











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkEn sistemas de producción de rumiantes, en donde el forraje se emplea en ciclos constantes de ocupación y descanso (pastoreo rotacional), permite un uso sostenible de este recurso (Gutiérrez, 1996). A nivel de sistema radical, está dinámica de pastoreo y recuperación de la pastura provoca la descomposición de tejido de las raíces durante el periodo interpastoreo (Georgieva, Chritensen & Stevnbak, 2005) que puede influir en las poblaciones de nematodos fitoparásitos, que se caracterizan por ser endoparásitos obligados e inclusive los ectoparásitos y los nematodos no fitoparásitos.
Otro aspecto del pastoreo rotacional que podría afectar las poblaciones de nematodos, es la deposición de excretas (boñiga y orina) en el aparto, las cuales son fuentes de materia orgánica y formas solubles de ni trógeno y otros elementos (Beetz, 2002). El impacto de estas deposiciones sobre la actividad microbiológica del suelo esta documentada (Wang, Mc Sorley, Bohlen & Gathumbis, 2006), pero la intensidad de la misma de penderá de la carga animal instantánea que se maneje en ese momento, debido a que la cantidad de excretas se relaciona a la cantidad de animales en pastoreo. Estas deposiciones de nutrimentos de forma periódica, pue den favorecer el crecimiento de microorganismos que presenten actividades depredadoras o de control sobre la población de nematodos fitoparásitos (Bardgett, Cook, Yeates & Denton, 1999).
Por último, las condiciones climáticas que se presen ten durante estos ciclos de pastoreo, podrían modificar el comportamiento de los animales, a nivel de consumo de forraje y en el desplazamiento (Bilotta, Brazier & Haygart, 2007), las condiciones de suelo, su temperatura y de cre cimiento del forraje. Factores que afectan el crecimiento y la dinámica de las poblaciones de nematodos fitopará sitos y no fitoparásitos. Por tal motivo el objetivo de este trabajo fue valorar la dinámica poblacional durante un año calendario de las poblaciones de nematodos fitopa rásitos y no fitoparásitos que se encuentren presentes en sistemas de producción de leche que aplican el pasto reo en áreas con cobertura permanente de pasto Estrella africana, San Juan, Tanner y Ratana.
Metodología
Localización de los sistemas. El área que se utilizó en este trabajo para determinar la dinámica poblacional de los nematodos fitoparásitos y no fitoparásitos en los pastos Estrella africana (Cynodon nlemfuensis), San Juan (Setaria anceps), Tanner (Brachiaria radicans) se ubica en la comunidad de Sucre de Ciudad Quesada, Costa Rica (10°19'25"N, 84°25'37"W). Esta misma área fue muestreada por WingChing-Jones, Salazar-Figueroa, Flores-Chavez y Rojas-Bourrillón (2008) para la identificación de nematodos en pastos de altura, con la variante en área de muestreo, como se describe en el Cuadro 1. En cambio, el pasto Ratana (Ischaemun ciliare) se ubicó en la locali dad de Jabillo de Ciudad Quesada, área muestreada por WingChing-Jones y Salazar-Figueroa (2008), en donde se identificó la nematofauna encontrada en este pasto en condiciones de bajura.
