











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
Las palmas son un elemento florístico dominante en los bosques tropicales, cubriendo diversos ecosistemas, que incluyen las tierras bajas, pantanos, bosques secos y bosques de alta montaña (Balslev, 2011). Dentro de estos bosques encontramos que las palmas también dominan los diferentes estratos del bosque, gracias a las múltiples adaptaciones que observamos en la familia como: el amplio rango de su tamaño y forma de sus hojas, tallos, raíces y frutos. Estas características a su vez han convertido a las palmas en una numerosa fuente de materiales habituales para uso humano en los trópicos (Balslev, 2011).
Uno de los ambientes en el que las palmas han logrado prosperar es el sotobosque de zonas tropicales, a pesar que este entorno presenta fuertes limitantes, especialmente de luz, los cuales pueden causar que las plantas que habitan allí sean sensibles a la sequía y tener deficiencias fotosintéticas. Las palmas no solo han colonizado este ambiente en abundancia (Svenning, 2001), sino también en diversidad y se conocen cerca de 15 géneros de palmas que incluyen algunas especies capaces de tolerar o son habitantes exclusivos del sotobosque (Baker & Dransfield, 2016). Inclusive, hay grupos completos en los que las especies se encuentran restringidas a este ambiente, como lo son los géneros Neotropicales Chamaedorea y Geonoma.
En varios géneros de palmas que habitan el sotobosque se presentan adaptaciones compartidas como la alta supervivencia desde la etapa de plántulas en adelante (Endress et al., 2004), la alta longevidad de las hojas (Ataroff & Schwarzkopff, 1994; Chazdon, 1992; Oyama, 1990), las bajas tasas de crecimiento, tanto en producción de hojas como en elongación del tallo (Kimura & Simbolon, 2002; Rodríguez-Buriticá et al., 2005; Souza et al., 2003), tamaño pequeño en la madurez, que compensa el costo por las bajas tasas de captura de luz (Chazdon, 1986a; Kimura & Simbolon, 2002), tallos y hojas flexibles que soportan daño mecánico, el cual es un evento común en el sotobosque (Chazdon, 1986a), y bajas tasas de saturación fotosintética (Chazdon, 1986b); entre otras.
Las estrategias de crecimiento son una ventana para conocer las adaptaciones al sotobosque que pueden implementar las especies. Según esto, las condiciones limitantes en el sotobosque pueden favorecer dos estrategias de crecimiento. Por un lado, existen plantas de rápido crecimiento, que tienen una mayor inversión en tejidos fotosintéticos y pueden reemplazar rápidamente los daños físicos generados por las dinámicas del dosel (Cooley et al., 2004). Por otro lado, encontramos plantas de lento crecimiento, que destinan sus recursos a la producción de estructuras de defensa para evitar los daños por caída de hojas y ramas (Cooley et al., 2004). Además, estas adaptaciones también favorecen distintas estrategias de selección de nicho, que se han descrito como habitat filtering, en la cual patrones de crecimiento similares se seleccionan; y exclusión competitiva, en la que se favorece la diferenciación de patrones (Andersen et al., 2012). Estas son dos estrategias descritas en palmas de sotobosque y determinan la selección de nicho al coexistir (Andersen et al., 2012).
Las tasas vitales son tratos determinantes en la historia de vida de las especies y sus poblaciones, y están expresadas a través de patrones y rasgos de supervivencia, crecimiento, y reproducción de los individuos (Bellier et al., 2018). A través del entendimiento de las tasas vitales, a nivel de una comunidad, es posible identificar diferentes adaptaciones de cada una de las especies que la conforman como respuesta a un mismo hábitat. Los comportamientos variables de las tasas vitales de las especies son explicados por la evolución de las historias de vida, aspecto que puede brindar herramientas para conocer algunos mecanismos ecológicos de diferenciación de nicho en una comunidad.
A diferencia de los estudios de impacto de la cosecha sobre la dinámica poblacional (Endress et al., 2004; Isaza et al., 2017), son escasos los trabajos que abordan el conocimiento de comunidades de especies de palmas y las estrategias que presentan para vivir en el sotobosque. Es por esto que el objetivo de este estudio fue determinar y comparar las tasas vitales de supervivencia, crecimiento y fecundidad en una comunidad de palmas andinas de sotobosque. A partir de esta información estimamos la longevidad de los individuos. Esta información contribuirá con el conocimiento de las diferentes estrategias de adaptación a las condiciones de sotobosque en una comunidad de palmas. Así como en el desarrollo de planes de manejo acordes a las estrategias de vida de las especies.
Materiales y métodos
Área de estudio: El estudio se llevó a cabo en el Parque Natural Chicaque (PN Chicaque), Cundinamarca, Colombia (4º37’9” N & 74º18’53.5” W). El área tiene una extensión de 300 ha, comprendido en un gradiente altitudinal desde los 2 000 a los 2 700 metros de altitud cubierto por bosques montanos de niebla andinos. La temperatura promedio del parque es de 17 ºC en la zona baja y 12 ºC en la zona alta. La humedad relativa se encuentra entre el 75 % al 86 % con una precipitación de 2 000 mm anuales, presentando una tendencia bimodal. El primer pico de lluvia se observa entre marzo y mayo, y el segundo entre octubre y noviembre (Rivera & Córdoba, 1998). Los suelos se caracterizan por ser de textura franco-arcillosa en la zona alta del parque, y arcillosos, franco-arenosos y pedregosos en el límite inferior (Rivera & Córdoba, 1998). Posee una vegetación característica de bosques secundarios mixtos, con múltiples estratos, uno arbustivo y herbáceo denso, compuesto principalmente por Passiflora sp., Cavendishia sp. y Bocconia sp., y uno arbóreo dominado por Croton sp., Cecropia sp., Ficus sp. y Clusia sp., además de tener una población conservada de Quercus humboldtii (Rivera & Córdoba, 1998).
