











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
La base estructural de cualquier arrecife o comunidad coralina es resultado, principalmente, de la presencia y crecimiento de corales hermatípicos o pétreos, los cuales forman colonias sólidas y tridimensionales que proveen un servicio de soporte a la biodiversidad y biomasa, tanto de vertebrados como de invertebrados asociados a estos ecosistemas coralinos (Graham & Nash, 2013). El coral funciona como un holobionte y depende de la relación simbiótica de su población de dinoflagelados endosimbintes y bacterias que fijan carbono como producto de desecho de su respiración celular (van Oppen & Blackall, 2019). Como resultado se fija hasta un 200 % de energía que es traslocada al coral y utilizada por la colonia para su mantenimiento basal y procesos de alto costo energético como el crecimiento y reproducción (Roth, 2014). Por lo tanto, el funcionamiento óptimo de esta relación simbiótica es esencial y dependiente de los factores ambientales ligados a la fisiología del holobionte, siendo la temperatura superficial del mar (TSM) y la luz las consideradas hasta la fecha como parámetros más relevantes (van Oppen & Blackall, 2019).
El efecto sinérgico e intensificado de estresores de tipo natural (p.ej. eventos El Niño Oscilación del Sur (ENOS), huracanes, afluentes pluviales) y antropogénico (p. ej. sobre-pesca, contaminación, aporte de nutrientes), han llevado en los últimos años al colapso y eventual pérdida de arrecifes enteros alrededor del mundo (Hughes et al., 2018; Souter et al., 2020). En respuesta, se han adoptado diferentes medidas de mitigación, entre ellas los métodos de restauración asistida, con el objetivo principal de incrementar la cobertura de coral vivo, contribuir a la supervivencia de la comunidad arrecifal y a la funcionalidad del ecosistema (Boström-Einarsson et al., 2020). Adicionalmente, ha permitido generar aproximaciones sobre evolución asistida, proporcionando un nuevo conocimiento en la capacidad de respuesta de la comunidad coralina ante la intervención humana (Sugget & van Oppen, 2022).
La región del Pacífico Oriental Tropical (POT), en términos ambientales se ha caracterizado como un sitio sub-óptimo para el crecimiento y mantenimiento de arrecifes coralinos (Glynn & Ault, 2000). A pesar de esto, ha desarrollado comunidades coralinas consideradas de tipo mono-especie por su alta cobertura de coral del género Pocillopora, y la presencia en menor abundancia de especies del género Porites y Pavona. Históricamente, las comunidades coralinas del POT han sufrido eventos de blanqueamiento masivo, y de manera particular en el Pacífico Central Mexicano (PCM), Pocillopora ha sido señalado como el género más sensible al estrés, pero con mayor capacidad de recuperación, evidenciado por una baja mortalidad en años recientes (Rodríguez-Troncoso et al., 2016; Romero-Torres et al., 2020). Por lo que, la capacidad de resistencia de cada especie de coral ante los diferentes estresores, así como de su recuperación posterior a una perturbación, es un factor clave no solo para la supervivencia, sino para su continuo crecimiento y el mantenimiento de la comunidad (Hughes et al., 2018; Romero-Torres et al., 2020). Sumado a esto, se ha evidenciado que cada especie coralina ha desarrollado una respuesta diferencial dependiendo de las condiciones locales (Cruz-García et al., 2020; Martínez-Castillo et al., 2020), por lo que su tasa de crecimiento y mortalidad podría ser variable, aún dentro de una misma región.