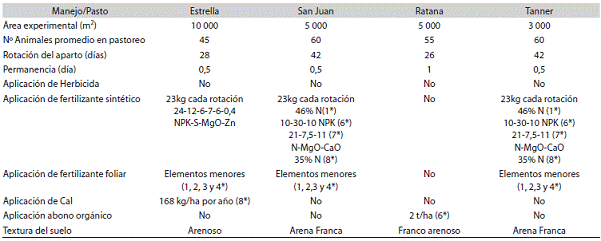
Cuadro 1: Descripción del área de los potreros utilizados, como el número de animales que pastorean, la rotación entre pastoreos y la aplica ción de productos sintéticos según el mes del año. Ciudad Quesada, San Carlos, Alajuela, Costa Rica, 2006
Caracterización del muestreo y manejo de la mues tra. Se trabajó con cuatro muestras compuestas de suelo por pasto recolectadas de forma aleatoria. Está se tomó a una profundidad de 0 a 20cm con ayuda de un palín, cada muestra pesó dos kg, producto de la toma de cinco submuestras en el área delimitada para la prueba, para un total de veinte puntos, cada punto de muestreo con tiene suelo como las raíces contenidas en el volumen de suelo. El muestreo se realizó cada 30 días durante doce meses de investigación (Enero 2004-Enero 2005), con la presencia de periodos de pastoreo y recuperación de las pastura, al inicio del mismo, por motivos de coordinación se respeto el ámbito de tres días antes y tres días después al momento exacto (inicio de la rotación) como óptimo para la toma de la muestra.
Manejo de la muestra. Luego de ser identificada la muestra, se traslado al Laboratorio de Nematología de la Universidad de Costa Rica, en donde la muestra de suelo y raíces se homogenizo y cuarteo, para ser procesada por el método de cernido y centrifugado en solución azuca rada (Alvarado & López, 1985), la lectura de las mismas se realizó con la ayuda de un microscopio estereoscopio a 45X, a nivel de género (Maggenti, 1991). El número de nematodos obtenido en las muestras de suelo como en la de raíces en este trabajo, se expresan en el docu mento como el logaritmo en base de 10 del número de nematodo por género obtenido en 100cc de suelo o 100g de raíces (log10 N° nematodos en 100g de raíces o 100cc de suelo).
Información meteorológica. Los datos de precipi tación empleados en este trabajo fueron suministrados por el Instituto Meteorológico Nacional, el cual por me dio de sus estaciones ubicadas en San Vicente y en Santa Clara, registra la precipitación de forma diaria, expresada en mililitros por día.
Análisis de los datos. Se determinó la significancia de los efectos principales por medio del modelo ANOVA de SAS (2003) para las poblaciones de nematodos en el sustrato (raíz o suelo), momento del muestreo y la in teracción sustrato con la época. En el momento que se detectó efectos significativos de los efectos principales, se realizó la prueba que compara las medias según los rangos establecidos para la prueba de Waller-Duncan (p<0,05).
Resultados
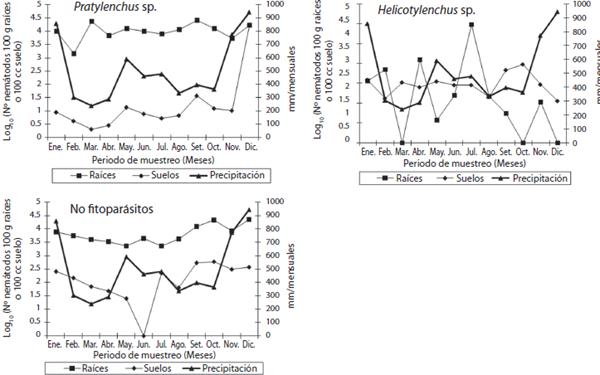
Pasto Estrella africana (C. nlemfuensis). Para Pratylenchus sp. la variable sustrato fue significativa (p<0,0001), donde se determinó una mayor cantidad en las muestras de raíces que en las de suelo (1,02 contra 0,16 respectivamente (log10 N° nematodos en 100g de raíces o 100cc de suelo)), no así el momento de toma de la muestra ni la interacción momentoXsustrato (Figura 1). Se determinó que la densidad poblacional en el suelo en este estudio es 6,37 veces menor a la densidad obtenida en las raíces. En el caso de Helicotylenchus sp., el sustra to