Especies de estudio: Las especies de palmas Chamaedorea linearis (Ruiz & Pav) Mart., Chamaedorea pinnatifrons (Jacq.) Oerst., Geonoma orbignyana Mart. y Geonoma undata Klotzsch. son propias del sotobosque de bosques montanos húmedos tropicales. En Colombia su distribución es principalmente andina (Galeano & Bernal, 2010). Chamaedorea linearis se distribuye entre 500 y 2 800 m, tiene un tallo solitario que alcanza 10 m de altura, es dioica, forma racimos infrafoliares, y frutos rojos esféricos o elipsoides de 1-2.5 cm de diámetro (Galeano & Bernal, 2010). Chamaedorea pinnatifrons se encuentra desde el nivel del mar hasta 2 500 m, frecuentemente sobre los 1 000 m, tiene un tallo solitario ocasionalmente postrado que alcanza 3 m de altura, es dioica, forma racimos infrafoliares y frutos naranjas de 1 cm de diámetro (Galeano & Bernal, 2010). Geonoma orbignyana se distribuye entre 900 y 2 900 m; tiene un tallo solitario ocasionalmente postrado que alcanza 7 m de altura y es raramente cespitosa, es monoica, sus inflorescencias son interfoliares y forma frutos negros y esféricos u ovoides de 0.7 cm de diámetro (Galeano & Bernal, 2010). Geonoma undata se encuentra entre 1 200 y 3 150 m; tiene un tallo solitario, es ocasionalmente cespitosa, alcanza 10 m de altura, es monoica, tiene racimos infrafoliares y forma frutos ovoides o elipsoides levemente quillados color negro de 0.6 cm de diámetro (Galeano & Bernal, 2010).
Datos demográficos: Los datos demográficos de las cuatro especies fueron recolectados de los individuos ubicados dentro de siete parcelas permanentes de 100 m2 (0.07 ha en total), distanciadas por más de 250 m entre ellas, desde los 2 130 m a los 2 370 m de altitud, establecidas empleando los parámetros de: 1) presencia de, por lo menos, una especie de la comunidad, 2) ambiente representativo de crecimiento de las especies (Taherdoost, 2016). Chamaedorea linearis, C. pinnatifrons y Geonoma undata se encontraron en las siete parcelas establecidas, abarcando el gradiente altitudinal de las parcelas. A diferencia de estas, Geonoma orbignyana se encontró en sólo tres parcelas de las siete establecidas, desde 2 130 m a 2 250 m de altitud.
Fueron censados 345, 480, 138 y 482 individuos de C. linearis, C. pinnatifrons, G. orbignyana y G. undata respectivamente. Los individuos fueron marcados permanentemente, identificados, medidos y mapeados para su ubicación posterior. Para construir la estructura poblacional, los individuos se clasificaron como plántulas, juveniles, subadultos y adultos. Se consideraron como plántulas los individuos con hojas bífidas, juveniles a los individuos con hojas pinnadas, pero sin presencia de tallo aéreo, subadultos a los individuos con tallo sin evidencia de inicio de reproducción (con cicatrices de inflorescencias o racimos secos colgando debajo del individuo), y adultos a los individuos con estructuras reproductivas o marcas en el tallo de estructuras reproductivas (Bernal & Galeano, 2013). Los resultados de la densidad poblacional se extrapolaron a 0.1 ha, para facilitar la comparación de estos datos.
Determinación de las tasas vitales: Los datos para estimar las tasas vitales fueron recolectados durante un año en cuatro censos trimestrales desde abril-junio de 2018 a abril-junio de 2019. Esto con el fin de tener un monitoreo adecuado de las poblaciones, además de registrar los ritmos de fecundidad como la aparición de nuevas plántulas y la producción de racimos. Sin embargo, para construir los modelos de las tasas vitales, se usaron los datos del primer y último censo.
Se estimaron las tasas vitales de supervivencia, crecimiento y fecundidad. El tamaño de los individuos fue estimado como el número de pinnas en la hoja expandida más joven, la longitud foliar (únicamente para Geonoma orbignyana) y la altura del tallo (en subadultos y adultos), porque estas variables permiten inferir su tamaño y las tasas vitales (Bernal & Galeano, 2013).
Para estimar la supervivencia se registraron los individuos vivos en el censo final y su tamaño. Para estimar el crecimiento se registraron: 1) la producción de hojas nuevas, 2) el incremento en el número de pinnas al lado derecho de la lámina de la hoja más joven, la longitud de la lámina de la hoja más joven (únicamente para Geonoma orbignyana), 3) la altura del tallo, y 4) el número de anillos cada 30 cm y 10 cm para G. orbignyana, para estimar el promedio de la longitud de los entrenudos en los intervalos de altura mencionados. Se obtuvo el promedio de los entrenudos a una altura determinada y se multiplicó por la producción foliar para calcular el incremento del tallo aéreo anual. Los parámetros de incremento número de pinnas e incremento de altura del tallo se usaron para estimar la edad de los individuos.
Se obtuvo un bajo número de individuos con tallo dentro de las parcelas en las cuatro especies. Por esto, en septiembre de 2019 se registraron fuera de las parcelas 88 individuos con tallo de Chamaedorea linearis, 78 de C. pinnatifrons, 68 de Geonoma orbignyana y 40 de G. undata. En estos individuos se registró el número de pinnas, la longitud de la lámina solo para Geonoma orbignyana, la altura del tallo y el número de anillos (como en el censo del 2018).
