











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
La familia Laridae reúne a gaviotas y gaviotines que habitan diversos ecosistemas, principalmente humedales como estuarios y zonas de la costa (Balachandran, 2012; Blanco, 1999). Debido a la naturaleza migratoria de muchos láridos, sus patrones de variación espaciotemporal a nivel global se ven afectados por factores como el cambio climático e, incluso, variaciones de los ciclos ligados a El Niño - Oscilación Sur (ENOS). Es importante destacar que las especies estenotermas son especialmente vulnerables a esas variaciones climáticas (Orgeret et al., 2022).
Los ecosistemas de humedales se encuentran amenazados a nivel global debido al cambio climático y a factores antropogénicos, lo que resulta en una disminución más intensa en las abundancias de las especies de zonas tropicales y ecuatoriales (Amano et al., 2020; Saino et al., 2011) y afecta particularmente a aquellas especies de aves con hábitos ligados al agua y durante el periplo migratorio de las aves migratorias (Podestá & Barona, 2021; Volkov et al., 2016). Otro aspecto relevante es la segregación de las aves en los ecosistemas de humedales, que puede estar influenciada por diversos factores como la heterogeneidad del hábitat, la estructura biológica de la especie, las características de la vegetación, los aspectos químicos del agua, así como los hábitos y dieta de las especies (Green, 1998; Monda & Ratti, 1988; Pulido et al., 2021).
En este sentido, los mapas de contorno con isolíneas de parámetros y/o índices biológicos son herramientas importantes para estudiar la distribución espaciotemporal de las especies, como en estudios sobre la abundancia de especies de aves (Castillo-Palacios et al., 2014), sobre el índice de Shannon-Weaver para comunidades de peces (Rico, 2000), sobre el porcentaje de cobertura vegetal (Del Castillo Ruiz, 2016), entre otros. El análisis de estos aspectos permite identificar patrones de distribución poblacional de las especies (Arana et al., 2022; Del Castillo Ruiz, 2016), así como, apreciar su dinámica a lo largo del tiempo. Por ejemplo, Cotillo et al. (2018) estudiaron la distribución espacial de las aves playeras limícolas en el humedal costero Poza de La Arenilla, donde se detectó la presencia y ausencia de estas especies, así como la riqueza y abundancia de forma estacional, lo cual contribuyó al conocimiento base de la estancia de estas aves. Estos resultados son relevantes para reconocer las áreas de uso de estas aves y promover estrategias de conservación en este humedal. Además, es importante destacar que las aves no se distribuyen de manera uniforme en estos ecosistemas (Blanco, 1999). Sin embargo, estos ecosistemas se muestran importantes para el reposo y alimentación para las aves migratorias (Apeño & Aponte, 2022; Naranjo & Mauna, 1996).
En el Perú se ha registrado que únicamente 14 de las 29 especies de láridos utilizan los humedales del Callao (Carazas et al., 2021; Gobierno Regional del Callao, 2021; Plenge, 2023). En el caso del humedal costero Poza de La Arenilla (HCPA), no existen estudios sobre la distribución espaciotemporal de las aves acuáticas de la familia Laridae, las cuales utilizan el humedal como una zona de reposo y descanso (Podestá & Cotillo, 2016). Además, en los últimos años, el humedal ha experimentado una degradación crítica por las actividades antropogénicas (Merizalde Burneo, 2020), las cuales alteran el comportamiento de estas aves. Por lo tanto, el objetivo de este trabajo fue evaluar la variación temporal de la abundancia, riqueza y diversidad de las aves de la familia Laridae en el humedal durante el periodo comprendido entre 2013 y 2018. Asimismo, aportar conocimiento sobre la distribución espacial de estas aves en las diferentes zonas del HCPA en Callao, Perú.
Materiales y métodos
Área de estudio: El área de estudio es el HCPA, ubicado en la zona sur del distrito de la Punta, Provincia Constitucional del Callao, Perú (UTM WGS 84 Zona18S: 264 539.768 - 265 273.988 m E & 8 664 186.604 - 8 665 043.327 m N) (Fig. 1). El humedal costero se encuentra en la costa central del Perú y comprende una superficie de 17.6 ha, se caracteriza por la presencia de dos cuerpos de agua como consecuencia de la existencia de dos Bocanas, las cuales se ubican al extremo este y oeste. Los cuerpos de agua se encuentran separados por una zona de arenilla con extensión dependiente del nivel de la marea. Adicionalmente, el sitio colinda con el ambiente urbano a través de un malecón que permite la observación de aves y otras actividades. Cabe mencionar que la zona urbana y la zona de orilla tienen presencia de flora terrestre reducida. El humedal posee una profundidad máxima de 2 m y posee diferentes especies de fauna y flora (Sánchez Rivas et al., 2014).