La supervivencia de una comunidad coralina depende de las acciones de manejo que se implementen para su conservación, desde mitigación de los estresores hasta la aplicación de acciones para su restauración o remediación (Boström-Einarsson et al., 2020). El éxito de un programa de restauración por propagación asexual está basado principalmente en atributos biológicos intrínsecos como el crecimiento y la supervivencia del fragmento de coral o recluta (Sugget & van Oppen, 2022), y para el caso particular del Pacífico Central Mexicano, se ha considerado la respuesta ante factores ambientales locales, y su potencial de mantenimiento a largo plazo (Martínez-Castillo et al., 2022; Tortolero-Langarica et al., 2014; Tortolero-Langarica et al., 2019). El objetivo de este estudio es evaluar la tasa de crecimiento y supervivencia de coral del género Pocillopora en dos localidades (insular y costera) dentro del PCM, y su relación con la temperatura superficial del mar (TSM), radiación fotosintéticamente activa (PAR, por sus siglas en inglés) y el coeficiente de atenuación lumínica a 490 nm (Kd490). Los resultados permitirán contar con una línea base de información que permita diseñar mejores estrategias y extender los esfuerzos de restauración a lo largo de la región del Pacífico Mexicano.
Materiales y métodos
Área de estudio: El estudio se llevó a cabo en el Parque Nacional Islas Marietas (PNIM; 20°40'35” - 20°41'45” N, 105°33'30” - 105°38'10” W) y, en la Caleta de Cuastecomates (CC; 19°13'29”-19°14'18” N & 104°43'45”-104°45'29” W), ambos localizados en la región del PCM con una distancia aproximada de 185 km entre cada localidad (Fig. 1). La región se caracteriza por ser una zona de transición oceanográfica, en donde la Corriente de California y la Corriente Costera de Costa Rica interactúan formando la Corriente Nor-ecuatorial (Portela et al., 2016). La actividad de remolinos formados a partir de los vientos alisios sobre la Zona de Convergencia Intertropical provoca cambios estacionales en la TSM, presencia de surgencias y fluctuaciones en la salinidad (Kessler, 2006; Portela et al., 2016); como resultado, la región presenta dos periodos hidroclimáticos: la temporada fría/seca de enero a junio, y el periodo cálido/húmedo de julio a diciembre.
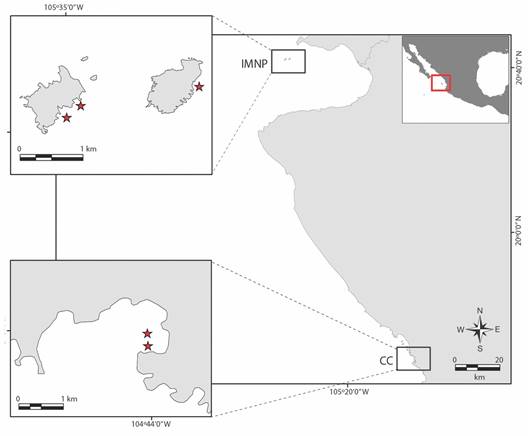
Fig. 1 Área de estudio con localidades y sitios de restauración. A. Parque Nacional Islas Marietas (PNIM), con tres sitios: Cueva del Muerto (CM), Zona de Restauración (ZR) y Plataforma Pavonas (PP). B. Caleta de Cuastecomates (CC) con dos sitios de restauración Cuaste 1 (C1) and Cuaste 2 (C2). / Fig. 1. Study area with localities and restoration sites. A. Islas Marietas National Park (PNIM), with three sites: Cueva del Muerto (CM), Zona de Restauración (ZR) and, Plataforma Pavonas (PP). B. Caleta de Cuastecomates (CC) with two restoration sites Cuaste 1 (C1) y Cuaste 2 (C2).
A pesar de localizarse dentro de la misma región, los sitios de estudio presentan diferentes características locales. El PNIM (Fig. 1A) es un Área Natural Protegida insular que se encuentra a 9 km de la costa (Cupul-Magaña & Rodríguez-Troncoso, 2017) al norte del PCM. El sitio está conformado por dos islas con presencia de sustrato rocoso, fondos arenosos y presencia de coral vivo, con una cobertura de ~17 %, distribuida entre 1-8 m de profundidad (Hernández-Zulueta et al., 2017). Los cambios estacionales regionales están definidos principalmente por la influencia de surgencias estacionales en el periodo frío y ondas internas que provocan fluctuaciones diarias, tanto en la TSM como en la salinidad (Portela et al., 2016). La distinción de parque nacional permite que se lleven a cabo actividades turístico-recreativas controladas (Cupul-Magaña & Rodríguez-Troncoso, 2017). En el PNIM se evaluaron tres sitios en restauración: Cueva del Muerto (CM), Zona de Restauración (ZR) y Plataforma Pavonas (PP).
