











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
Los bosques ribereños son esenciales en el mantenimiento de diversos procesos ecológicos y generación de servicios ecosistemicos (Chará, Giraldo, Chará-Serna, Pedraza y Camargo 2011; Fernández, Souza y Tanaka, 2014). A pesar de ello en la actualidad se encuentran altamente amenazados en el Gran Área Metropolitana de Costa Rica (GAM) y enfrentan situaciones como el aumento de la población, falta de planificación de las actividades urbanas, deforestación y pérdida de conectividad biológica (Leandro, Coto y Salgado 2010; Calvo y Mora 2012).
Por esta razón la Compañía Nacional de Fuerza y Luz (CNFL); una empresa dedicada a la venta y distribución de electricidad en Costa Rica, en conjunto con instituciones gubernamentales y municipalidades de la GAM, proponen la creación de corredores biológicos interurbanos como elemento base para la recuperación de la cuenca del Virilla, mediante el fortalecimiento de la conectividad biológica y los servicios ecosistémicos (Feoli, 2013). Uno de ellos es el Corredor Biológico Interurbano Río Tibás (CBRT).
Este corredor biológico pretende restaurar la cobertura vegetal en territorios ubicados 300 m a ambos lados del cauce principal del río Tibás, integrando áreas naturales, públicas, privadas, rurales y urbanas, para lo cual se requiere información de línea base. La presente investigación tiene como objetivo determinar la estructura y composición de la cobertura arbórea del sitio propuesto como CBRT para orientar futuras estrategias de restauración.
Materiales y métodos
Área de estudio
La ubicación del CBRT fue establecida por la CNFL y corresponde a un área de 300 m en ambos lados del río Tibás, entre las coordenadas 10°01,1091’latitud Norte y 83°03,358’O longitud Oeste. El río se extiende 19,5 km entre los 1000 y 2000 msnm y se ubica en la parte alta de la subcuenca del río Virilla. Administrativamente tiene jurisdicción en 3 cantones; San Rafael, San Isidro y Santo Domingo de Heredia (ITCR, 2014). Su temperatura media anual varía desde los 22,5 ºC en la parte más baja hasta 16 ºC en la zona más alta y la precipitación oscila aproximadamente entre 1600 mm anuales hasta 2500 mm anuales en la parte baja y alta respectivamente.
Para este estudio el área de interés se estratificó en tres zonas según las franjas altitudinales, definidos como zona alta (1530-2000 msnm), media (1190-1529 msnm) y baja (1000-1189 msnm) (figura 1). Dicha zonificación fue elaborada por la CNFL con base en las características del cauce.
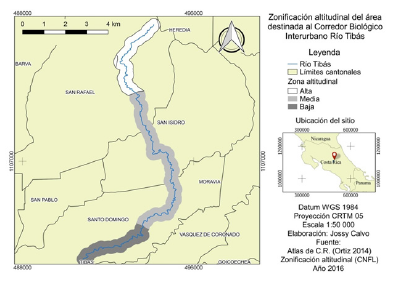
Muestreo
El muestreo se realizó entre enero y junio de 2016. Se delimitaron las zonas boscosas del CBRT a través de imágenes satelitales, trabajo de campo y el programa de sistemas de información geográfica QGIS (versión 2.12 Lyon, 2009). Posteriormente en el mismo software, para cada estrato del sitio se ubicaron y seleccionaron al azar parcelas de muestreo de 15 x 15 m cada una.
El número de parcelas en cada zona altitudinal se determinó por medio de curvas de acumulación de especies hasta alcanzar una curva asintótica, como indicador representativo del número de especies en cada estrato. Los datos fueron procesados con el programa “Estimates Win 820” y se utilizó el estimador de riqueza CHAO 1 Villarreal et al. (2006). En total se establecieron 61 parcelas de muestreo (15 x 15 m); 20 en la parte baja, 23 en la zona media y 18 en la parte alta del corredor.