es una variable importante (p=0,0009) y la interac ción entre el sustrato y el mes de muestreo (p<0,01). Se notó una mayor cantidad de nematodos en las muestras de raíces que en el suelo que rodea las raíces del pasto Estrella africana (2,36 contra 1,76 respectivamente (log10 N° nematodos en 100g de raíces o 100cc de suelo)) (Figura 1). Esta mayor población se observa cuando se da una reducción en la precipitación mensual de 900mm a 350mm y sufre un descenso al aumentar la precipitación. La población de nematodos de este género presenta la menor cantidad de individuos por gramo de raíces du rante los meses de enero y diciembre, con respecto a los otros meses, los cuales presentan una población prome dio que varia significativamente. Para Tylenchus sp., su dinámica poblacional se ve afectada significativamente por el sustrato, el momento del muestreo y la interacción sustratoXmes (p<0,001) (Figura 1). La población registrada en las raíces supera en 2,97 veces el número de indivi duos presente en las muestras de suelo (1,28 contra 0,43 respectivamente (log10 N° nematodos en 100g de raíces o 100cc de suelo)). En este caso, se nota incrementos en las poblaciones a principios y a finales del año y el mes de mayo como el momento en el cual se registró la menor población de individuos de este género. Respecto a los individuos del grupo de los nematodos no fitoparásitos (Figura 1), se obtuvo relevancia significativa para el sustrato, el momento del muestreo y a la interacción sustratoX-mes (p<0,0001). En este grupo, la población determina da en las raíces supera en 1,87 veces la población en el suelo que rodea las raíces (3,87 contra 2,07 respectiva mente (log10 N° nematodos en 100g de raíces o 100cc de suelo)). También se observa una tendencia de cuantificar mayores conteos de individuos a finales del año, lo cual se podría relacionar a un aumento en la precipitación pluvial. De forma general, al analizar las poblaciones de los nematodos fitoparásitos en las raíces, se nota que la mayor población promedio de individuos es registrada para el género Tylenchus sp. (3,87 log10 N° nematodos en 100g de raíces), seguida de Helicotylenchus sp. (2,36 log10 N° nematodos en 100g de raíces) y por último, Pratylenchus sp. (1,01 log10 N° nematodos en 100g de raíces). Además de los géneros evaluados, se presentó en las muestras de raíces de forma ocasional poblacio nes promedio de nematodos (datos o rango entre pa réntesis) de Criconemella sp. (1,78), Hemicycliophora sp. (1,60), Ditylenchus sp. (1,3-2,0) y Xiphinema sp. (2,38-3,42) (log10 N° nematodos en 100g de raíces). Mientras que en las muestras de suelo, se cuantificó Criconemella sp. (0,17-1,62), Hemicycliophora sp. (0,60-2,07), Xiphinema sp. (1,25), Meloidogyne sp. (1,01), Paratylenchus sp. (0,47 1,55) y Criconema sp. (0,54-1,25) (log10 N° nematodos en 100cc de suelo).
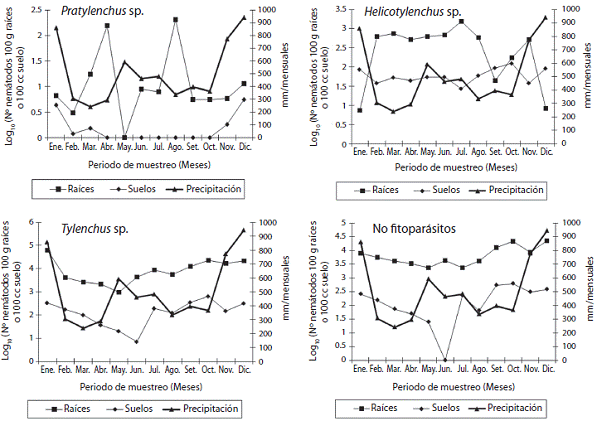
Figura 1: Fluctuación en la población de nematodos de los géneros Pratylenchus sp., Helicotylenchus sp., Tylenchus sp. y no fitoparásitos presente en muestras de raíces y de suelo en el pasto Estrella. Cuidad Quesada, San Carlos, Alajuela, Costa Rica, 2006.