Por otro lado, la Caleta de Cuastecomates es una localidad costera que se localiza en el límite sur del PCM (Fig. 1B), que alberga una comunidad coralina con una cobertura de coral vivo registrada del 20 % (Hernández-Zulueta et al., 2017), siendo la mayor con relación a sitios adyacentes (Hernández-Zulueta et al., 2021). La CC tiene una profundidad promedio de 8.4 m, con sustrato de roca y arena, y con un aporte estacional de sedimento y nutrientes, provocando una alta turbidez durante la época de lluvias (Rojo-Vázquez et al., 2001). La mayor cobertura de coral se encuentra a una profundidad de 0 a 2.5 m, en donde se llevan a cabo actividades turísticas no controladas como son el buceo libre, pesca artesanal, y uso como zona de anclaje de embarcaciones, por lo que se ha observado altos niveles de fragmentación en colonias adultas. En este sitio se consideraron dos sitios a restaurar: Cuaste 1 (C1) y Cuaste 2 (C2).
Trabajo de campo: En ambas localidades, se colectaron fragmentos de oportunidad de corales del género Pocillopora, de ≥ 3 cm de largo, empleando el método propuesto por Tortolero-Langarica et al. (2014). Cada fragmento fue evaluado visualmente en su condición, con el fin de utilizar solo fragmentos sanos y sin presencia de la esponja bioerosionadora Cliona sp. Los fragmentos obtenidos de cada localidad se fijaron directamente en sustrato natural en huecos o grietas encontrados en la roca o matriz arrecifal, por medio de cintillas de plástico (Edwards, 2010); cabe resaltar que no hubo transporte de fragmentos entre sitios o localidades. Dada la distancia entre localidades, la fijación de los fragmentos se llevó en dos etapas. Durante septiembre 2019, se colocaron un total de 60 fragmentos en los tres sitios dentro del PNIM: Cueva del Muerto (CM, n = 20), Zona de Restauración (ZR, n = 20) y en Plataforma Pavonas (PP, n = 20). En los sitios de CC se colocaron un total de 93 fragmentos entre julio y octubre 2019: Cuaste 1 (C1, n= 60) y Cuaste 2 (C2, n= 33). Cada fragmento se etiquetó y monitoreó individualmente durante un año, con una periodicidad bimestral en el PNIM y trimestral en la CC. En cada muestreo, se registró el tamaño del fragmento desde la base hasta la altura máxima apical (cm), con la ayuda de un vernier de plástico (precisión de 0.05 mm). Finalmente, se determinó la tasa de crecimiento anual (cm·año-1) y la supervivencia (%) para cada sitio. Considerando número final de fragmentos sanos respecto al inicio del experimento, que no presentaron signos de fragmentación, blanqueamiento o sobrecrecimiento de organismos competidores (p. ej. macroalgas y esponjas).
Trabajo de gabinete. Se analizaron las diferencias en el crecimiento entre las localidades de estudio (PNIM vs CC) utilizando una prueba t-student (a = 0.05). Para analizar las diferencias en la tasa de crecimiento entre sitios de ambas localidades (CM, PP, ZR; C1 y C2) se llevó un Análisis de Varianza de una Vía (ANDEVA), y una prueba a posteriori de Holm-Sidak (P = 0.05). La caracterización de las condiciones ambientales incluyó datos mensuales de TSM, PAR y Kd490, mediante el procesado de imágenes satelitales (Aqua-MODIS), con una resolución espacial de 4x4 km y nivel de procesamiento 3 de la plataforma Oceancolor (https://oceancolor.gsfc.nasa.gov/l3/). Para determinar diferencias mensuales de las variables ambientales entre localidades, se utilizó un análisis de varianza no paramétrico de Kruskal-Wallis (P < 0.05). Con el fin de determinar la relación de la tasa de crecimiento con las diferentes variables ambientales, se realizaron pruebas de correlación de Pearson (r), con base en la periodicidad de monitoreo de crecimiento para cada localidad. Todos los análisis estadísticos se llevaron a cabo en el programa SigmaPlot Ver. 11.0. (SPSS Inc., USA).