Se determinó la riqueza, abundancia y diámetro a la altura de 1,3 m (DAP) de individuos con DAP mayor a 10 cm. Se evaluó la regeneración natural según la abundancia y riqueza de brinzales (individuos entre 30 cm y 1,5 m de altura) en sub-parcelas de 2 × 2 m ubicadas en las esquinas de cada parcela (para un total de 244). Asimismo, con un densiómetro se midió el porcentaje de cobertura de dosel en cada sub-parcela (Mostacedo y Fredericksen, 2000). Adicionalmente, se mencionaron y categorizaron especies de interés en conservación de acuerdo con su grado de endemismo o vulnerabilidad según la lista roja de la IUCN.
Análisis de datos
Se determinó el índice de valor de importancia (I.V.I) para cada especie y familia (Curtis y McIntosh, 1950). Se determinaron índices de diversidad de Shannon-Wiener y equidad de Pielou (Villarreal et al., 2006), mediante el programa PAST versión 3.8 (Hammer, 2001). Para estimar el grado de similitud en la composición arbórea entre estratos se realizó un análisis de ordenamiento por escalamiento multidimensional no métrico (NMDS), usando índices de disimilitud de Bray-Curtis, con el software R versión 3.3.1 (R Core Team, 2012). Posteriormente se aplicó una prueba ANOSIM para comprobar diferencias significativas entre las tendencias observadas (Clarke y Warwick, 2001).
Para evaluar la estructura forestal se realizaron gráficos de frecuencias con los datos de DAP, con el fin de separar categorías diamétricas y estimar el estado de sucesión de los ecosistemas. Asimismo, se estimó la densidad de árboles y brinzales (N) por hectárea para cada estrato (Mostacedo y Fredericksen, 2000). Los datos obtenidos de porcentajes de cobertura vegetal y densidad de individuos fueron analizados estadísticamente mediante la prueba no paramétrica de Kruskal-Wallis para valorar diferencias significativas entre los distintos estratos y relacionar su grado de perturbación. Finalmente, se evaluó el porcentaje de especies e individuos según el gremio ecológico que representan (heliófitas, esciófitas) y el síndrome de dispersión (anemocoria, antropocoria, autocoria y zoocoria) con el fin de inferir sobre su diversidad funcional.
Resultados
Composición arbórea: Se registraron en total 1651 individuos; 45 familias y 159 especies. De estos 650 individuos (43 familias y 113 especies) fueron árboles (DAP mayor a 10 cm) y 1001 brinzales (41 familias y 122 especies). No se logró la identificación de once especies de brinzales debido a su desarrollo temprano y falta de caracteres sexuales (tres Piperaceae, dos Asteraceae, una Solanaceae y cinco desconocidos). Estos no se incluyeron en los análisis a nivel de especie.
El esfuerzo de muestreo para estimar la riqueza se considera adecuado en la zona baja y media del CBRT. Ambas curvas de acumulación para árboles y brinzales empezaron a estabilizarse indicando baja probabilidad de encontrar nuevas especies. No obstante, en la zona alta se determinó que es muy probable encontrar nuevas especies., El estimador Chao 1 sugirió una riqueza de árboles (DAP > 10 cm) de 74 especies, de las solo 48 registradas (figura 2).
En esta última zona se dificultó la realización de más parcelas debido a que no se pudo contactar con muchos propietarios para adquirir el permiso de ingreso a fincas privadas. Asimismo, la región Norte, a partir de cerca de los 1800 msnm, corresponde a bosques muy poco intervenidos sin caminos de acceso, colindante con el Parque Nacional Braulio Carillo.