Fig. 1 Delimitación de las 11 zonas censadas en el humedal costero Poza de La Arenilla, distrito de la Punta, Provincia Constitucional del Callao, Perú. / Fig. 1. Delimitation of the 11 census zones in the coastal wetland Poza de La Arenilla, district of La Punta, Constitutional Province of Callao, Peru.
El HCPA es uno de los tres humedales más representativos del Callao y el más pequeño. Sin embargo, aporta con registros importantes en esta provincia (Podestá, Cotillo et al., 2021). Debido a su gran biodiversidad reportada de 98 especies de aves (Podestá et al., 2017; Podestá, Gil et al., 2021; Podestá & Cotillo, 2016), este humedal se ha convertido en un hábitat importante para aves acuáticas.
Actualmente, el HCPA cuenta con 11 zonas delimitadas, las cuales fueron diferenciadas según el uso que las aves hacen de este ecosistema costero, así como criterios biofísicos y de composición del humedal (Cotillo et al., 2018). Cabe señalar que previamente se había establecido una división en 14 zonas o biotopos (Troll, 2000), pero en años recientes se han observado cambios en el terreno y la composición biológica del humedal debido a factores climáticos y antrópicos (García, 2006).
Delimitación de zonas: Para la identificación de zonas de este humedal se procedió a replicar las zonas previamente establecidas por Cotillo et al. (2018), las cuales fueron distribuidas de acuerdo con la utilización de las aves en este humedal costero. Estas se distribuyen de la siguiente manera: A: Arenilla, C: Canal, C.R.1: Cantos Rodados R1, E.A.1: Espejo de Agua 1, E.A.2: Espejo de Agua 2, M.C: Muro/Canal, O: Orilla, O.A.1: Orilla Arenilla 1, O.A.2: Orilla Arenilla 2, R.1: Roquedal 1 y R.2: Roquedal 2 (Fig. 1).
Muestreo de campo: Se realizaron muestreos quincenales entre enero de 2013 y diciembre de 2018 durante la mañana (07:00-11:00 h) y la tarde (14:00-16:00 h), utilizando el método del conteo total, teniendo en cuenta que este método es recomendado para aves acuáticas y playeras, y el área de estudio es un hábitat pequeño y abierto, lo que permitió que las especies fueran fácilmente observables (Alegría Mont, 2018; Ministerio del Ambiente, 2015; Torres et al., 2006). Se desarrolló el método de conteo por puntos limitados a distancia, se utilizaron un total de seis puntos separados por un mínimo de 250 m de modo que los diferentes hábitats del humedal fueron cubiertos. El tiempo de evaluación por cada punto fue de 5 min, considerando una distancia máxima de avistamiento de 150 m, tal y como indica la metodología de Ralph et al. (1995). Se registró la riqueza de especies, la abundancia y la distribución. Se utilizaron binoculares (10 × 42 mm) y una cámara fotográfica con lentes de 70-200 mm. El método de conteo por bloque se utilizó en determinados casos cuando las bandadas fueron mayores a los 500 individuos (Bibby et al., 2000). Todas las aves fueron identificadas a nivel de especie (DeGraaf & Rappole, 1995; Plenge, 2023; Schulenberg et al., 2010).
Procesamiento de los datos: Los datos de abundancia, obtenidos de forma quincenal en la mañana y en la tarde, se promediaron para obtener una abundancia promedio mensual para el análisis de biodiversidad y una abundancia promedio estacional para el análisis geoespacial (mapas de isolíneas).
Cálculo de la biodiversidad: Se calculó el índice de diversidad de Shannon-Weaver (H’) (calculado con logaritmos naturales) para evaluar la diversidad mensual para el periodo evaluado. También se analizó la variación temporal de la abundancia y la riqueza de especies, y la variación de la riqueza y la abundancia entre zonas dentro del HCPA.
Elaboración de mapas: Los resultados del muestreo de campo fueron representados a través de mapas de isolíneas de abundancia promedio de cada especie de lárido observado por estación del año (verano, otoño, invierno y primavera) durante el periodo de evaluación (2013-2018). La ubicación de los puntos de muestreo representativos para cada zona se estableció por el centroide geométrico de estas. La equidistancia entre las isolíneas varía para cada mapa según rango de abundancia observado. Adicionalmente, se agregaron mapas de calor en escala de grises donde el color negro indica abundancia promedio máxima y el color blanco, abundancia promedio mínima.
Análisis estadístico: Se realizó un análisis de correlación de Spearman entre la diversidad medida a través del índice de H’ y la abundancia y riqueza para determinar cuál de estos últimos muestra una mayor asociación respecto del valor final del índice. Para determinar cuál de las especies contribuye más a las diferencias entre años, se realizó una prueba ANOSIM de similitudes y una prueba post-hoc SIMPER, a partir de las abundancias y la composición de especies tanto a escala mensual como anual. Todos los análisis estadísticos fueron realizados con el programa PAST v2.17c (Hammer et al., 2001), tomando en cuenta un nivel de significancia de 0.05.