Resultados
Los fragmentos de oportunidad plantados en el PNIM tuvieron una supervivencia del 61.66 %, con un rango de crecimiento de 2.0 -7.2 cm·año-1. A nivel de sitio, CM presentó una supervivencia 55 %, y una tasa de crecimiento de 4.51 ± 0.49 cm·año-1, mientras que en ZR la supervivencia fue del 60 % con un crecimiento de 3.62 ± 0.41 cm·año-1, y finalmente PP con una supervivencia del 70 % y una tasa de crecimiento de 4.12 ± 0.24 cm·año-1 (Fig. 2). Con respecto a CC, se estimó una supervivencia del 68.08 % (n = 64), con un rango de crecimiento de 1.80-8.00 cm·año-1. El sitio C1 presentó una supervivencia del 56.66 % y tasa de crecimiento de 5.09 ± 0.24 cm·año-1, mientras que en C2 se observó una supervivencia de 87.87 % y un crecimiento de 3.16 ± 0.26 cm·año-1 (Fig. 2). No se evidenció diferencias significativas en la tasa de crecimiento entre localidades (t = 1.169, P = 0.246), pero a nivel de sitios C1 fue el único que mostró diferencias (F = 6.610, P < 0.001), siendo además, en el que se registró la mayor tasa de crecimiento.
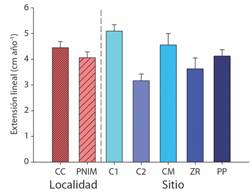
Fig. 2 Tasa de crecimiento anual (promedio ± error estándar) resultante de los fragmentos de oportunidad en la Caleta de Cuastecomates (CC): C1- Cuaste 1, C2- Cuaste 2; y en el Parque Nacional Islas Marietas (PNIM): CM- Cueva del muerto, ZR- Zona de restauración, PP- Plataforma pavonas, localizados en la región del Pacífico Central Mexicano. / Fig. 2. Annual growth rate (mean ± standard error) resulted from fragments of opportunity fragments in Caleta de Cuastecomates (CC): C1- Cuaste 1, C2- Cuaste 2; and in Islas Marietas National Park (PNIM): CM- Cueva del muerto, ZR- Zona de restauración, PP- Plataforma pavonas, located in the Central Mexican Pacific region.
La temperatura media en el PNIM fue de 29.89 ± 0.56 °C, con valores mínimos en marzo de 25.02 ± 0.17°C y máximo de 30.81 ± 0.16 °C registrado en septiembre; mientras que en CC la temperatura media fue de 30.43 ± 0.30 °C, con el mínimo registrado en abril (24.19 ± 0.27 °C) y el máximo en agosto (31.64 ± 0.36 °C; Fig. 3A). A pesar de las diferencias en la fluctuación anual de la TSM entre localidades (5.7 °C en el PNIM y 7.45 °C en CC), estas no fueron estadísticamente significativas (H= 0.0696, P= 0.792). Con relación a la radiación fotosintética (PAR), se encontró que el valor promedio en el PNIM fue de 46.15 E·m-2·d-1, con un mínimo de 34.12 ± 1.82 E·m-2·d-1 y valores máximos de 57.24 ± 1.14 E·m-2·d-1, mientras que para CC, la media resultó en 49.29 E·m-2·d-1 en la CC con un rango de 37.19 ± 1.56 E·m-2·d-1 hasta 60.22 ± 0.19 E·m-2·d-1 (Fig. 3B). Sin embargo, los valores de PAR entre localidades no evidenciaron diferencias estadísticas (H = 0.0511, P = 0.821). Con respecto a los valores del coeficiente de atenuación lumínica, en el PNIM se registró un promedio anual de 0.08 ± 8.80x10-3 m-1, con el valor más bajo registrado durante julio (0.06 ± 5.85x10-3 m-1), y el más altos en octubre (0.10 ± 0.03 m-1); mientras que en CC se estimó un intervalo de 0.04 ± 0.06 m-1 hasta 2.71 ± 0.97 m-1, con su valor máximo en abril- mayo (Fig. 3C). No se encontraron diferencias estadísticas entre PNIM y CC (H = 0.249, P = 0.618).