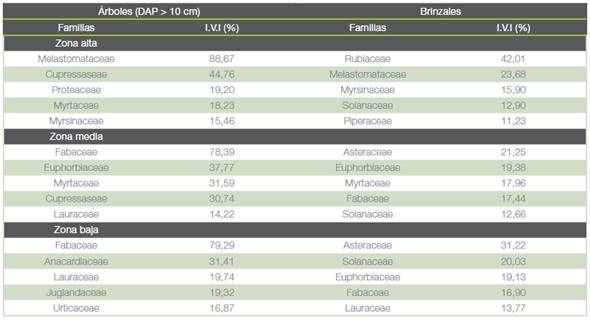
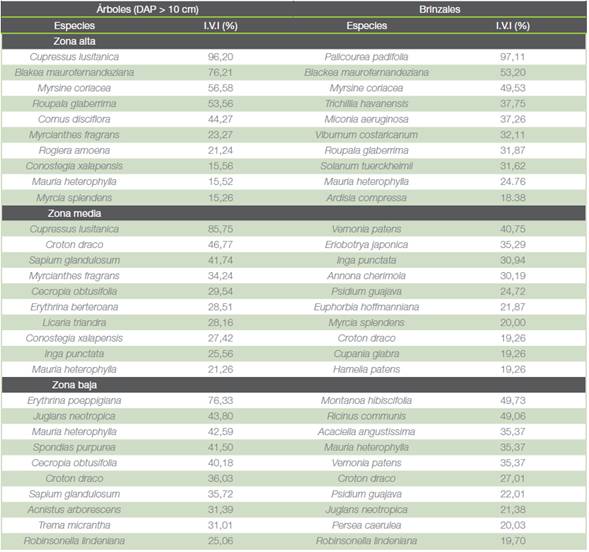
Las familias en árboles (DAP > 10 cm) de mayor importancia de acuerdo con el I.V.I fueron Melastomataceae (88,7 %) en la zona alta y Fabaceae en las zonas media y baja (78 % y 79 % respectivamente). En brinzales destacaron las familias Rubiaceae (42 %) en la zona alta y Asteraceae y Euphorbiaceae en los estratos más bajos (cuadro 1). Las especies con mayor peso ecológico en cada estrato según el índice se mencionan en el cuadro 2. Se destaca que las especies arbóreas con valores más altos son exóticas: Cupressus lusitanica Mill en las zonas alta y media y Erythrina poeppigiana Walp. O.F. Cook en la zona baja.
Se registraron 23 especies exóticas; 18 especies de árboles y 12 en regeneración. Las más abundantes fueron el ciprés que representó el 9 % de todos los árboles registrados, seguido del E. poeppigiana y el cedro nogal (Juglans neotropica Diels). En la zona baja la higuerilla (Ricinus communis L) representó el 8 % del total de brinzales registrados.
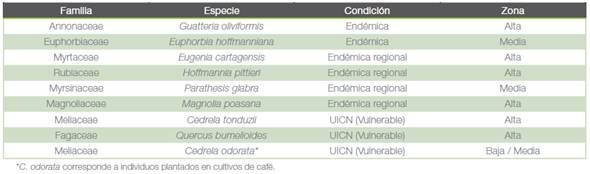
Nueve especies registradas son de interés especial en conservación. De estas, dos son endémicas: Guatteria oliviformis (Donn. Sm.) y Euphorbia hoffmanniana (Klotzsch y Garcke) Boiss), cuatro endémicas regionales (Costa Rica, Nicaragua y/o Panamá) y tres catalogadas como vulnerables según la lista roja de la UICN (cuadro 3).
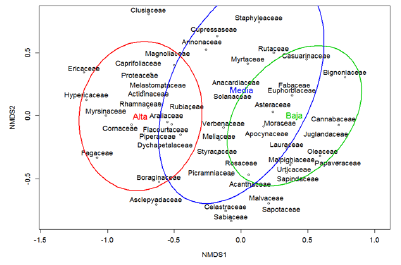
Figure 2 Spatial management of tree families according to the multidimensional non-metric staggering (NMDS) in altitudinal areas of the area destined for the Interurban Biological Corridor Tibás River, Heredia, Costa Rica (2d stress = 0.17, R = 0.48, P = 0.001). The insimilarity index of Bray Curtis was used.
Cuadro 1 Familias de árboles (DAP > 10 cm) y brinzales más importantes, según el Índice de Valor de Importancia, en los tres estratos altitudinales del área destinada al Corredor Biológico Interurbano Río Tibás, Heredia, Costa Rica.