Pasto Ratana (I. ciliare). Se determinó para el grupo de nematodos no fitoparásitos diferencias significativas debidas al sustrato, el momento del muestreo y a la in teracción (p<0,0001). En las raíces se determinó el doble de individuos presentes en las muestras de suelo, don de se obtuvo poblaciones de 3,53 contra 1,76 respecti vamente (log10 N° nematodos en 100g de raíces o 100cc de suelo). Con respecto al momento de la toma de la muestra se determina setiembre y diciembre como los meses que registraron la población mayor, mientras que mayo y julio, se describen los valores menores (Figura 2). En relación a Meloidogyne sp., solo se determinó diferencias entre las poblaciones presentes en las raíces y el suelo (p<0,05), donde los individuos en las muestras de raíces son 2,56 veces más a las presentes en las muestras de suelo (2,97 contra 1,16 respectivamente (log10 N° nema-todos en 100g de raíces o 100cc de suelo)). Se determinó octubre, el mes con la mayor población de Meloidogyne sp., mientras que, se obtuvo la menor población en el mes de abril. El sustrato (p<0,0001), el momento del muestreo (p<0,0015) y la interacción sustratoXmomento (p<0,009), presentó efecto directo sobre la población de Pratylenchus sp.. En el caso del sustrato, la población contabilizada en las raíces es 3,02 veces más alta que la encontrada en el suelo (2,51 contra 0,83 respectivamen te (log10 N° nematodos en 100g de raíces o 100cc de sue lo)). El mes donde se determinó la menor población fue febrero, en cambio, para marzo y setiembre se obtuvo las mayores poblaciones (Figura 2). Otro nematodo determina do en pasto Ratana fue Paratylenchus sp., cuya población fue afectada por el sustrato, el momento del muestreo y la interacción evaluada (p<0,001). Las raíces presentan 3,94 veces más individuos que los contabilizados en el suelo (3,47 contra 0,95 respectivamente (log10 N° nema-todos en 100g de raíces o 100cc de suelo)). En relación a este nematodo, durante los meses de enero, abril y mayo, no se determinó ningún individuo en las muestras anali zadas, comportamiento que no se podría explicar, debi do en que los meses restantes, se presentan poblaciones en raíces superiores a 4 (log10 N° nematodos en 100g de raíces). En el caso de los individuos en suelo, no se obtie nen valores superiores a 2 (log10 N° nematodos en 100cc de suelo). De forma ocasional se determinaron (datos o rango entre paréntesis) en las muestras de raíces y de suelo nematodos del género Tylenchus sp. (2,38-2,48 y 0,10-1,32 respectivamente (log10 N° nematodos en 100g de raíces o 100cc de suelo)) y Criconemella sp. (1,30-2,38 y 0,90-1,62 respectivamente (log10 N° nematodos en 100g de raíces o 100cc de suelo)). En el caso de Helicotylenchus sp., éste se encontró durante los 12 muestreos, tanto en las muestras de raíces y del suelo contenido en la mues tra, con poblaciones promedio de 1,3-2,91 y de 1,17-2,26 respectivamente (log10 N° nematodos en 100g de raíces o 100cc de suelo). Las poblaciones en las raíces fueron 1,46 veces mayores que las encontradas en el suelo (p<0,05) para este nematodo (1,21 contra 0,83 respectivamente (log10 N° nematodos en 100g de raíces o 100cc de suelo)).
