b, 3b al ubicarse la nube de puntos muy cercana a la línea de tendencia central cumpliéndose con el supuesto de normalidad.
Los estadígrafos de predicción (RCME y EMA) menor a 0,11 para teca e inferior a 0,23 en melina, cifras también son aceptables, significan que la biomasa total calculada tiene una diferencia con relación a observada de 0,11 Mgha-1 en teca y de 0,23 Mgha-1 en melina. Los otros parámetros estadísticos también presentan valores bajos (muy aceptables) en ambas especies (Cuadro 3 y 4).
Los resultados indicaron una relación positiva fuerte entre el área basal y la biomasa, que se confirma con el coeficiente de determinación (R2); para todos los modelos seleccionados, el área basal explicó más 85 % de la variabilidad observada en la biomasa.
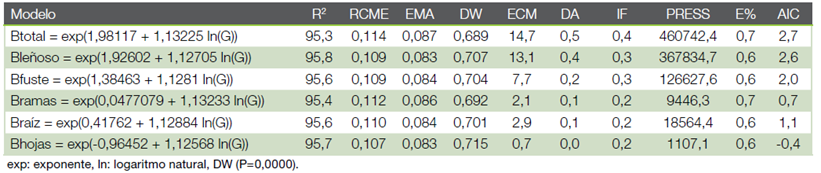
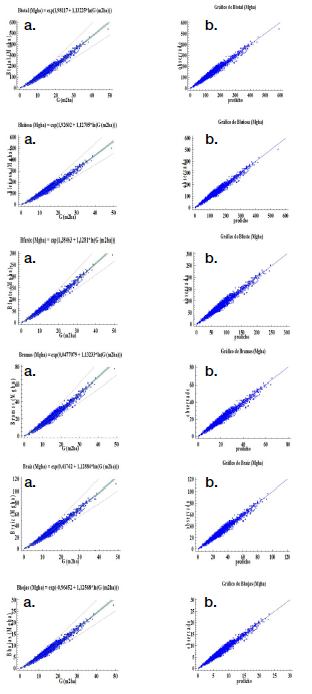
Figura 2 Modelos para estimar biomasa por componente y biomasa total (Mgha-1) en T. grandis: a) gráfico para el modelo ajustado, b) gráfico de valores predichos vs observados.
Figure 2. Models to estimate biomass by component and total biomass (Mgha-1) in T. grandis: a) graph for the adjusted model, b) graph of predicted vs observed values.
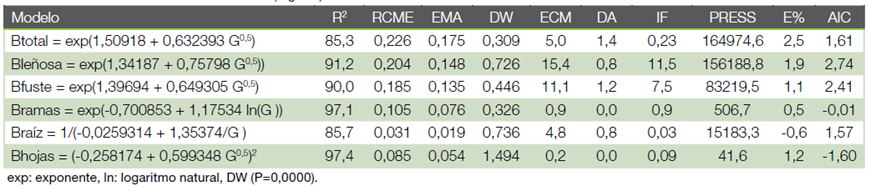
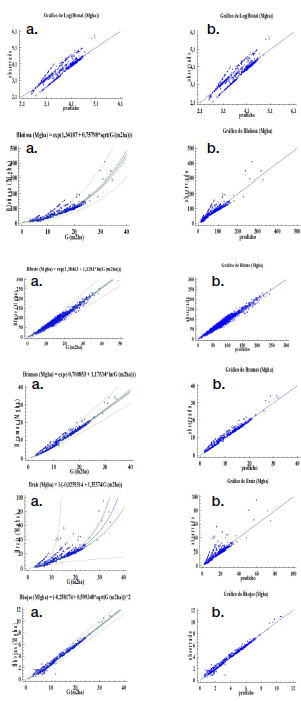
Figura 3. Modelos para estimar biomasa por componente y biomasa total (Mgha-1) en G. arborea: a) gráfico para el modelo ajustado, b) gráfico de valores predichos vs observados.
Figure 3. Models to estimate biomass by component and total biomass (Mgha-1) in G. arborea: a) graph for the adjusted model, b) graph of predicted vs observed values.