Los resultados de riqueza, abundancia e índices de diversidad para cada estrato se presentan en el Cuadro 4. A pesar de la presencia de plantaciones de especies exóticas y perturbación por actividades antropogénicas (principalmente en las zonas media y baja) todos los estratos altitudinales presentaron índices de Shannon relativamente altos y baja dominancia de especies según el índice de Pielou.
Por otro lado, los tres estratos presentaron porcentajes de disimilitud de Bray Curtis por encima del 61 %. Estas diferencias también se explican a través de la composición de familias en distintos estratos (R=0,48, P=0,001). En la Figura 2 se presenta un modelo de ordenación bidimensional demostrando asociación de las familias vegetales con las distintas zonas altitudinales.
Estructura arbórea
La distribución de categorías diamétricas no difirió mucho entre las zonas altitudinales. Los tres estratos presentaron una distribución de “J” invertida (figura 3) y la proporción de árboles con DAP entre 10 y 39,9 cm osciló entre el 70 % en la zona media al 77 % en la zona alta, incluyendo dos clases diamétricas donde se ubicó la mayor cantidad de individuos.
La cobertura del dosel fue estadísticamente mayor en la zona alta (cuadro 5), la cual fue moderada en las zonas baja y media (Kruskal-Wallis, p < 0,05). La densidad estimada de árboles (DAP > 10 cm) para todo el sitio fue de 474 individuos/ha y no varió significativamente entre los estratos, sin embargo la densidad de brinzales sí presentó diferencias y fue mucho mayor en la zona alta (cuadro 5) (Kruskal-Wallis, p < 0,05).
Diversidad funcional
Las proporciones encontradas de acuerdo con la agrupación de gremios ecológicos y síndromes de dispersión se comportaron de manera similar según los análisis por número de especies o de individuos. Se observó mayor proporción de especies esciófitas (parciales y totales) y de zoocoria en la zona alta, mientras que en los estratos más bajos la proporción de los grupos de heliófitas efímeras y de anemocoria fue mayor (figuras 4 y 5 respectivamente). Cabe resaltar que entre el 2 y 10 % de especies no se clasificaron por la falta de información documentada en literatura (Sandor, 2012).
Cuadro 2 Porcentajes del Índice de Valor de Importancia para las 10 especies de árboles y brinzales con mayor valor en los tres estratos altitudinales del área destinada al Corredor Biológico Interurbano Río Tibás, Heredia, Costa Rica.
Cuadro 3 Especies de interés especial en conservación de acuerdo a su distribución geográfica limitada o estado poblacional según la lista roja de la UICN, registradas en distintos estratos altitudinales del área del Corredor Biológico Río Tibás, Heredia, Costa Rica.

Discusión
Composición arbórea
La consolidación de la mayor parte de la población costarricense en el Valle Central y el consecuente desarrollo de actividades agropecuarias y urbanas ha llevado a la pérdida de cerca del 80 % de bosques húmedos premontanos originales, así como la fragmentación del porcentaje restante (Sánchez- Azofeifa, Harris y Skole, 2001). En la región del CBRT la mayor parte del área está ocupada por cobertura boscosa (27 %), no obstante el 58 % de la misma se concentra en la zona alta, dominando en menores altitudes usos agrícolas (principalmente café) y urbano.
Las especies abundantes en las zonas media y baja del CBRT son típicas de zonas intervenidas en el Valle Central, tales como el yos (Sapium glandulosum L. Morong), guarumos (Cecropia sp.) y targuá (Croton draco Schltdl. y Cham) (Hammel, Grayum, Herrera y Zamora, 2003). Al igual que otros estudios en bosques similares del Valle Central destacó la familia Fabaceae como la más diversa y abundante, entre otras bien representadas como Myrtaceae, Lauraceae y Euphorbiaceae (Cascante- Marín y Estrada-Chavarría, 2012; Villalobos, 2013).