Resultados
De enero de 2013 a diciembre de 2018 se registraron 12 especies de aves acuáticas pertenecientes a la familia Laridae, de las cuales diez estuvieron presentes en las cuatro estaciones del año, una especie presente en tres estaciones y una especie presente en sólo una estación (Tabla 1). Cinco especies fueron migratorias y siete especies residentes. La estación con mayor abundancia promedio fue el verano con 11 473.3 individuos de las 12 especies registradas, seguido por la primavera con 7 690.5 individuos de 11 especies, mientras que las estaciones de otoño e invierno presentaron menor abundancia promedio con 3 192.2 individuos de 11 especies, y 756.4 individuos de 10 especies, respectivamente (Tabla 2).
Tabla 1 Aves acuáticas de la familia Laridae registradas en el humedal costero Poza de La Arenilla, distrito de la Punta, Provincia Constitucional del Callao, Perú en el periodo 2013-2018. / Table 1. Aquatic birds of the family Laridae recorded in the coastal wetland Poza de La Arenilla, district of La Punta, Constitutional Province of Callao, Peru in the period 2013-2018.
| Especie | Nombre común | Verano | Otoño | Invierno | Primavera |
| Chroicocephalus cirrocephalus | Gaviota de Capucha Gris | x | x | x | x |
| Chroicocephalus serranus | Gaviota Andina | x | x | x | x |
| Larosterna inca | Gaviotín Zarcillo | x | x | x | x |
| Larus belcheri | Gaviota Peruana | x | x | x | x |
| Larus dominicanus | Gaviota Dominicana | x | x | x | x |
| Leucophaeus atricilla | Gaviota Reidora | x | x | x | x |
| Leucophaeus modestus | Gaviota Gris | x | x | x | x |
| Leucophaeus pipixcan | Gaviota de Franklin | x | x | x | x |
| Sterna hirundinacea | Gaviotín Sudamericano | x | - | - | - |
| Thalasseus elegans | Gaviotín Elegante | x | x | x | x |
| Thalasseus maximus | Gaviotín Real | x | x | x | x |
| Thalasseus sandvicensis | Gaviotín de Pata Negra | x | x | - | x |
x: Presencia / Presence.
Tabla 2 Abundancia promedio estacional de aves acuáticas en el humedal costero Poza de La Arenilla, distrito de la Punta, Provincia Constitucional del Callao, Perú en el periodo 2013-2018. / Table 2. Seasonal average abundance of aquatic birds in the coastal wetland Poza de La Arenilla, district of La Punta, Constitutional Province of Callao, Peru in the period 2013-2018.
| Estación / Especies | C. cirrocephalus | C. serranus | L. inca | L. belcheri | L. dominicanus | L. atricilla | L. modestus | L. pipixcan | S. hirundinacea | T. elegans | T. maximus | T. sandvicensis | Total | ||||||
| Arenilla | V | 0.8 | 15.2 | 1.3 | 20.4 | 2 | 0 | 0.1 | 7 217 | 0 | 4.4 | 3 | 0.1 | 7 264.2 | |||||
| O | 53.5 | 36.6 | 1.9 | 38.3 | 28.6 | 0.2 | 3.3 | 1 114.6 | 0 | 2.4 | 0.4 | 0 | 1 279.9 | ||||||
| I | 162.4 | 99.4 | 0 | 33.1 | 50.7 | 0.3 | 0.3 | 0.3 | 0 | 0.7 | 0.03 | 0 | 347 | ||||||
| P | 16.4 | 14.1 | 0 | 35.3 | 172.8 | 0 | 3.1 | 3 345 | 0 | 0.5 | 0.3 | 0.03 | 3 587.5 | ||||||
| Canal | V | 0.2 | 0 | 0 | 0.2 | 0 | 0 | 0 | 147.5 | 0 | 0 | 0 | 0 | 147.9 | |||||
| O | 0 | 0 | 0 | 0.1 | 0 | 0 | 0.3 | 4.2 | 0 | 0 | 0 | 0 | 4.6 | ||||||
| I | 0.7 | 3.3 | 0 | 0.1 | 6 | 0 | 0.03 | 0 | 0 | 0 | 0 | 0 | 10.1 | ||||||
| P | 0.3 | 0 | 6.5 | 0.3 | 0 | 0 | 0 | 2.7 | 0 | 0.03 | 0 | 0 | 9.7 | ||||||