Para estimar la fecundidad, se registró si los individuos eran reproductivos, y el número de racimos y frutos que produjeron. En las mismas parcelas, se registró la germinación y el reclutamiento de nuevas plántulas, entendido como las plántulas que, emergen entre el primer y segundo censo y aquellas que sobreviven después de un año, respectivamente (Bernal & Galeano, 2013). El número de racimos y frutos se registraron durante 18 meses, desde abril-junio de 2018 hasta octubre-diciembre de 2019. Se realizó una subcategorización de los individuos adultos para determinar diferencias en el número de frutos y racimos. La categoría Adultos1 corresponde a adultos inferiores a 2, 1.5, 1 y 3.5 m en C. linearis, C. pinnatifrons, G. orbignyana y G. undata, respectivamente. La categoría Adultos 2 corresponde a individuos superiores a las alturas anteriores. El registro de la germinación y el reclutamiento de nuevas plántulas se inició a partir del segundo censo y se registró durante 15 meses, desde julio-septiembre de 2018 a octubre-diciembre de 2019. Para el reclutamiento, se contaron las plántulas nuevas y se registró su supervivencia y producción foliar después de un año. La proporción de individuos en fructificación se obtuvo como la sumatoria de los racimos con fruto sobre el total de los adultos.
Análisis de datos: Para establecer la relación entre las tasas vitales y el tamaño de los individuos (número de pinnas, longitud foliar y altura del tallo), se parametrizaron modelos lineares generalizados simples. Se ajustaron modelos logísticos para analizar las variables binomiales (probabilidad de supervivencia, de formación de tallo y de inicio de reproducción) y discretas (incremento en el número de pinnas) (Zar, 1999). Se realizaron transformaciones logarítmicas y de raíz cuadrada en las variables cuando se presentó sobre dispersión (Apéndice 1). Las funciones de las regresiones y modelos fueron seleccionadas a partir del valor P, el criterio informativo Akaike (AIC) y el coeficiente de determinación (R2) (Akaike, 1981; Zar, 1999). Empleamos estadística descriptiva para determinar la estructura poblacional, número y producción de hojas en categorías de crecimiento, supervivencia de plántulas germinadas, número de racimos formados e individuos germinados en cada censo. Se realizaron pruebas de normalidad Shapiro y homocedasticidad de los datos recopilados para medir las tasas vitales. Se empleó la prueba no paramétrica Kruskal-Wallis y la prueba post-Hoc Mann-Whitney para identificar las diferencias en el tamaño, número y producción de hojas entre categorías de crecimiento (Zar, 1999). Se implementó la prueba t-Student y Mann-Whitney-Wilcoxon (Zar, 1999) para observar diferencias en las tasas vitales entre los sexos de C. linearis y C. pinnatifrons. El análisis estadístico se realizó en el software R con un nivel de significancia del 95 % (R Core Team, 2020), con los paquetes Rmisc 1.5 (Hope, 2013), dunn.test 1.3.5 (Dinno, 2017) y rsq 2.2 (Zhang, 2021).
Resultados
Densidad y estructura poblacional: Se registraron un total de 597, 687, 453 y 690 individuos en 0.1 ha de C. linearis, C. pinnatifrons, G. orbignyana y G. undata, respectivamente. Estos valores muestran que la densidad es variable entre especies, factor atribuido a la distribución agregada en sitios con características diferentes para cada especie. Por ejemplo, para G. orbignyana, la especie con la menor densidad, se observaron individuos densamente agregados en un solo sector de la reserva. Este sitio presentó condiciones de alta humedad, suelos drenados y baja formación de claros en el dosel. Mientras que G. undata, la especie con mayor densidad, se distribuyó en sitios con una alta heterogeneidad del suelo y condiciones de mayor intensidad lumínica. La densidad de los individuos de las cuatro especies en las áreas muestreadas fue de 2 427 individuos / 0.1 ha.
La estructura poblacional en las cuatro especies tuvo una forma de “J” invertida, con un alto número de individuos en las categorías de plántulas, que disminuyeron en las categorías sucesivas (Fig. 1). Comparado con las otras especies, G. undata tuvo una proporción mayor de subadultos respecto a los adultos, probablemente por un incremento en la permanencia de los individuos como subadultos a causa de una lenta elongación del tallo durante esta etapa, como veremos más adelante (Fig. 1).
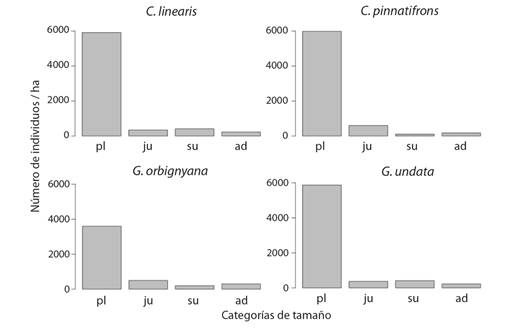
Fig. 1 Estructura poblacional de la comunidad de palmas en el PN Chicaque. Las letras en las categorías indican plántulas (pl), juveniles (ju), subadultos (su) y adultos (ad). / Fig. 1. Population structure of palm community in NP Chicaque. Letters indicate seedlings (pl), juveniles (ju), subadults (su) and adults (ad).