Asimismo, la riqueza (familias y especies) de estos dos estratos fue muy similar a lo reportado en el río Pirro en Heredia; un ecosistema urbano en zonas de vida equivalentes (Villalobos, 2013). Sin embargo, fue considerablemente inferior a lo reportado en la franja de bosques premontanos húmedos y muy húmedos en la Zona Protectora el Rodeo (zonas de vida equivalentes) con 115 especies arbóreas y 40 familias. Cabe resaltar que este último corresponde a uno de los remanentes boscosos más importantes y menos intervenidos de estas zonas de vida en el Valle Central y recibe una mayor influencia de flora del Pacífico seco y la región Sur del país (Cascante-Marín y Estrada-Chavarría, 2012).
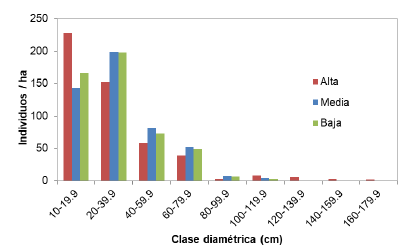
Figure 3 Categorization of trees by diameter classes in different areas altitudinal area of Biological Corridor Tibás River, Heredia, Costa Rica.
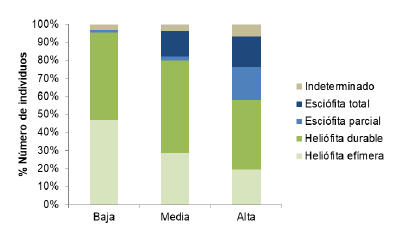
Figure 4 Proportion of individuals according to the ecological guild for the different altitudinal zones of Biological Corridor Tibás River, Heredia, Costa Rica.
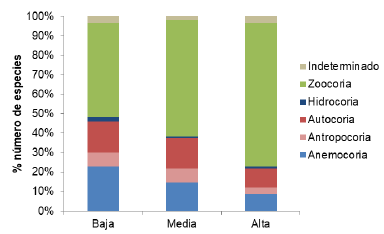
Figure 5 Proportion of species according to the dispersion syndrome for different altitudinal zones of Biological Corridor Intercity Tibás River, Heredia, Costa Rica.
En relación con el estrato alto del corredor, este pertenece a una zona de vida mayormente conservada en el Valle Central, pero se mantiene como un área poco estudiada. No obstante, hay reportes de especies representativas que concuerdan con este estudio, como la dominancia encontrada de arbustos de cafecillo en regeneración (Palicourea sp., Rubiaceae), así como de los árboles lloró (Cornus disciflora DC.), Mauria heterophylla Kunth y Roupala glaberrima Pittier en el dosel (Maglianesi, 2010; Ortega-Rivera y Vílchez-Alvarado, 2013).
Resalta que en los tres estratos una especie exótica es la de mayor peso ecológico (según el I.V.I). El ciprés durante mucho tiempo fue la especie más utilizada en reforestación en el Valle Central del país (Barrientos y Monje-Nájera, 2010) y en el área de estudio su dominancia persiste. Los otros casos más relevantes son el de E. poeppigiana (poró gigante) y Juglans neotropica Diels (cedro nogal), especies típicamente plantadas en sistemas de cafetales (Rojas-Rodríguez, Canessa y Ramírez, 2005) y actualmente corresponden al bosque ribereño en muchas áreas del corredor como cultivos abandonados.
A pesar de ello, es muy probable que estas especies no representen una amenaza seria para el futuro planteamiento del corredor y sucesión ecológica. Las condiciones de regeneración natural encontradas son favorables en términos de riqueza e índices de diversidad y ésta fue muy escasa para especies como el ciprés y poró gigante. Condición que se podría relacionar con la óptima cobertura vegetal, el aumento en la competencia por nutrientes y el espacio (Chaverri, Zamora, Aguilar y Gutiérrez, 1998). La especie que podría representar un riesgo mayor es la invasora R. communis que fue muy abundante en varios sitios de la zona baja, especialmente en áreas más perturbadas.