| Canto Rodados R1 | V | 0 | 0 | 0 | 15.8 | 5.7 | 0 | 0.03 | 871 | 0 | 0.1 | 0 | 0 | 892.6 | |||||
| O | 24.2 | 11.2 | 0.3 | 11.7 | 20.2 | 0 | 1.9 | 118.8 | 0 | 0 | 0 | 0 | 188.2 | ||||||
| I | 4.4 | 0.2 | 0 | 33.5 | 54.1 | 0 | 4.1 | 0 | 0 | 0 | 0 | 0 | 96.3 | ||||||
| P | 1.3 | 0 | 0.2 | 13.2 | 59.9 | 0 | 7.6 | 1 153 | 0 | 0 | 0 | 0 | 1 235.2 | ||||||
| Espejo de Agua 1 | V | 0.5 | 0 | 0 | 3.1 | 0 | 0 | 0 | 57.1 | 0 | 1.8 | 0 | 0 | 62.4 | |||||
| O | 0 | 0 | 1.4 | 2.5 | 4.1 | 0 | 4.3 | 13.1 | 0 | 1.9 | 0 | 0 | 27.2 | ||||||
| I | 0.7 | 0.2 | 0 | 1.3 | 2.1 | 0 | 0.6 | 0 | 0 | 0 | 0 | 0 | 4.8 | ||||||
| P | 3.2 | 0.3 | 0.03 | 5.5 | 0.8 | 0 | 0.3 | 69.7 | 0 | 0.03 | 0 | 0 | 79.9 | ||||||
| Espejo de Agua 2 | V | 0.4 | 0 | 0 | 4.7 | 0.6 | 0 | 1.5 | 192.4 | 0.03 | 0.3 | 0 | 0 | 199.9 | |||||
| O | 12.6 | 3.6 | 5.2 | 1 | 5.3 | 2.1 | 6.4 | 14 | 0 | 2.1 | 0 | 0 | 52.3 | ||||||
| I | 2.1 | 1.4 | 6.4 | 10.6 | 28.4 | 0 | 0.03 | 0.1 | 0 | 0.6 | 0 | 0 | 49.5 | ||||||
| P | 3.3 | 0 | 0.5 | 2.6 | 2.4 | 0 | 0 | 109.7 | 0 | 0.2 | 0 | 0 | 118.6 | ||||||
| Muro/Canal | V | 0 | 0 | 0 | 1.6 | 0 | 0 | 0 | 1.8 | 0 | 0 | 0 | 0 | 3.4 | |||||
| O | 13.2 | 0 | 0 | 3.0 | 9.5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 25.7 | ||||||
| I | 0 | 0 | 0 | 3.1 | 16.3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 19.4 | ||||||
| P | 0 | 0 | 0 | 1.0 | 6.1 | 0 | 0 | 26.8 | 0 | 0 | 0 | 0 | 33.9 | ||||||
| Orilla | V | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |||||
| O | 0 | 0 | 0 | 0.1 | 0 | 0 | 0.9 | 61.8 | 0 | 0 | 0 | 0 | 62.8 | ||||||
| I | 0 | 0 | 0 | 0.9 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.9 | ||||||
| P | 0 | 0 | 0 | 0.4 | 0.7 | 0 | 0 | 645.2 | 0 | 0 | 0 | 0 | 646.2 | ||||||
| Orilla Arenilla 1 | V | 6.8 | 0.1 | 0 | 17.1 | 1.9 | 46.4 | 0 | 917.7 | 0 | 8.3 | 6.3 | 0.2 | 1 004.7 | |||||
| O | 28.4 | 2.1 | 0.5 | 19.8 | 13.6 | 0.03 | 1.2 | 621.4 | 0 | 1.2 | 0.03 | 0.4 | 688.8 | ||||||
| I | 2.4 | 8.3 | 0 | 9.2 | 30.7 | 0 | 0 | 0.1 | 0 | 0 | 0 | 0 | 50.7 | ||||||
| P | 8.3 | 1.2 | 0 | 8.6 | 9.4 | 0.03 | 0.1 | 657 | 0 | 1 | 0.1 | 0 | 685.7 | ||||||
| Orilla Arenilla 2 | V | 2.8 | 2 | 1.1 | 21.8 | 8.6 | 0 | 0 | 1 804.7 | 0.03 | 3.2 | 0.8 | 0 | 1 845.1 | |||||
| O | 50.5 | 12.5 | 0 | 4.2 | 9.6 | 0.7 | 0.6 | 779.0 | 0 | 1.9 | 1 | 0 | 860.1 | ||||||
| I | 29.6 | 62.2 | 0 | 18.2 | 46.6 | 0 | 6 | 0.2 | 0 | 0 | 0 | 0 | 162.8 | ||||||
| P | 22.3 | 5.6 | 0 | 4.8 | 12.1 | 0 | 4 | 1 138.8 | 0 | 0.2 | 0 | 0 | 1 187.8 | ||||||
| Roquedal 1 | V | 0 | 0 | 0.1 | 1 | 0 | 0 | 0 | 36.8 | 0 | 0 | 0 | 0 | 37.9 | |||||
| O | 0 | 0 | 0 | 1 | 1.1 | 0 | 0 | 0 | 0 | 0.5 | 0.1 | 0 | 2.6 | ||||||
| I | 0.4 | 0 | 0 | 8.6 | 2.3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 11.3 | ||||||
| P | 0.7 | 0 | 1.6 | 0.7 | 4.8 | 0 | 0 | 97.5 | 0 | 0 | 0 | 0 | 105.4 | ||||||
| Roquedal 2 | V | 1.2 | 0 | 0 | 0.9 | 0 | 0 | 0 | 7.6 | 0 | 5.0 | 0.6 | 0 | 15.2 | |||||
| O | 0 | 0.03 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.03 | ||||||
| I | 0.1 | 0 | 0 | 1.1 | 2.4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3.5 | ||||||
| P | 0 | 0 | 0 | 0.03 | 0.1 | 0 | 0 | 0 | 0 | 0.4 | 0 | 0 | 0.5 | ||||||
V: verano, O: otoño, I: invierno y P: primavera / V: summer, O: autumn, I: winter and P: spring.