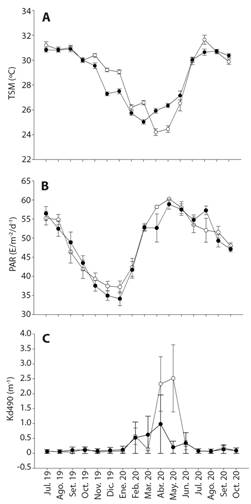
Fig. 3 A. Valor promedio mensual de la temperatura superficial del mar (TSM) (°C ± error estándar); B. radiación fotosintéticamente disponible (PAR) (E/m-2/d-1 ± error estándar) C. el coeficiente de atenuación difusa (Kd490) (m-1 ± error estándar), en el Parque Nacional Islas Marietas (PNIM) (●) y en la Caleta de Cuastecomates (CC) (○), Pacífico Central Mexicano durante el periodo de muestreo. / Fig. 3. A. Monthly mean value of the sea surface temperature (TSM) (°C ± standard error.); B. photosynthetically available radiation (PAR) (E/m-2/d-1 ± standard error); C. diffuse attenuation coefficient (Kd490) (m-1 ± standard error), in Islas Marietas National Park (IMNP) (●) and Caleta de Cuastecomates (CC) (○), Central Mexican Pacific during the sampling period.
Finalmente, se encontró que la tasa de crecimiento del coral en la CC muestra una correlación negativa con la TSM, tanto en C1 (r = -0.773, P = 0.003), como en la C2 (r = -0.760, P = 0.004). En contraste los sitios PNIM no presentaron correlación significativa entre el crecimiento y las variables ambientales consideradas en el presente trabajo (P > 0.05).
Discusión
La restauración efectiva debe de tener como objetivo llevar a la rehabilitación del sistema tanto a una escala local como regional, sin embargo, hasta el momento la mayoría de las iniciativas son programas de restauración enfocados a esfuerzos dentro de una misma localidad a una escala de tiempo menor a dos años (Boström-Einarsson., et al. 2020). Además del financiamiento, uno de los retos para escalar regionalmente es reconocer que los resultados pueden ser diferentes en respuesta a los mecanismos de aclimatación que los corales hayan desarrollado a los estresores tanto naturales como antropogénicos (Ban et al., 2014; Martínez-Castillo et al., 2020). El presente estudio se llevó a cabo en una localidad al norte (PNIM) y otra al sur (CC) de la ecorregión del Pacífico Mexicano Tropical (Spalding et al., 2007), por lo que, aunque están influenciadas por características regionales similares, durante los períodos transicionales presentarían condiciones oceanográficas mixtas del norte y sur del Pacífico Mexicano. Esto sumado a las condiciones locales como son la presencia de surgencias estacionales, ríos, así como el impacto antropogénico, se esperaban evidenciar diferencias en la tasa de crecimiento del coral Pocillopora. El PNIM es un sitio insular que está sometido a una intensa, pero controlada presión, ya que presta al turismo de Puerto Vallarta, servicio de visitación al parque, y con influencia estacional moderada de los aportes continentales al estar separada 9 km de la costa. La CC está a escasos metros de una pequeña comunidad ribereña, con un moderado desarrollo turístico (Gobierno del Estado de Jalisco, 2016), con libre acceso a la pesca, así como visitas no controladas al parche arrecifal, además de recibir descargas domésticas sin tratamiento de manera directa. Nuestros resultados muestran que, hasta el momento, el impacto antropogénico y las condiciones ambientales locales no hay generado un efecto diferencial en la tasa de crecimiento de Pocillopora. Sin embargo, se puede considerar a CC como un sitio con una mayor vulnerabilidad, por lo que durante la restauración, se debe de considerar las medidas mitigatorias para evitar que los corales rebasen su umbral de tolerancia y se vea afectado no solo el crecimiento sino inclusive, la supervivencia.