Por otro lado, se encontró que los estratos altitudinales explican casi un 50 % las diferencias en composición florística, destacando que son tres ecosistemas considerablemente diferentes al menos en cuanto a su composición, asociado en gran parte a diferencias climáticas. No obstante, el grado de intervención urbana y diferencias en porcentajes de cobertura de dosel también podría incidir en este resultado. Finalmente, se resalta que la presencia de varias especies arbóreas endémicas o con algún grado de amenaza refuerza el valor de conservación y mantenimiento de conectividad biológica en estas zonas boscosas.
Estructura arbórea
Un indicador óptimo de sostenibilidad para los tres estratos es el de establecimiento de especies sucesoras según la distribución diamétrica de J invertida encontrada (Olivier y Larson, 1990). Así bien, las densidades de árboles estimadas (entre 469-491 individuos/ha) siguen un comportamiento similar a lo reportado por otros estudios en bosques tropicales premontanos y montanos menos intervenidos en Costa Rica (Wattenberg y Breckle, 1995; Orozco, 1991; Cascante-Marín y Estrada- Chavarría, 2012).
Cuadro 5 Descripción general de los estratos altitudinales en el área destinada al Corredor Biológico Interurbano Río Tibás y resultados obtenidos de densidad de árboles (N) y porcentaje de cobertura vegetal.
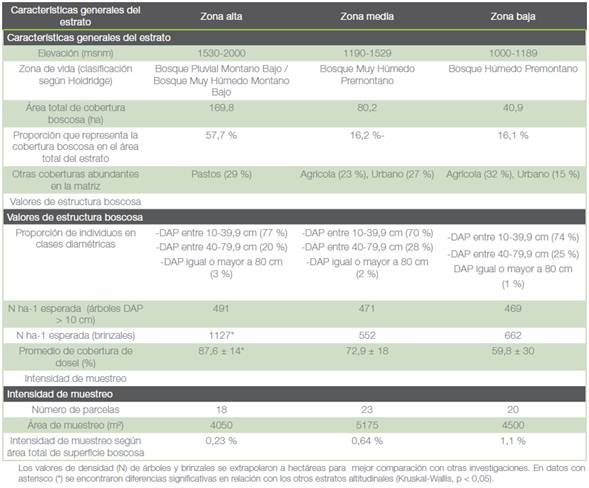
Los individuos excepcionalmente grandes, diámetros mayores a 100 cm, correspondieron a cipreses en plantaciones y árboles remanentes como R. glaberrima. y C. disciflora en la zona alta, o bien a higuerones (Ficus sp.) en las zonas media y baja. Estos últimos pudieron haberse conservado por su valor como sombra o cercas vivas.
Diversidad funcional: Los tres estratos altitudinales presentaron proporciones de clases diamétricas e indicadores de diversidad y equidad similares. No obstante el porcentaje de cobertura de dosel y la densidad de brinzales fueron significativamente mayores en la zona alta, que destaca como la menos perturbada. Dicha condición también se refleja en cambios detectados en la proporción de gremios ecológicos y síndromes de dispersión de las especies. Hacia la parte alta se encontró menor proporción de heliófitas efímeras, especies pioneras demandantes de mucha luz, debido al cierre del dosel y mayor proporción de plantas de sombra o esciófitas, características de sucesión tardía (Pütz Groeneveld, Alves, Metzger y. Huth, 2011).
Este aumento de especies esciófitas es indicador de mayor sostenibilidad del ecosistema y una posible convergencia hacia bosques primarios (Ruschel, Mantovani, Sedrez y Onofre, 2009; Higuchi et al. 2008). En las zonas media y baja, la disminución de cobertura de dosel y aumento de claros dentro del bosque favorece la colonización de especies heliófitas y anemócoras, típico de bosques sometidos a frecuentes perturbaciones (Bocanegra-González, K. T., Fernández-Méndez, F. y Galvis-Jiménez, J. (2015).