La especie Leucophaeus pipixcan es la de mayor abundancia promedio (dominancia) en 10 de las 11 zonas estudiadas con una abundancia total promedio de 21 226.5 individuos, individuos, su presencia se destaca principalmente en las estaciones de verano y primavera, aunque en las zonas de Arenilla, Orilla Arenilla 2 y Orilla Arenilla 1 también destaca en otoño. La especie con menor dominancia fue Sterna hirundinacea, la cual presentó una abundancia promedio de 0.06 individuos en el periodo de estudio, y se observó solo un individuo en verano en las zonas Espejo de Agua 2 y Orilla Arenilla 2.
Las especies más distribuidas en el humedal son L. pipixcan, Larus dominicanus y Larus belcheri, ya que fueron reportadas en las 11 zonas estudiadas. Por otro lado, las especies de menor distribución son las especies Thalasseus sandvicensis que solo fue reportada en las zonas de Arenilla y Orilla Arenilla 1, y S. hirundinacea reportada en Espejo de Agua 2 y Orilla Arenilla 2.
La zona más importante fue la Arenilla, en donde se registró una abundancia total promedio de 12 478.5 individuos pertenecientes a 11 especies, las estaciones de verano y primavera representaron en conjunto un 87.0 % de la abundancia promedio registrada para la zona; seguido de la zona Orilla Arenilla 2 con 4 055.8 individuos pertenecientes a 10 especies, las estaciones de verano y primavera representaron un 74.8 % de la abundancia promedio registrada para la zona. Las zonas con menor reporte de avistamiento fueron el Roquedal 2 y Muro/Canal con 19.2 y 82.4 individuos, respectivamente (Fig. 2). Los mapas de isolíneas de abundancia promedio por estación del año para cada especie registrada en el HCPA se muestran en las Fig. 3 y Fig. 4.
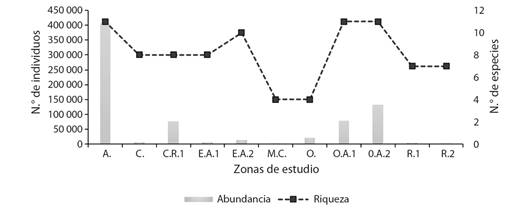
Fig. 2 Abundancia y riqueza de aves acuáticas de la familia Laridae en el humedal costero Poza de La Arenilla, distrito de la Punta, Provincia Constitucional del Callao, Perú. / Fig. 2. Abundance and richness of aquatic birds of the Laridae family in the Poza de La Arenilla coastal wetland, district of La Punta, Constitutional Province of Callao, Peru.
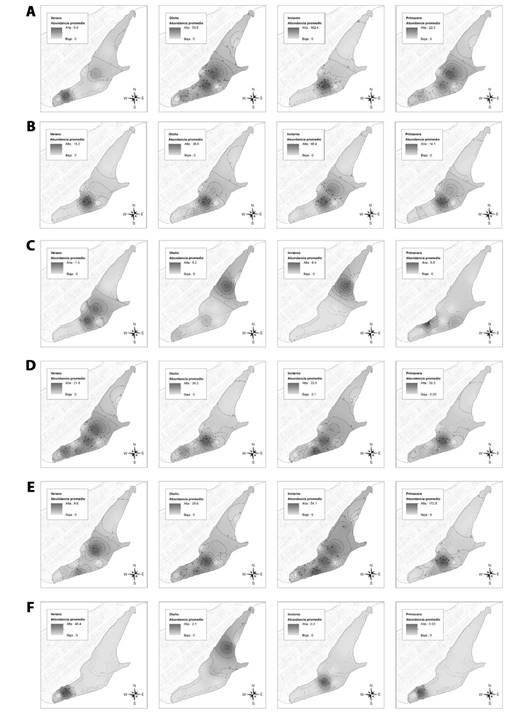
Fig. 3 Mapas de isolíneas de abundancia promedio en las diferentes estaciones del año (2013-2018) en el humedal costero Poza de La Arenilla, distrito de la Punta, Provincia Constitucional del Callao, Perú. A. Chroicocephalus cirrocephalus; B. Chroicocephalus serranus; C. Larosterna inca; D. Larus belcheri; E. Larus dominicanus; F. Leucophaeus atricilla. / Fig. 3. Maps of average abundance isolines in the different seasons of the year (2013-2018) in the coastal wetland Poza de La Arenilla, district of La Punta, Constitutional Province of Callao, Peru. A. Chroicocephalus cirrocephalus; B. Chroicocephalus serranus; C. Larosterna inca; D. Larus belcheri; E., Larus dominicanus; F. Leucophaeus atricilla.