Aunque las condiciones regionales y locales no provocaron diferencias importantes en el crecimiento de los fragmentos, se debe considerar la influencia de los procesos a macroescala de carácter interanual como es el fenómeno ENOS. Históricamente se ha atribuido a los eventos ENOS El Niño las mortandades de coral más importantes a nivel global (Hughes et al., 2018; Souter et al., 2020). Sin embargo, Pocillopora ha demostrado una resistencia diferente en la región del POT (Romero-Torres et al., 2020), y contrario a lo esperado el crecimiento ha sido positivo ante la exposición a anomalías térmicas (Jiménez & Cortés, 2003; Tortolero-Langarica et al., 2017). De manera particular el PCM registró la mortandad coralina más severa en la historia reciente, durante el evento El Niño 1997-98, a partir de la cual se evidenciado una lenta pero constante recuperación (Hernández-Zulueta, et al., 2017; Martínez-Castillo et al., 2022). La resistencia a las anomalías térmicas por parte de Pocillopora, se representa por su alta supervivencia, y por un incremento constante en su tasa de crecimiento en los últimos diez años (Tabla 1), el cual además, fue mayor en comparación con otros sitios a lo largo del Pacífico Oriental Tropical. Esto incrementa su relevancia ya que durante el periodo 2015-2016 se registró el evento el Niño más intenso de las últimas tres décadas, el cual provocó un blanqueamiento moderado, pero sin mortandad en la región (Cruz-García et al., 2020). Esto muestra una alta resistencia a estresores regionales, así como una alta tasa de crecimiento, el cual incrementa la factibilidad de los esfuerzos de rehabilitación en la región, ya que la historia de vida de Pocillopora demuestra una capacidad de resiliencia la cual puede ser asistida utilizando técnicas activas de restauración, con una mayor probabilidad de éxito.
Table 1 Records of annual linear extension of the genus Pocillopora in the Eastern Tropical Pacific.
| País | Sitio | Periodo | Especie | Extensión lineal (cm año-1) | Fuente |
| México | Islas María Cleofas | 2018 - 2019 | Pocillopora spp. | 2.74 ± 0.33 (DE) | 1 |
| Islas Marietas | 2010 - 2011 | P. damicornis | 2.31 (2.14-2.47) | 2 | |
| P. verrucosa | 2.69 (2.50-2.89) | 2 | |||
| P. capitata | 2.31 (2.01-2.69) | 2 | |||
| 2012 - 2013 | P. damicornis | 3.73 (3.21-4.25) | 2 | ||
| P. verrucosa | 3.69 (3.29-4.09) | 2 | |||
| P. capitata | 3.93 (3.35-4.50) | 2 | |||
| Pocillopora spp. | 4.98 ± 1.68 (DE) | 3 | |||
| Pocillopora spp. | 5.29 ± 1.74 (DE) | 3 | |||