Estas proporciones de gremios ecológicos son similares a lo reportado por Villavicencio (2012) en un Bosque Muy Húmedo Premontano Bajo en la cuenca del río Aranjuez, Puntarenas. En éste, las especies heliófitas representaron hasta el 90 % en zonas intervenidas, porcentaje que bajó a cerca del 70 % en la zona más alta y menos perturbada de la cuenca. Se ha encontrado también que este grupo de esciófitas es particularmente más sensible a la fragmentación por lo que en las zonas media y baja del corredor se refuerza la necesidad de mejorar las condiciones de cobertura del dosel y conectividad del paisaje (Metzger, 2000).
En relación con los síndromes de dispersión, generalmente en los bosques tropicales húmedos más del 50 % de las especies son dispersadas por animales, condición que se encontró en los tres estratos del CBRT (Howe y Smallwood, 1982; Di Stéfano, Nielsen, Hoomans y Fournier, 1996). No obstante, en la zona alta se registró mayor proporción de zoocoria (cerca de 75 %) y disminución de la anemocoria, lo cual se asocia con un mejor estado de conservación (Sandor, 2012; Bocanegra-Gonzáles, et al., 2015).
La representación de diversos grupos funcionales encontrados en los tres estratos, según gremios ecológicos y síndromes de dispersión, indican alguna capacidad de resiliencia ante perturbaciones. No obstante mayores proporciones de especies esciófitas y zoócoras son ideales (Folke et al., 2004; Bocanegra- González et al. 2015).
En la zona baja del CBRT cerca del 50 % de individuos corresponden a heliófitas efímeras. Esto puede afectar negativamente la sustentabilidad del ecosistema, especialmente en sitios con una matriz antropogénica donde dominan usos agrícolas y urbanos. Los bosques están expuestos a mayor frecuencia de aperturas de claros y muchas especies suelen ser poco competitivas, por lo que son susceptibles a agentes externos como plagas y enfermedades (Lozano-Zambrano, 2009; Ceccon, 2014). En este estudio se observó una considerable mayor afectación por el pasto gigante Pennisetum purpureum Schumach en ecosistemas de la zona baja; una especie invasora que limita seriamente la regeneración natural e hidrología del río (Lambert, Dudley y Saltonstall, 2010), situación que podría verse favorecida por la condición mencionada.
Conclusiones
La composición florística difirió hasta un 61% entre las tres zonas altitudinales. En las zonas media y baja resaltó la familia Fabaceae como la más diversa y abundante, entre otras bien representadas como Myrtaceae, Lauraceae y Euphorbiaceae, con muchas especies típicas de bosques intervenidos. Mientras que en la zona alta destacaron mayormente familias como Melastomataceae, Rubiaceae y Cupressaceae.
Los tres estratos altitudinales mostraron índices de diversidad de Shannon mayores a 3.1 y alta equidad en la composición de especies según el índice de Pielou. Asimismo la distribución diamétrica encontrada sugiere que se encuentran en recuperación y es indicadora de procesos adecuados de reclutamiento.
No obstante en la zona alta se concentra cerca del 58 % de la cobertura boscosa del área del CBRT y ésta destaca como la menos perturbada, con mayor proporción de especies esciófitas, zoócoras, porcentajes de cobertura en el dosel cercano al 87 % y mayor densidad de brinzales (hasta un 51 % más alta que otros estratos). Las zonas media y baja reflejaron mayor grado de perturbación debido a su moderada y baja cobertura de dosel (72,9 ± 18 y 59,8 ± 30 respectivamente) y proporciones altas de especies heliófitas efímeras y anemócoras.
Recomendaciones
En conjunto con las acciones propuestas para mejorar la conectividad en la zona destinada al CBRT, es importante manejar zonas boscosas de los estratos medio y bajo a través de enriquecimiento, utilizando especies nativas heliófitas durables y esciófitas. Esto con el fin de contrarrestar la dominancia encontrada por especies exóticas y heliófitas efímeras, que pueden incidir negativamente en la sustentabilidad. Asimismo es importante localizar y manejar las principales zonas del CBRT afectadas por la invasión de pasto gigante, para permitir procesos óptimos de sucesión ecológica.