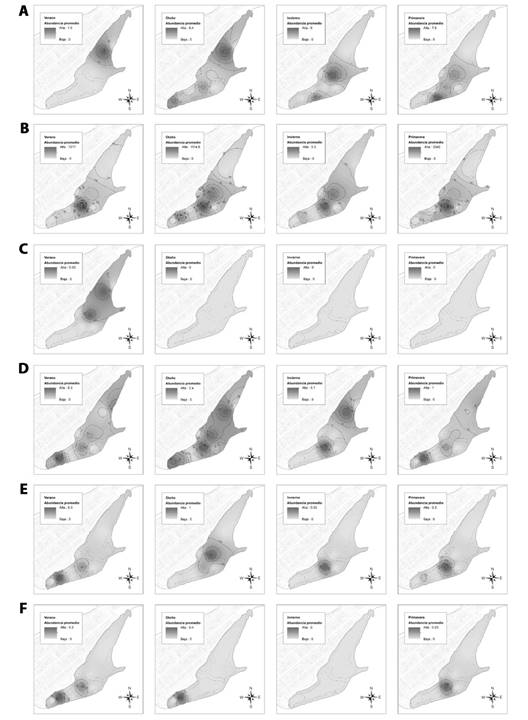
Fig. 4 Mapas de isolíneas de abundancia en las diferentes estaciones del año (2013-2018) en el humedal costero Poza de La Arenilla, distrito de la Punta, Provincia Constitucional del Callao, Perú. A. Leucophaeus modestus; B. Leucophaeus pipixcan; C. Sterna hirundinacea; D. Thalasseus elegans; E. Thalasseus maximus; F. Thalasseus sandvicensis. / Fig. 4. Isoline maps of abundance in the different seasons of the year (2013-2018) in the coastal wetland Poza de La Arenilla, district of La Punta, Constitutional Province of Callao, Peru. A. Leucophaeus modestus; B. Leucophaeus pipixcan; C. Sterna hirundinacea; D. Thalasseus elegans; E. Thalasseus maximus; F. Thalasseus sandvicensis.
Se ha observado un incremento gradual en la abundancia promedio de las aves de la familia Laridae en el periodo 2013-2018 y simultáneamente una tendencia hacia el incremento de la riqueza de aves de la familia Laridae para los meses de primavera y verano entre 2016 y 2018, más no así para la variación global de la riqueza (Fig. 5). De manera similar, en la Fig. 6 se muestra la variación mensual del índice de diversidad de H’ durante el periodo 2013-2018 y se observa un patrón de variación estacional similar de un año al siguiente ya que los láridos son aves migratorias, aunque también se observa una tendencia a la disminución del índice de diversidad a medida que transcurre el tiempo.
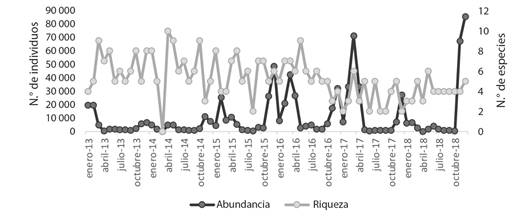
Fig. 5 Variación mensual de la abundancia y riqueza de aves acuáticas de la familia Laridae en el humedal costero Poza de La Arenilla, distrito de la Punta, Provincia Constitucional del Callao, Perú en el periodo 2013-2018. / Fig. 5. Monthly variation in the abundance and richness of aquatic birds of the Laridae family in the Poza de La Arenilla coastal wetland, district of La Punta, Constitutional Province of Callao, Peru in the period 2013-2018.
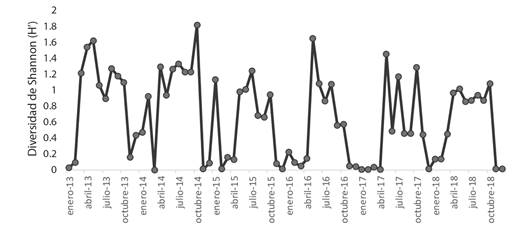
Fig. 6 Variación mensual del índice de diversidad de Shannon-Weaver para las aves acuáticas de la familia Laridae en el humedal costero Poza de La Arenilla, distrito de la Punta, Provincia Constitucional del Callao, Perú en el periodo 2013-2018. / Fig. 6. Monthly variation of the Shannon-Weaver diversity index for waterfowl of the Laridae family in the Poza de La Arenilla coastal wetland, district of La Punta, Constitutional Province of Callao, Peru in the period 2013-2018.