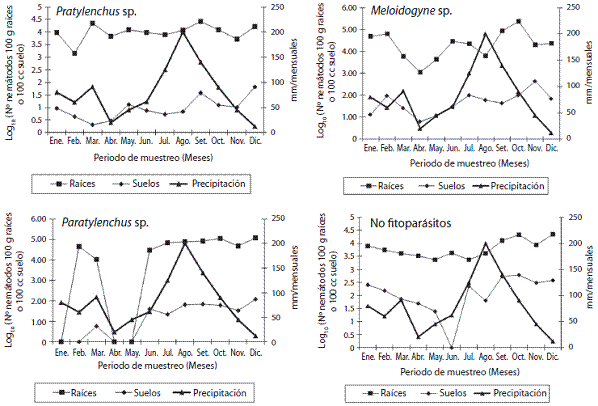
Figura 2: Fluctuación en la población de nematodos de los géneros Pratylenchus sp., Meloidogyne sp., Paratylenchus sp. y no fitoparási tos presente en muestras de raíces y de suelo en el pasto Ratana. Cuidad Quesada, San Carlos, Alajuela, Costa Rica, 2006.
Pasto San Juan (S. anceps). Para este forraje, el ne-matodo Pratylenchus sp. presentó efectos significativos para el sustrato (p<0,0001) y la interacción sustratoXmo-mento del muestreo (p<0,005). Se obtuvo una relación de 5,73 nematodos en las raíces por cada nematodo en las muestras de suelo (2,92 contra 0,51 respectivamente (log10 N° nematodos en 100g de raíces o 100cc de sue lo)). Los meses de enero, abril, setiembre y noviembre se contabilizó la mayor cantidad de individuos en las mues tras de raíces y el mes de mayo con la población menor (Figura 3). En el caso de las muestras de suelo analizadas, las poblaciones obtenidas presentan un comportamien to similar a la variación observada en las poblaciones en raíces. La población de Helicotylenchus sp. varió significa tivamente por efecto del sustrato (p<0,0008), el momen to de la toma de la muestra ((p<0,0002) y la interacción sustrato X momento de muestreo (p<0,04). La población de Helicotylenchus sp. cuantificada en raíces supero en 1,50 veces la población de nematodos en las muestras de suelo (2,02 contra 1,35 respectivamente (log10 N° ne-matodos en 100g de raíces o 100cc de suelo)). En este forraje, se nota una tendencia entre la población de ne-matodos en las raíces y la precipitación registrada. Para los nematodos no fitoparásitos, las variables evaluadas presentaron una alta significancia (p<0,0001). En el caso del sustrato, la población en las raíces fue de 3,57 (log10 N° nematodos en 100g de raíces) mientras que las de terminadas en las muestras de suelo alcanzó un valor de 2,01 (log10 N° nematodos en 100cc de suelo). Se nota un comportamiento similar en las poblaciones de nemato-dos en las raíces y el suelo alrededor de las raíces, según el patrón de las lluvias en cada mes, como se describió antes, lo que se podría relacionar con el crecimiento del forraje. El nematodo Tylenchus sp., también se encontró de forma ocasional en muestras de suelo y de raíces en el pasto San Juan, su valor o rango promedio durante el periodo experimental, tanto en el suelo como en las raí ces fue de 0,51 a 1,73 y de 1,14 a 3,42 respectivamente (log10 N° nematodos en 100g de raíces o 100cc de suelo). Mientras que para Ditylenchus sp., solo se observo en las muestras de suelo (1,07 a 1,51 (log10 N° nematodos en 100cc de suelo)) y Xiphinema sp. con un valor de 3,15 in dividuos en las muestras de raíces (log10 N° nematodos en 100g de raíces).
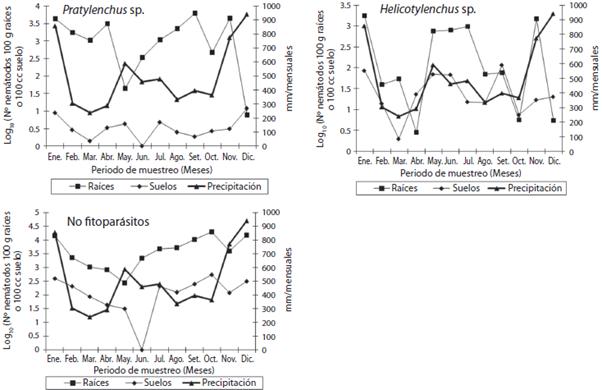
Figura 3: Fluctuación en la población de nematodos de los géne ros Pratylenchus sp., Helicotylenchus sp. y no fitoparásitos pre sente en muestras de raíces y de suelo en el pasto San Juan. Cuidad Quesada, San Carlos, Alajuela, Costa Rica, 2006.