Supervivencia: Las plántulas fueron la etapa más vulnerable de Chamaedorea linearis, Geonoma orbignyana y G. undata con porcentajes de supervivencia de 56 %, 70 % y 62 %, respectivamente. La mortalidad de las plántulas estuvo relacionada principalmente a perturbaciones en el bosque, como caída de hojas sobre los individuos y plena exposición lumínica por disturbios en el dosel. En las siguientes categorías de crecimiento la supervivencia fue mayor (83 a 90 %) y las causas de mortalidad fueron el marchitamiento de los individuos por depredación y por perturbación humana. La supervivencia de C. pinnatifrons fue similar a lo largo del ciclo de vida, teniendo un porcentaje de supervivencia del 72 % en plántulas y de 73 % en las siguientes categorías. Por esto, el modelo aplicado indicó que la supervivencia de C. pinnatifrons no estuvo determinada por el tamaño de los individuos, contrario a lo encontrado en las otras tres especies (Fig. 2).
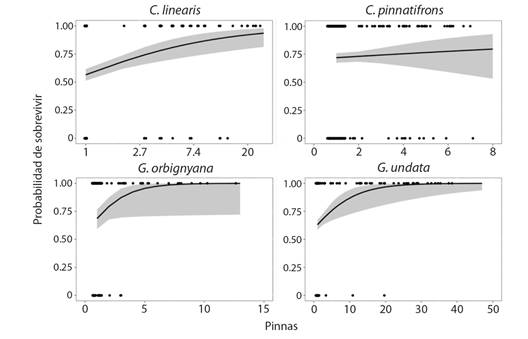
Fig. 2 Probabilidad de supervivencia de las especies de palmas en el PN Chicaque. Los puntos corresponden a los datos observados, la línea corresponde a la tendencia del modelo logístico (P > 0.05), el área sombreada corresponde a los intervalos de confianza del modelo. Los datos de pinnas de C. linearis fueron transformados a logaritmo y se presentaron los ejes de la gráfica en escala natural para facilitar su interpretación. / Fig. 2. Survival probability of palms species in NP Chicaque. Points are observed data, the line is the logistic model tendency (P> 0.05), shadow areas are the confidence intervals of the model. Pinnae of C. linearis were transformed to logarithm and the axis in the graph were represented in natural scale to ease of interpretation.
Crecimiento: Hubo un incremento de tamaño de los individuos sin tallo en la población, evaluado a través del número de pinnas, el cual incrementó desde el censo inicial al final para todas las especies. En promedio se produjeron 2.8 ± 1.6 pinnas×año-1 en Chamaedorea linearis, 1.6 ± 1.0 pinnas × año-1 en C. pinnatifrons, 2.2 ± 2.1 pinnas × año-1 en Geonoma orbignyana y 2.9 ± 2.6 pinnas x año-1 para G. undata. Sin embargo, este comportamiento es variable y las cuatro especies presentaron retrocesos del número de pinnas en algunos individuos, que pasaron de la etapa juvenil (hoja dividida) a plántula (hoja bífida) (Fig. 3). Al mismo tiempo, se observó que pocos individuos (< 10) pueden tener un rápido incremento de pinnas en las etapas tempranas de crecimiento. En algunos casos registramos individuos que en el censo inicial tenían una (1) pinna y produjeron hasta 9 pinnas luego de un año (Fig. 3). Sin embargo, este crecimiento tiene un límite y a medida en que los individuos crecen, la velocidad del incremento en el número de pinnas se reduce, hasta alcanzar su máximo, que es aproximadamente 22, 7, 12 y 41 pinnas en C. linearis, C. pinnatifrons G. orbignyana y G. undata, respectivamente (Fig. 3).
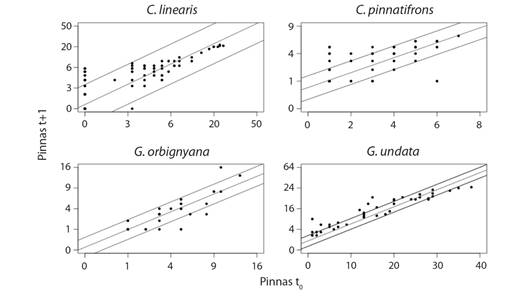
Fig. 3 Crecimiento de tamaño expresado como el incremento en el número de pinnas durante un año para las especies de palmas en el PN Chicaque. Los puntos corresponden a los datos observados, la línea central corresponde a la tendencia obtenida del modelo lineal (P < 0.05) y las líneas externas corresponden al intervalo de confianza. Se transformaron los datos de Pinnas t+1 y Pinnas t0 a logaritmo en C. linearis; en C. pinnatifrons, G. orbignyana y G. undata la variable Pinnas t+1 se transformaron a raíz cuadrada. La variable Pinnas t0 se transformó a raíz cuadrada en G. orbignyana. Los ejes transformados a logaritmo y raíz cuadrada se presentaron en escala natural para facilitar su interpretación. / Fig. 3. Size growth expressed as increment in number of pinnae over one year for each palm species in NP Chicaque. Points are observed data; the central line is the tendency of the model (P < 0.05) and the outer lines are the confidence interval. Data was transformed to logarithm and square root but axes was represented in to natural scale to ease of interpretation. Pinnae t+1 and Pinnae t0 transformed to logarithm in C. linearis. Pinnae t+1 transformed to square root in C. pinnatifrons, G. orbignyana and G. undata. Pinnae t0 transformed to root square in G. orbignyana.