| 2013 - 2014 | Pocillopora spp. | 4.47 ± 1.07 (DE) | 4 | ||
| 2019 - 2020 | Pocillopora spp. | 4.05 ± 0.23 (EE) | * | ||
| Cuastecomates | 2019 - 2020 | Pocillopora spp. | 4.45 ± 0.23 (EE) | * | |
| Zacatoso | 2010 - 2013 | P. damicornis | 2.94 ± 0.32 (DE) | 5 | |
| P. verrucosa | 3.42 ± 0.32 (DE) | 5 | |||
| La Entrega | P. capitata | 2.92 ± 0.23 (DE) | 5 | ||
| P. meandrina | 2.02 ± 0.39 (DE) | 5 | |||
| Francia | Isla Clipperton | 1988 - 1993 | Pocillopora spp. | 2.54 ± 0.57 (DE) | 6 |
| Pocillopora spp. | 2.65 ± 0.34 (DE) | 6 | |||
| Costa Rica | San Pedrito | 1996 - 1997 | P. damicornis | 3.80 ± 0.5 (DE) | 7 |
| P. meandrina | 4.46 ± 0.63 (DE) | 7 | |||
| P.elegans | 4.38 ± 0.57 (DE) | 7 | |||
| P. inflata | 3.15 ± 0.8 (DE) | 7 | |||
| Huevos | 1991 - 1992 | P.elegans | 5.21 ± 0.96 (DE) | 7 | |
| 1996 - 1997 | P. damicornis | 6.68 ± 0.87 (DE) | 7 | ||
| Palmitas | 1996 - 1997 | P. eydouxi | 3.08 ± 0.48 (DE) | 7 | |
| P. meandrina | 3.42 ± 0.66 (DE) | 7 | |||
| P.damicornis | 5.31 ± 0.35 (DE) | 7 | |||
| P.elegans | 4.12 ± 0.58 (DE) | 7 | |||
| Isla del Caño | 1985 - 1987 | P. damicornis | 3.46 ± 0.42 (EE) | 8 | |
| P. damicornis | 2.98 ± 0.31 (EE) | 8 | |||
| P.elegans | 3.48 ± 0.34 (EE) | 8 | |||
| P.elegans | 3.17 ± 0.27 (EE) | 8 | |||
| Bahía Culebra | 2020 | Pocillopora spp. | 4.49 ± 1.19 (DE) | 9 | |
| 5.35 ± 1.48 (DE) | |||||
| 3.25 ± 2.22 (DE) | |||||
| Panamá | Golfo de Chiriquí | 1989 -1990 | P. damicornis | 3.32 ± 0.15 (DE) | 9 |
| 2003 - 2004 | P. damicornis | 2.82 (2.44-3.12) | 10 | ||
| 2005 - 2006 | P. damicornis | 2.75 (2.40-3.30) | 10 | ||
| Golfo de Panamá | 1971 - 1974 | P. damicornis | 3.08 (2.55-3.61) | 11 | |
| 1972 - 1974 | P. damicornis | 3.86 (3.39-4.33) | 11 | ||
| Ecuador | Onslow, Galápagos | 1975 | P. damicornis | 2.24 (DE) | 12 |
Dado el origen diferente de los datos, se presentan el valor promedio, recorrido, desviación estándar (DE) o error estándar (E.E.). / Given the different origin of the data, the average value, range, standard deviation (SD) or standard error (S.E.) are presented. Referencias / References: (*) Presente estudio / present study; (1) Tortolero-Langarica et al., (2019); (2) Tortolero-Langarica et al., (2017); (3) Tortolero-Langarica et al., (2014); (4) Tortolero-Langarica et al., (2020); (5) Medellín-Maldonado et al., (2016); (6) Glynn et al. (1996); (7) Jiménez & Cortés, (2003); (8) Guzmán & Cortés, (1989); (9) Combillet et al., (2022); (10) Eakin, (1996); (11) Manzello, (2010); (12) Glynn, (1977).