Pasto Tanner (B. radicans). En esta investigación, la población de Pratylenchus sp. presente en las raíces di fiere significativamente con respecto a los individuos determinados en las muestras de suelo (p<0,0001), en una relación de 3,43:1 (3,99 contra 1,16 respectivamente (log10 N° nematodos en 100g de raíces o 100cc de suelo)). Estas poblaciones no fueron afectadas por el momento en que se tomó la muestra y la interacción sustratoXmo-mento de toma de la muestra (p>0,05). La evaluación de las poblaciones del nematodo Helicotylenchus sp. en esta investigación, determina que el sustrato (p<0,03), el mo mento de toma de la muestra (p<0,01) y la interacción sustratoXmomento de toma de la muestra (p<0,001) son significativas. Se determina una relación de 1,34 nematodos en las muestras de raíces por cada nemato-do en el suelo (1,47 contra 1,10 respectivamente (log10 N° nematodos en 100g de raíces o 100cc de suelo)). El mes de julio obtuvo la mayor población de nematodos en raíces, mientras que, se registra marzo, octubre y di ciembre, como los meses con la menor población. En el caso de las poblaciones en las muestras de suelo, estás presentan una constancia en cada muestreo, que fluctúa entre 1 y 2 (log10 N° nematodos en 100cc de suelo) (Figura 4). Para los nematodos no fitoparásitos, se determinó que el sustrato, el momento de toma de la muestra y la interacción sustratoXmomento de toma de la muestra afectan significativamente las poblaciones de nemato dos en el pasto Tanner (p<0,0001). En el caso de las den sidades en las muestras de raíces se determinó un valor de 3,79 nematodos (log10 N° nematodos en 100g de raí ces), mientras que, para las muestras de suelo la densi dad fue 1,88 veces menor (2,02 log10 N° nematodos en 100cc de suelo). En este forraje, la población de nematodos no fitoparásitos presentó mayor uniformidad, donde se considera octubre y julio como los meses con mayor y menor población de nematodos respectivamente. Otros géneros de nematodos que se determinó de forma oca sional en las muestras de suelo y de raíces fue Tylenchus sp., Criconemella sp. y Paratylenchus sp.. Para cada género se determinó rangos promedios de poblaciones en las muestras de raíces de 2 a 2,36, de 1,30 a 2,98 y de 2,68 a 3,46 respectivamente (log N° nematodos en 100g de raíces). En cambio, los conteos que se obtienen en las muestras de suelo alrededor de las raíces fluctuaron entre 0,1 a 1,59; 0,86 a 2,44 y 1,75 a 1,89 para Tylenchus sp., Criconemella sp. y Paratylenchus sp. respectivamente (log N° nematodos en 100cc de suelo). En el caso de Ditylenchus sp, este género solo se identificó en las muestras de raíces, con una población entre 2,08 a 2,41 individuos (log N° nematodos en 100g de raíces), caso contrario con Xiphinema sp., que se cuantificó solo en las muestras de suelo con una población de 0,17 individuos por cada 100cc de suelo (log10 N° nematodos en 100cc de suelo).