La diversidad medida con el índice de Shannon-Weaver guarda una correlación más fuerte con las abundancias (ρ = -0.69, P < 0.05) que con la riqueza (ρ = 0.52, P < 0.05). Debido a que el índice de diversidad de Shannon-Weaver tiende a incrementarse mientras mayor es la riqueza y la equidad, la correlación inversa hallada entre la diversidad y la abundancia se debe a que, en ensambles comunitarios más equitativos, las abundancias totales tienden a ser más bajas que en ensambles con unas pocas especies claramente dominantes en términos numéricos.
La especie que contribuyó más a las diferencias entre años (2013-2018) fue L. pipixcan, con 69.8 % de contribución según el análisis SIMPER. Esta es la especie de lárido más abundante durante primavera, verano y otoño, habiéndose incrementado su número particularmente en dichas estaciones entre 2016 y 2018.
Discusión
Se registraron 12 especies de láridos en el HCPA durante el periodo 2013-2018: cinco especies migratorias y siete residentes. Es conocida la dominancia de L. pipixcan en las estaciones de verano y primavera en sitios costeros (Bayly, 2015; González et al., 2011; Podestá & Cotillo, 2016; Pulido, 2018), además de visitar lugares de parada y reposo a lo largo de su periplo migratorio (Delgado et al., 2019; Ramírez & Aponte, 2018). No obstante, es importante destacar que existen factores que determinan la distribución de estas aves, en su mayoría gaviotas, dentro de un ecosistema, en este caso humedales costeros donde se concentran importantes especies de aves migratorias (Gauthier et al., 2005). Por ejemplo, González et al. (2011) determinan que el aumento del nivel del agua, la disponibilidad de alimento y otros factores son importantes en la variación estacional de diferentes especies migratorias. A su vez, Yapura et al. (2022) señalan que en ciudades se identifican cambios estacionales en comunidades de aves que podrían estar relacionados a la migración y disponibilidad de alimento en ciertas épocas del año. Estos cambios tendrían relación con la hidrología en humedales costeros, especialmente el funcionamiento biológico y químico (Flores-Verdugo et al., 2007; Keddy, 2010).
Se observó un uso marcado de los hábitats del HCPA por parte de especies dominantes de láridos. En particular, se destaca la presencia en otoño de L. pipixcan en las zonas de Arenilla, Orilla Arenilla 2 y Orilla Arenilla 1. La dominancia de L. pipixcan en las 11 zonas descritas, como se constata en las isolíneas de abundancia promedio que se distribuyen ampliamente por el humedal costero y presentan los valores más altos entre todas las especies, coincide con lo observado por Chung-Velásquez & López-Manrique (2021) en los Pantanos de Villa, un humedal ubicado en la costa sur de Lima, Perú, donde esta especie cubre la totalidad de las zonas de dicho refugio de vida silvestre. Otro punto notable, es la baja abundancia de S. hirundinacia en comparación con otras especies, especialmente del grupo de gaviotines. Con un único registro, se trata de una especie “singleton” (identificación de un solo individuo de una especie en el muestreo) (Podestá & Cotillo, 2016). Esta situación no generaría un impacto significativo en la abundancia promedio de especies en la estación de verano, lo cual no es inusual a este humedal costero, considerando la previa presencia de especies de escolopácidos migratorios con esta misma denominación, como es el caso de Calidris himantopus, que tampoco generó cambios significativos en la abundancia y diversidad de aves playeras en este humedal (Podestá et al., 2017). También Limosa fadea fue identificada como un “singleton” en un estudio realizado en Pantanos de Villa (García-Olaechea et al., 2018).
Entre las especies que utilizan con mayor frecuencia este humedal costero y presentan mayor abundancia promedio en invierno, se encuentra L. dominicanus, especie residente, generalista y oportunista que se distribuye en diferentes tipos de hábitats (Mudge & Ferns, 1982; Pierotti & Annett, 1990). La alta abundancia de L. dominicanus no es un hecho aislado, ya que estudios realizados en otros ecosistemas marinos también han reportado su destacada presencia. Por ejemplo, Delgado et al. (2019) encontraron que la gaviota dominicana fue una de las aves costeras más abundantes encontradas en las islas Desertores, comuna de Chaitén en la Patagonia Chilena. Otro estudio llevado a cabo en las Islas Tenglo y Maillen, también en Chile, demostró que L. dominicanus era una de las especies con mayor abundancia en estas islas, superando incluso a especies de gaviotas que presentaban alta abundancia en otras zonas costeras, tales como L. pipixcan (Gallardo & Rau, 2019). Incluso en otras regiones relevantes con presencia de aves acuáticas durante todos los meses del año, como la laguna de Cahuil, en Chile Central, L. dominicanus se encontró sobrepasando las decenas de individuos (Mella-Romero, 2021). Sin embargo, es importante señalar que L. dominicanus podría encontrarse en diferentes ecosistemas con características ambientales particulares, incluyendo aquellos que han sido significativamente impactados (Silva et al., 2000). En el HCPA, estos cambios ocurren dentro del propio humedal costero, el cual posee diferentes zonas y muestra una distribución particular de las especies acuáticas que se encuentran en él. Es en este contexto que especies como L. belcheri y L. dominicanus, especies oportunistas y generalistas (Silva et al., 2000), se distribuyen en las 11 zonas identificadas en la Poza de la Arenilla, abarcando la totalidad de estas con sus diferencias y similitudes (Cotillo et al., 2018).