Discusión
Con el fin de evaluar la capacidad de los ecosistemas para mitigar el cambio climático, el esfuerzo en desarrollar modelos matemáticos (regresiones o ecuaciones) para predecir la biomasa del árbol o de sus componentes es abundante, y la literatura mundial sigue reportando con frecuencia nuevos estudios de esta naturaleza. También se ha demostrado que variables dasométricas del árbol como el diámetro y la altura, principalmente, son buenas predictoras de la biomasa o el carbono (28). No obstante, son escasos los modelos para estimar o predecir la biomasa o el carbono por unidad de área (por hectárea), y específicamente para teca y melina no existen. Ejemplo de ello es el estudio desarrollado por James et al. (9), quienes debieron emplear modelos desarrollados en distintos países y condiciones, con lo que se incrementa el nivel de incertidumbre y veracidad del resultado final.
Este tipo de modelos de regresión, también llamados modelos agregados, tienen la ventaja de que permiten hacer el cálculo directo por hectárea, sin necesidad de extrapolaciones, como en el caso de las ecuaciones a nivel de árbol. López, Estuardo y Alvarado (29) reportan que dicho procedimiento en algunos casos llevaría a cometer ciertos errores, sino se tiene experiencia y cuidado.
Los modelos elegidos son robustos, es decir, tienen buenos ajustes (R2 > 85%), con errores o sesgos de predicción inferior a 2,5% y parámetros de predicción muy bajos (RCME y EMA), menor a 0,11 para teca y a 0,23 en melina, valores que coinciden, en forma general con lo reportado en la literatura para otras especies. En Alnus acuaminta HBK se logró un ajuste de 99,3% para la biomasa arbórea y de 89,6% para la biomasa total, en ambos casos se reporta un RCME inferior a 0,23, ECM menor a 11,8 y el error de estimación menor 1,2% (30). Para Hieronyna alchorneoides y Vochysia guatemalensis, especies que son nativas pero se han establecido en plantaciones, Fonseca et al. (31) modelaron la biomasa arbórea y total en función del área basal con R2 mayor a 94% y un RCME entre 0,35 y 0,66. Cifras inferiores reportaron (32), las que representan menor robustez y precisión de los modelos, con R2 de 79,6 % a 81,5 %, RCME entre 10 y 23,0, EMA de 8,15 a 19,5 y E% entre 4 % y 10% para biomasa y carbono aéreo y total en bambúes.
Los modelos de predicción de la biomasa o el carbono en función del área basal en bosque natural, también muestran ajustes tan buenos como los logrados en el presente estudio. Para un grupo de especies de bosque secundario (33), registraron un R2 de 95,7 %; 96,0 % y 97,8 % con RCME menor a 0,31 para la biomasa total, carbono total y carbono en la biomasa arbórea, respectivamente. Para bosques naturales caducifolios, (34) lograron ajustes (R2>0,96%), con cifras de RCME 0,08 y 0,54, EMA entre 0,07 y 0,43 y sesgos menores a 0,6%.
Llama la atención los resultados obtenidos en el presente estudio por varias razones: en primer lugar, sorprende la robustez de los modelos para teca con ajustes (R2) tan similares para todos los componentes, y en segundo lugar, el excelente ajuste para el componente hojas y ramas en ambas especies, porque el reporte de la literatura indica que, por lo general, son más difíciles de modelarlos (34), (35), (36), (37) al registrar R2 más bajos y los demás estadísticos con cifras más altas que indican menor precisión. Los modelos elegidos para ambas especies (Cuadro 3 y Cuadro 4), complementados con la fracción de carbono determinada para la especie, a saber: en teca, 0,41; 0,45; 0,46 y 0,46 para el componente hojas, ramas, raíz y fuste, respectivamente, y para melina, 0,39; 0,46; 0,48 y 0,45 (38)- (39), permiten determinar la capacidad de mitigación del cambio climático del sistema productivo en un momento de su vida.
Conclusiones
Los modelos elegidos, además de su sencillez y practicidad por usar solo el área basal como variable independiente, presentan muy buena robustez y precisión, y representan un avance importante para la cuantificación confiable y el monitoreo de la biomasa o el carbono en una plantación forestal, lo cual los convierte en una herramienta muy útil siempre que se utilicen bajo condiciones similares.
Este tipo de estudios permite el establecimiento de indicadores nacionales de secuestro de carbono (al multiplicar la biomasa calculada por la fracción de carbono de la especie) y la no dependencia de modelos desarrollados en otras latitudes para evaluar la capacidad de mitigación del cambio climático de los ecosistemas forestales, esto es fundamental para Costa Rica que se declaró la carbono neutralidad con el máximo nivel de precisión, la Tier 3 según el IPCC.