Discusión
Pasto Estrella Africana. Con respecto al género Pratylenchus sp, las densidades de nematodos podría explicarse por el hábito de alimentación que presentan los nematodos de este género, los que, al ser endoparásitos, se encontraban en proceso de alimentación en el momento de la toma de la muestra (Norton, 1978). En el caso, de la relación entre raíces y suelo, el proceso de migración de los nematodos hacia la fuente de alimenta ción (raíces) podría ser una explicación (Yeates, Bongers, de Goede, Freckman & Georgieva, 1993). En el caso de este género, WingChing-Jones et al. (2008) en la misma área informa de poblaciones en raíces de 2,46 individuos (log N° nematodos en 100g de raíces nematodos) en tre los meses de mayo a setiembre, población 2,5 veces mayor a la determinada en este estudio, para el mismo periodo. Además, un comportamiento diferente de la precipitación y del cultivo, de la dinámica del pastoreo rotacional, del aporte de nutrimentos y de materia orgá nica por las deposiciones de la boñiga y la orina, podrían ser variables que afectan la población de nematodos (Liang et al., 2009). Para el nematodo Helicotylenchus sp. los resultados se podrían relacionar al comportamien to ectoparásito migratorio o semiendoparásito cuando se encuentra en procesos de alimentación (Davis, Bell, Watson & Rohan, 2004), donde al momento de la toma de la muestra, este nematodo se encontraba alimentándose de las raíces. En este mismo sistema de producción, WingChing-Jones et al. (2008), informa de poblaciones similares de 2,83 individuos (log10 N° nematodos en 100g de raíces) durante los meses de mayo a setiembre. Variación que se podría relacionar a la disponibilidad de alimento, humedad y temperatura en el suelo, va riables propias del sistema, según la época de toma de la muestra (Mc Groary, Crow, McSorley, Giblin-Davis & Cisar, 2009). Las poblaciones en raíces y suelo para el gé nero Tylenchus sp. podría relacionarse a la dinámica del cultivo bajo un sistema de manejo de forma rotacional, que favorece o perjudica la población de nematodos de este género. Al comparar los resultados obtenidos en este trabajo para este género, con los informados por WingChing-Jones et al. (2008), en donde se obtiene una población de 2,21 individuos (log10 N° nematodos en 100g de raíces) durante los meses de mayo a setiembre, población 1,72 veces mayor a la obtenida en esta inves tigación. Comparación, que fortalece la hipótesis, de que las poblaciones de este género podrían ser afectadas por variables climáticas y de manejo del sistema. En el caso de los especímenes determinados en las muestras de raíces para los nematodos no fitoparásitos, se podría relacionar a que en el momento de la toma de la mues tra, estos se encontraban en proceso de alimentación en áreas de las raíces no funcionales. Además, se determina una población 1,16 veces mayor en esta investigación, a la determinada por WingChing-Jones et al. (2008) duran te los meses de mayo a setiembre. Lo que se podría rela cionar a un cambio de las condiciones bióticas y abióticas del sistema. El comportamiento obtenido para las pobla ciones de los géneros Helicotylenchus sp. y Pratylenchus sp. podría indicar, que el ambiente generado en la rizos-fera de este cultivo, provoca que estos géneros, no se de sarrollen de forma adecuada al compararse a Tylenchus sp. En el caso de los nematodos, que se determinaron en forma ocasional, en las muestras de suelo y raíces, los resultados concuerdan con lo descrito por WingChing-Jones et al. (2008), cuando evaluó este mismo sistema de producción. Comportamiento que se podría relacionar, a las condiciones que genera esta pastura en la rizosfera, la cual, no propicia el desarrollo de estos géneros de nematodos (Mazzola, 2007)