En Chamaedorea, el incremento de la elongación del tallo fue menor a medida que los individuos ganaban altura (Fig. 4). Por su parte, G. orbignyana no presentó diferencias en el incremento de la elongación con respecto a su altura inicial (p > 0.05). Mientras que, en G. undata la elongación del tallo fue mayor al incrementar la altura y fue más baja a menor altura, especialmente por debajo de 1 m altura (Fig. 4). Por otro lado, en promedio, la elongación del tallo presentó un incremento de 0.2 ± 0.1 m ×año-1 de su altura inicial en C. linearis, 0.1 ± 0.08 m ×año-1 en C. pinnatifrons, 0.03 ± 0.01 m ×año-1 en G. orbignyana y 0.07 ± 0.03 m×año-1 en G. undata. De acuerdo a estas cifras se observa que las tasas de crecimiento del tallo para los individuos de Geonoma son mucho más bajas que las de los de Chamaedorea, aunque en todo caso hay una alta variabilidad del incremento de la elongación del tallo en las cuatro especies (Fig. 4).
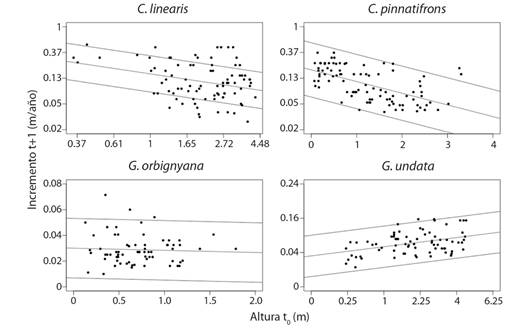
Fig. 4 Crecimiento de tamaño expresado como incremento de la altura del tallo durante un año en las especies de palmas en el PN Chicaque. Los puntos corresponden a los datos observados, la línea central corresponde a la tendencia obtenida del modelo lineal (P < 0.05) y las líneas externas corresponden al intervalo de confianza. Los ejes de los datos transformados a logaritmo y raíz cuadrada se presentaron en escala natural para facilitar su interpretación. Incremento t+1 y Altura t0 transformado a logaritmo en C. linearis. Incremento t+1 transformado a logaritmo en C. pinnatifrons. Incremento t+1 y Altura t0 transformado a raíz cuadrada en G. undata. / Fig. 4. Size growth expressed as increment in height over one year for palms species in NP Chicaque. Points are observed data; central line is the tendency of the model (P < 0.05) and outer lines are the confidence interval. Axis of data transformed to logarithm and square root were presented in natural scale to ease of interpretation. Increment t+1 and Height t0 transformed to logarithm in C. linearis. Increment t+1 transformed to logarithm in C. pinnatifrons. Increment t+1 and Height t0 transformed to square root in G. undata.
Las especies Chamaedorea linearis, C. pinnatifrons y Geonoma undata inician la formación del tallo, que marca la transición de la categoría de juveniles a subadultos, cuando tienen aproximadamente 17, 5, y 20 pinnas en la hoja más joven, respectivamente (Fig. 5). En G. orbignyana, la formación del tallo inicia cuando sus hojas alcanzan una longitud cercana a los 0.4 m (Fig. 5).
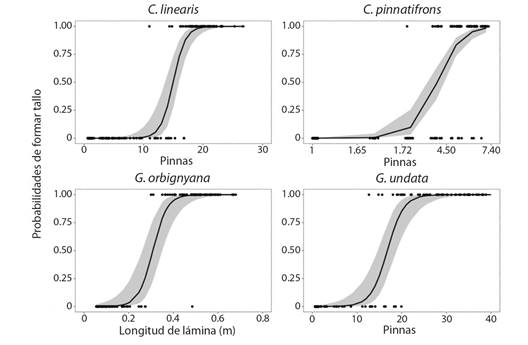
Fig. 5 Probabilidad de formar tallo aéreo respecto al tamaño inicial de los individuos de la comunidad de palmas en el PN Chicaque. Los puntos corresponden a los datos observados, la línea corresponde a la tendencia del modelo logístico (P < 0.05), el área sombreada corresponde a los intervalos de confianza. Los datos de pinnas de C. pinnatifrons fueron transformados a logaritmo y en la gráfica se presentaron en escala natural para facilitar su interpretación. / Fig. 5. Probability of stem formation regarding the initial size of individual of the palm community in PN Chicaque. Points are observed data, the line is the logistic model tendency (P > 0.05), shadowed areas are the confidence intervals. Pinnae of C. pinnatifrons were transformed to logarithm and were presented in natural scale in the graph to ease of interpretation.
El número de hojas, su tamaño y la producción foliar, incrementan con la categoría de tamaño de los individuos en las cuatro especies. En el caso del número de hojas en la corona, los adultos de C. pinnatifrons, G. orbingyana y G. undata tuvieron mayor cantidad (Tabla 1). La excepción fue la especie C. linearis, pues son los subdultos quienes tienen, en promedio, el mayor número de hojas en la corona (Tabla 1). En las especies del género Chamaedorea el incremento del número de hojas totales desde juveniles hasta subadultos es gradual y son similares entre especies, contrario a lo que se presenta en las especies del género Geonoma que prácticamente se duplica de una categoría a otra, produciendo así coronas más grandes y con más hojas (Tabla 1).
Tabla 1 Promedio de hojas, tamaño foliar y hojas producidas para las poblaciones de palmas. / Table 1. Average leaves, leaf size and leaves produced of populations of palms.