Un aspecto relevante es que tanto la riqueza como la abundancia promedio de láridos son mayores en tres de las 11 zonas del humedal para todo el periodo evaluado: La Arenilla, Orilla Arenilla 2 y Orilla Arenilla 1. Adicionalmente, la zona Espejo de agua 2 también está incluida entre las zonas con mayor riqueza de láridos, principalmente debido a su mayor extensión longitudinal en relación con la Orilla Arenilla 1 (Cotillo et al., 2018). Los láridos utilizan principalmente estos hábitats intermareales de baja profundidad como zonas de alimentación y descanso (Burger & Gochfeld, 2019). Estudios previos han mostrado que los láridos pueden verse negativamente afectados cuando el nivel del agua se incrementa (Wu et al., 2021). En la Orilla Arenilla 2, la abundancia promedio de láridos supera a la encontrada en la Orilla Arenilla 1, estando esta última bañada por el agua proveniente del Espejo de agua 1 cuyo flujo de agua es mayor que el del Espejo de Agua 2, y que constituye un hábitat más adecuado para aves de hábitos limícolas (Cotillo et al., 2018). Por otro lado, las concentraciones de oxígeno disuelto cerca a la Orilla Arenilla 2 son mayores que en las inmediaciones de la Orilla Arenilla 1 (Sánchez Rivas et al., 2014), lo cual podría favorecer un incremento de la diversidad acuática que sirve de alimento a las aves del humedal (Hsu et al., 2011).
En cuanto a la variación de abundancias promedio y riqueza de Laridae, se ha observado una tendencia a la disminución en la riqueza de láridos durante todo el periodo evaluado, así como una tendencia al incremento en las abundancias totales, siendo notorios estos incrementos entre las primaveras y los veranos de un año al siguiente, lo cual se traduce también en una disminución de la diversidad medida a través del índice de H’. Estos patrones de variación de riqueza, abundancia y diversidad podrían estar ligados a los cambios en temperatura superficial del mar (TSM) registrados para esta zona durante el mismo periodo (Podestá & Barona, 2021). Debido a que L. pipixcan es la especie de lárido que alcanza mayores abundancias, particularmente durante verano, primavera y otoño, la diversidad medida a través del índice de Shannon-Weaver tiende a disminuir. Esta disminución de la diversidad se corresponde también con el hecho de que las abundancias promedio de L. pipixcan han mostrado un incremento sostenido en los veranos de varios de los años estudiados, influyendo en mayor medida sobre los valores de diversidad en comparación a la influencia que tiene la riqueza de especies. Debido a que esta especie es omnívora y generalista (Burger, 1988; Calvino-Cancela, 2011), la disminución de la abundancia promedio de otras especies de aves más especializadas debido al incremento de la TSM, es posible que haya dejado el camino libre a L. pipixcan en relación con la ocupación del hábitat y su dominancia numérica (Washburn et al., 2013), por ello es que los resultados muestran que esta ha sido la especie que más ha contribuido en las diferencias de abundancias promedio entre los años evaluados.
Se concluye que el humedal costero Poza de La Arenilla (HCPA) en Callao, Perú, desempeña un papel fundamental como hábitat para la avifauna acuática de la región. Se ha registrado la presencia de cinco especies migratorias y siete residentes de la familia Laridae, las cuales muestran preferencias espaciales particulares en diferentes zonas del humedal. A pesar de que se observa una disminución en la riqueza global de las especies en el tiempo (2013-2018), la abundancia promedio de aves Laridae aumenta.
Se destaca la importancia de conservar el humedal como hábitat clave para las aves acuáticas de la familia Laridae. Se requieren investigaciones adicionales para comprender mejor los factores que influyen en la distribución espaciotemporal de las estas aves en el humedal, así como evaluar los efectos a largo plazo del cambio climático y las actividades antropogénicas en estas especies.
Declaración de ética: los autores declaran que todos están de acuerdo con esta publicación y que han hecho aportes que justifican su autoría; que no hay conflicto de interés de ningún tipo; y que han cumplido con todos los requisitos y procedimientos éticos y legales pertinentes. Todas las fuentes de financiamiento se detallan plena y claramente en la sección de agradecimientos. El respectivo documento legal firmado se encuentra en los archivos de la revista.