Pasto Ratana. El comportamiento obtenido para los nematodos no fitoparásitos no permite obtener una tendencia o relación con la precipitación promedio para esos meses con la población de nematodos. Al comparar la dinámica de precipitación y el comportamiento de la población de este nematodo, se describe una tendencia a disminuir la población de individuos al aumentar la pre cipitación. En el caso de este género, WingChing-Jones y Salazar-Figueroa (2011), informa de poblaciones de 4,99 individuos (log10 N° nematodos en 100g de raíces) en este forraje, lo que indica, una alta capacidad de este forraje para cumplir funciones de hospedero. Las diferencias entre las poblaciones determinadas en raíces y el suelo, se podrían relacionar a las características de endoparásito que presenta los especímenes de este género. En un estudio previo, WingChing-Jones y Salazar-Figueroa (2011), informa de poblaciones de 3,22 individuos (log10 N° nematodos en 100g de raíces) para este forraje, en condiciones similares de manejo y de clima. Lo cual in dica, un posible efecto de las prácticas de manejo de los animales como se indicó en el pasto Estrella, por efecto de la adición de materia orgánica durante el pastoreo. En el primer trimestre del año, se nota una tendencia simi lar entre la población de nematodos en raíces y la preci pitación, lo cual se podría relacionar con el crecimiento del forraje, durante los periodos de menor precipitación, posterior a este evento, la población de nematodos se mantiene constante. En el caso de los nematodos del género Paratylenchus sp., los resultados obtenidos concuerdan a lo informado por WingChing-Jones y Salazar-Figueroa (2011), en donde describen poblaciones de 4,30 individuos (log10 N° nematodos en 100g de raíces) en este mismo sistema de producción. En el caso de este forraje, llama la atención las poblaciones de nematodos fitoparásitos y no fitoparásitos, superiores a 1 000 indivi duos por cada 100g de raíces analizadas, lo que podría indicar que el Ratana no presenta una susceptibilidad a estas cargas parasitarias o estas cargas son responsables a la baja producción de biomasa (kg/ha) de esta pastura (Villareal,1992).
Pasto San Juan. Para el género Pratylenchus sp. los resultados obtenidos en esta investigación presentan similitud con lo descrito por WingChing-Jones et al. (2008), donde informan de una población de 3,22 individuos (log10 N° nematodos en 100g de raíces) durante los me ses de mayo a setiembre. La tendencia obtenida entre los nematodos en raíces y el suelo alrededor de las raíces para el género Helicotylenchus sp., podría estar relacio nada a la tasa de crecimiento del forraje (kg/ha de biomasa), la cual promueve un crecimiento radical, lo que genera una mayor disponibilidad de alimento. La pobla ción de individuos determinada en esta investigación para el género Helicotylenchus sp. fue 1,42 veces menor a la informada por WingChing-Jones y Salazar-Figueroa (2008) cuando evaluó este sistema de producción duran te los meses de mayo y setiembre, variación relacionada a la dinámica del cultivo de forraje, producto al sistema rotacional de pasturas y a las condiciones climáticas im perantes en cada investigación. En el caso de las pobla ciones de individuos para los nematodos no fitoparásito, se encontró una similitud a las descritas por WingChing-Jones y Salazar-Figueroa (2008), lo que indica, una rela ción de 1,78 nematodos en raíces por cada nematodo contabilizado en el suelo.
Pasto Tanner. Al comprar las poblaciones obte nidas en esta investigación con lo informado por WingChing-Jones y Salazar-Figueroa (2008) para el gé nero Pratylenchus sp. (3,64 (log10 N° nematodos en 100g de raíces), se determina una similitud en los resultados obtenidos, lo cual podría indicar, que este forraje es un buen hospedero para nematodos de este género. Para el género Helicotylenchus sp. se determinaron poblacio nes de nematodos 1,65 veces menor a las obtenidas por WingChing-Jones y Salazar-Figueroa (2008) para este género de nematodos en el mismo forraje y en la mis ma localidad. Lo cual podría indicar, que las condiciones de manejo de la finca y las condiciones climáticas, po drían afectar estas poblaciones. Los valores obtenidos en esta investigación para los nematodos no fitoparásitos, concuerdan con los descritos por WingChing-Jones y Salazar-Figueroa (2008) para este forraje, donde informó poblaciones de 3,39 individuos (log10 N° nematodos en 100g de raíces), lo cual, se podría relacionar con el reci claje de la materia orgánica, producto al pastoreo y la de posición de las excretas y la orina en este sistema.