| Especie | Número de hojas | Tamaño foliar1 | Hojas producidas |
| C. linearis | |||
| pl | 2.06 ± 0.96a | 1.00 ± 0a | 1.09 ± 0.95a |
| ju | 2.90 ± 1.08a | 6.73 ± 3.94b | 1.69 ± 0.71b |
| su | 3.50 ± 1.85bc | 19.75 ± 2.81c | 3.33 ± 0.57c |
| ad | 3.61 ± 0.98c | 19.78 ± 2.28c | 3.00 ± 1.58bc |
| C. pinnatifrons | |||
| pl | 2.04 ± 1.05a | 1.00 ± 0a | 1.23 ± 0.93a |
| ju | 2.84 ± 1.13b | 3.53 ± 1.24b | 1.35 ± 0.72a |
| su | 2.93 ± 1.12b | 5.64 ± 0.88c | 2.42 ± 0.78b |
| ad | 3.29 ± 1.10b | 5.46 ± 1.00c | 1.58 ± 0.79ab |
| G. orbignyana | |||
| pl | 2.29 ± 1.11a | 0.11 ± 0.04a | 1.98 ± 0.87a |
| ju | 2.50 ± 1.91a | 0.21 ± 0.12b | 2.2 ± 1.31ab |
| su | 8.00 ± 1.87b | 0.44 ± 0.09c | 3.16 ± 1.47ab |
| ad | 11.06 ± 3.15b | 0.52 ± 0.08c | 3.25 ± 1.03b |
| G. undata | |||
| pl | 2.56 ± 1.13a | 1.00 ± 0a | 1.36 ± 1.12a |
| ju | 4.39 ± 2.25b | 8.64 ± 6.03b | 2.14 ± 1.26b |
| su | 13.25 ± 2.66c | 27.45 ± 7.35c | 3.21 ± 1.74b |
| ad | 13.91 ± 2.34c | 37.06 ± 3.80d | 2 ± 1.41ab |
1El tamaño foliar en C. linearis, C. pinnatifrons y G. undata corresponde al número de pinnas. En G. orbignyana corresponde a la longitud de la lámina (m). pl: plántulas; ju: juveniles; su: subadultos; ad: adultos. Las pruebas K-W fueron significativas en las cuatro especies (P < 0.05). Los subíndices en cada valor corresponden a la prueba post-hoc U de Mann-Whitney. / 1Leaf size for C. linearis, C. pinnatifrons and G. undata is the number of pinnae. For G. orbignyana, is the leaf longitude (m). pl: seedlings; ju: juveniles; su: subadults; ad: adults. Tests K-W were significant for the four species (P < 0.05). Subscripts in each value are the post-hoc test Mann-Whitney U.
El tamaño de las hojas se basó en el número de pinnas en C. linearis, C. pinnatifrons, y G. undata, y por la longitud de la lámina de la hoja más joven en G. orbignyana. Estas variables incrementan con la categoría de tamaño de los individuos (Tabla 1). En Chamaedorea, las hojas con mayor número de pinnas se encuentran en los individuos subadultos, aunque no son significativamente diferentes del número de pinnas en los adultos (Tabla 1). Mientras que, en las especies del género Geonoma las hojas de mayor tamaño se encuentran en los adultos.
Igualmente, en las cuatro especies, la producción foliar incrementó con el aumento en el tamaño de los individuos. La diferencia en la producción foliar entre categorías fue significativa. Esta tendencia fue clara en Geonoma orbignyana, en la que los individuos adultos tuvieron la mayor producción foliar en la población (Tabla 1). Mientras que en C. linearis, C. pinnatifrons y G. undata fueron los subadultos quienes tuvieron la mayor producción foliar (Tabla 1), una etapa en la que las especies de palmas exhiben un rápido crecimiento.
A pesar de que la mayor producción foliar ocurre en individuos subadultos y adultos en las cuatro especies, algunos pocos individuos (2.6 % del total), excepcionales, en estado de plántulas de C linearis, C. pinnatifrons y G. undata alcanzaron una alta producción foliar, llegando a producir aproximadamente 5 hojas por año. A partir del número de hojas y la producción de hojas calculamos la longevidad promedio de las hojas. Las hojas de las plántulas y juveniles de las cuatro especies tuvieron una duración cercana a los 2 años. En la etapa de subadulta la relación cambia y se reduce la longevidad de las hojas en Chamaedorea a 1 año, mientras que en Geonoma esta osciló entre los 1.5 a 2 años. Por último, en la etapa de adultos en Geonoma se obtuvo una duración foliar de 2 a 3 años y en Chamaedorea su longevidad fue de casi la mitad con 1.5 a 2 años. De manera en el género Geonoma hay una retención de hojas prolongada que le permite formar coronas más numerosas y grandes.
Fecundidad: La probabilidad de reproducirse dada una altura determinada, fue variable en cada especie. Por ejemplo, en C. linearis y C. pinnatifrons la producción de racimos inicia cuando los individuos alcanzan una altura aproximada de 1.5 m, pero puede iniciar desde los 0.2 m para la primera especie. Por otro lado, los individuos de C. pinnatifrons pueden permanecer como no reproductivos incluso hasta alcanzar los 2.7 m (Fig. 6). Mientras que en G. orbignyana se alcanza a 1 m, pero se observaron individuos maduros a menor altura (Fig. 6). En G. undata la formación de racimos inicia cuando los individuos alcanzan una altura de 3 m aproximadamente (Fig. 6). No se encontró una relación entre el número de racimos, el número de frutos y la altura de los individuos (Tabla 2; Apéndice 1), es decir que estas variables no cambiaron con el tamaño de los individuos.