Uno de los eventos regionales que no se ha considerado como un factor determinante en el crecimiento de Pocillopora, pero que influenció la región durante el periodo de estudio son los eventos meteorológicos como tormentas y huracanes. El impacto de estos eventos puede ser diferente a nivel regional y local, ya que está en función tanto de su trayectoria como de sus posibles cambios en fuerza (NOAA, 2022). Y si bien pueden ser considerados como estresores negativos o destructivos, dependiendo de su intensidad contribuyen a la circulación y mezcla en la columna de agua evitando la permanencia de sedimentos, alta concentración de nutrientes inorgánicos, así como la disminución de temperatura durante los eventos de estrés térmico (Heron, 2008). Sin embargo, por su morfología, los corales corales ramificados como el género Pocillopora serán los mayormente afectados, incrementando su fragmentación, pero no siempre disminuyendo su cobertura (Lirman, 2001; López-Pérez & Hernández-Ballesteros, 2004). Como resultado, las tormentas pueden contribuir positivamente al principal modo de reproducción asexual en la región (Chávez-Romo et al., 2013); sin embargo, al ser un daño mecánico, la fragmentación puede provocar una disminución de hasta el 50 % en la tasa de crecimiento (Tortolero-Langarica et al., 2014). El sitio CC fue impactado por los huracanes Lorena (Categoría II) y Narda (Categoría I), durante el mismo año, lo cual pudo haber provocado la posible pérdida de fragmentos (~20 %), pero no afectó negativamente la tasa de crecimiento. Por lo que la pérdida de la estabilidad del sustrato y el desprendimiento de los fragmentos por la energía mecánica del oleaje constituye un factor puntual importante en la supervivencia de los fragmentos, más no en su crecimiento individual.
Otro factor relevante que debe de ser considerado dentro del éxito de un programa de restauración es la actividad humana del sitio. Se ha reportado que la actividad turística no controlada puede no solo llevar a la fragmentación, sino a la pérdida de cobertura de coral vivo (Cupul-Magaña & Rodríguez-Troncoso, 2017; De et al., 2020), y a la presencia y propagación de enfermedades coralinas (Lamb et al., 2014). Ambos sitios presentan diferentes presiones por uso turístico, en el caso del PNIM al ser un ANP cuenta con un programa de manejo, así como con la implementación de la capacidad de carga (Cupul-Magaña & Rodríguez-Troncoso, 2017), lo cual contribuye a disminuir el impacto del turismo, mientras que en CC el turismo no está controlado, lo que lleva a que las embarcaciones anclen en los sitios coralinos, además del impacto por las aletas.
Las iniciativas y programas de restauración se han incrementado de manera exponencial en los últimos años (Boström-Einarsson et al., 2020) y de aquí la relevancia de contar con información del área a restaurar que permita establecer un programa que sea exitoso tanto a corto como a largo plazo. Los resultados del presente estudio demuestran que los corales del género Pocillopora tienen un crecimiento similar en localidades insulares y costeros, en respuesta a las condiciones ambientales regionales; y que la mortandad o daño a las colonias de coral se atribuye al efecto puntual de eventos meteorológicos, los cuales, si bien son eventos regionales, su impacto a los arrecifes de coral es siempre diferenciado y se manifiesta a escala local. Sumado a esto, Pocillopora ha demostrado ser resiliente ante eventos ENOS y por lo tanto, lo cual es parte de su capacidad adaptiva natural e incrementa su potencial de rehabilitación asistida. Los resultados muestran la factibilidad y escalabilidad de un programa de restauración en las comunidades coralinas del PCM.
Finalmente, si bien los resultados no muestran diferencias en el crecimiento a pesar de la diferencia en el impacto antropogénico, se deben de considerar medidas de manejo participativo que contribuyan a que la restauración activa tenga un resultado positivo en la cobertura de coral vivo y de sus organismos asociados, para que, a largo plazo, se pueda reducir la vulnerabilidad de los parches arrecifales cercanos a la costa con una fuerte influencia antrópica.
Declaración de ética: los autores declaran que todos están de acuerdo con esta publicación y que han hecho aportes que justifican su autoría; que no hay conflicto de interés de ningún tipo; y que han cumplido con todos los requisitos y procedimientos éticos y legales pertinentes. Todas las fuentes de financiamiento se detallan plena y claramente en la sección de agradecimientos. El respectivo documento legal firmado se encuentra en los archivos de la revista.