Tabla 2 Número de racimos y frutos dentro de la categoría adulta. / Table 2. Number of racemes and fruits on the adult category.
| C. linearis | C. pinnatifrons | G. orbignyana | G. undata | ||||||
| SubCategoría | Racimos | Frutos | Racimos | Frutos | Racimos | Frutos | Racimos | Frutos | |
| Adultos 1 | 1.5 ± 0.8 | 320 ± 216 | 1.2 ± 0.4 | 182 ± 113 | 2.2 ± 1.1 | 104 ± 101 | 1.7 ± 1 | 859 ± 503 | |
| Adultos 2 | 1.4 ± 0.6 | 233 ± 75.9 | 1.1 ± 0.3 | 120 ± 51 | 2.3 ± 1 | 98 ± 64.7 | 1.9 ± 0.9 | 1 565 ± 963 | |
La prueba Mann-Whitney U para probar diferencias entre las categorías adultas no fue significativa en ningún caso (p > 0.05). Adultos1 individuos inferiores a 2, 1.5, 1 y 3.5 m en C. linearis, C. pinnatifrons, G. orbignyana y G. undata respectivamente. Adultos2 individuos con alturas mayores a los Adultos1. / Test Mann-Whithey U to test differences between adult categories were not significant in any case (p > 0.05). Adults1 individuals under 2, 1.5, 1 and 3.5 m in C. linearis, C. pinnatifrons, G. orbignyana y G. undata respectively. Adults2 individuals with heights bigger than in Adults1.
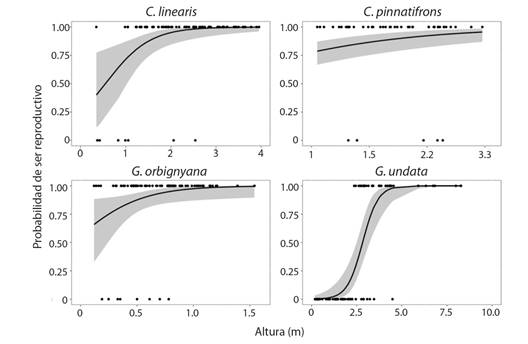
Fig. 6 Probabilidad de ser reproductivo con respecto a la altura inicial del tallo de los individuos en el PN Chicaque. Los puntos corresponden a los datos observados, la línea corresponde a la tendencia del modelo logístico (P < 0.05), el área sombreada corresponde a los intervalos de confianza del modelo. Los datos de altura de C. pinnatifrons fueron transformados a logaritmo, los ejes se presentaron a escala natural en la gráfica para facilitar su interpretación. / Fig. 6. Probability of being reproductive regarding the initial height of individuals in the community of Chicaque NP. Points are observed data, the line is the logistic model tendency (P > 0.05), shadowed areas are the confidence interval. The height of C. pinnatifrons was transformed to logarithmic scale but the axe was presented in natural scale in the graph to ease of interpretation.
Para las especies dioicas (Chamaedorea) la relación entre sexos fue de 1:1 (H:M). Sólo se encontraron 1 y 2 individuos adultos de C. linearis y C. pinnatifrons respectivamente, que no fueron categorizados por su sexo, debido a la ausencia de estructuras reproductivas en buen estado que permitieran inferirlo. No se encontraron diferencias significativas en el número de hojas, producción foliar y número de pinnas de los dos sexos para las especies, excepto para el número de pinnas en C. pinnatifrons, que fue mayor en hembras (5.7 ± 0.7) que en machos (5.1 ± 0.9).
Los individuos de Chamaedorea presentaron una menor, pero continua producción de racimos. No hubo diferencias en la producción de racimos entre machos y hembras en las dos especies de Chamaedorea. Pero, si observamos diferencias entre el tiempo de desarrollo de las estructuras reproductivas para las dos especies. El tiempo de desarrollo desde la apertura de la bráctea que recubre la inflorescencia hasta tener frutos maduros en C. linearis es de 9 a 15 meses y 6 a 9 meses en C. pinnatifrons. Por otro lado, los abortos de racimos completos fueron de 38 % en C. linearis y de 40 % en C. pinnatifrons. Las Geonomas produjeron un mayor número de racimos que las Chamaedoreas. El tiempo de desarrollo de las estructuras reproductivas, que va desde la apertura de la bráctea que recubre la inflorescencia hasta tener frutos maduros, es de 9 a 12 meses en G. orbignyana y 6 a 12 meses en G. undata. Los abortos de racimos fueron el 51 % en G. orbignyana y el 38 % en G. undata.
De julio a septiembre de 2018 se encontraron 146 plántulas germinadas de las cuatro especies. Chamaedorea linearis produjo 20, C. pinnatifrons 86, Geonoma orbignyana 17 y G. undata 23 (Fig. 7). Después de un año, la probabilidad de supervivencia de las plántulas de G. undata fue la más alta con el 0.65, y en cambio C. linearis tuvo la más baja con 0.35 (Fig. 7). Luego de producir 2 hojas, el porcentaje de supervivencia fue de 75 % en C. linearis, 45 % en C. pinnatifrons, 87 % en G. orbignyana y 81 % en G. undata. La tasa de producción foliar anual de las plántulas recién emergidas fue de 1.9 ± 0.9 en C. linearis, 1.2 ± 1 en C. pinnatifrons, 1.8 ± 0.7 en G. orbignyana y 2 ± 0.8 en G. undata.
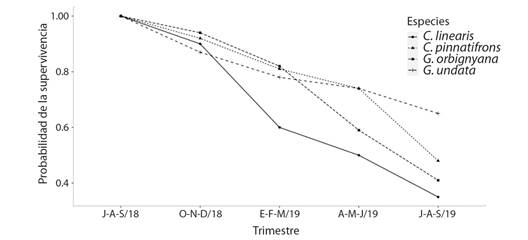
Fig. 7 Supervivencia de individuos reclutados en cada especie durante el tiempo del estudio. / Fig. 7. Survival of recruited individuals for each species during the study time.
Estimación de edades: A partir de la medición de las tasas vitales pudimos estimar las edades de cada especie, las cuales fueron diferentes para cada una (Tabla 3). En Chamaedorea, los individuos crecen rápidamente y pasan la mayor parte de su vida como adultos, iniciando a reproducirse a los 14 y 26 años en C. linearis y C. pinnatifrons, respectivamente, con ciclos de vida de 37 a 62 años. En Geonoma ocurre lo contrario, las palmas pasan la mayor parte de su vida como no reproductivos, iniciando la floración a los 76 años aproximadamente, con longevidades que superan los 100 años de edad.
Tabla 3 Tiempo de duración en cada categoría de tamaño por especie (años). / Table 3. Duration of each size category by specie (years).
| Categorías | C. linearis | C. pinnatifrons | G. orbignyana | G. undata |
| Plántulas | 2.3 | 9 | 23.6 | 21.6 |
| Juveniles | 9.4 | 14.4 | 42 | 22.5 |
| Subadultos | 2.4 | 2.4 | 11.3 | 32.8 |
| Adultos | 23 | 35.7 | 40.8 | 43.8 |
| Duración total | 37.1 | 61.5 | 117.7 | 120.7 |
Discusión
Las características demográficas encontradas en la comunidad de palmas, y que parecen ser propias de las condiciones limitantes del sotobosque, son: una alta supervivencia de las plántulas, bajas tasas de crecimiento en individuos sin tallo, baja producción de hojas con tamaño foliar reducido, pocas hojas totales y en consecuencia una alta longevidad de las hojas e inicio de la reproducción con un tallo aéreo de baja altura. Las especies que recogen la mayoría de estas características son G. orbignyana y C. pinnatifrons, por lo que podemos decir que las estrategias de crecimiento de estas las restringe a las condiciones del sotobosque. Por el contrario, C. linearis y G. undata parecen tener estrategias mixtas de crecimiento, pues sus hojas y coronas tienen un mayor tamaño, las tasas de crecimiento del tallo cambian con la altura de los individuos, y la reproducción inicia a una altura considerable además de encontrarlas fácilmente en espacios abiertos. A nivel de los géneros en Chamaedorea se tiende a priorizar el crecimiento y la reproducción, observado en sus ciclos de vida cortos y mayor inversión del tiempo como adultos reproductivos. Por otro lado, en el género Geonoma parece ser más importante la permanencia y defensa de los individuos de la vegetación circundante.
Supervivencia: La supervivencia de C. linearis, G. orbignyana y G. undata tuvo un incremento a lo largo del crecimiento, es decir que los individuos más grandes tuvieron más probabilidad de sobrevivir, aunque esta no fue considerable. Mientras que en C. pinnatifrons la supervivencia no cambió con el tamaño. En general, las cuatro poblaciones tuvieron una alta supervivencia en los individuos más pequeños (plántulas y juveniles). En palmas se han registrado altas tasas de supervivencia de individuos una vez superan la etapa de plántula (Isaza et al., 2017; Ninazunta et al., 2016). No obstante, aquí encontramos que la supervivencia de las plántulas reclutadas (al cabo del primer año) fue mayor al 30 % en las cuatro especies. Este se encuentra en el límite inferior considerando que previamente, en condiciones de sotobosque, se ha reportado una supervivencia mayor en esta etapa como respuesta a la baja competencia entre plántulas (Ataroff & Schwarzkopf, 1992). Se ha propuesto que la mortalidad de los individuos más jóvenes ocurre por una marcada escasez de luz en el sotobosque, el ataque de herbívoros y patógenos, el daño físico por la caída de hojas y ramas de los árboles circundantes, y el pisoteo por la fauna (Ataroff & Schwarzkopf, 1992). Por otro lado, la supervivencia de los individuos reclutados incrementa considerablemente luego de producir la segunda hoja, como respuesta posiblemente a la independencia de la semilla y el inicio del crecimiento a partir de recursos propios (Ataroff & Schwarzkopf, 1992). Los patrones de supervivencia en las poblaciones generan una estructura poblacional de tipo “J invertida”, indicando que las especies se encuentran en proceso de regeneración (Bernal & Galeano, 2013).
Crecimiento: El crecimiento fue estimado a partir de cuatro parámetros, tres basados en el aumento de hojas, su tamaño y producción y un cuarto basado en la elongación de los entrenudos del tallo aéreo. De estos parámetros, el cambio en el tamaño de hojas y la elongación del tallo fueron aquellos que permitieron identificar con mayor claridad los ritmos de crecimiento de las cuatro especies, a pesar que todos los parámetros mostraron un incremento progresivo entre las categorías de tamaño, en términos de tamaño inicial vs. tamaño final. En este sentido, se ha observado en otras poblaciones de palmas de sotobosque que las tasas de crecimiento de elongación de tallo, producción de hojas y tamaño de hojas incrementan notablemente cuando inicia la producción de un tallo aéreo (subadultos) posiblemente debido a un mejor acceso a la luz, dado por un aumento en la altura, (Rodríguez-Buriticá et al., 2005). En otras especies de sotobosque como G. macrostachys, G. congesta y C. tepejilote las etapas de juveniles tienen una menor tasa de crecimiento, pues se ha observado que esta etapa es susceptible a la baja iluminación en el sotobosque (Oyama, 1990).
Si bien existen tendencias generales del crecimiento entre las especies, encontramos diferencias en otros aspectos del crecimiento, tanto a nivel de géneros como entre las especies, que pueden explicarse por su arquitectura o sus requerimientos de nicho. Las diferencias entre los géneros se observan en el número de hojas y la elongación del tallo. Para el caso del número de hojas, las cantidades son mayores en las especies de Geonoma que en las de Chamaedorea, en los individuos con tallo. Esta diferencia puede deberse a la arquitectura de las plantas, pues en Geonoma encontramos tallos más robustos y peciolos más fuertes, que permiten sostener coronas más grandes y con más hojas (Chazdon, 1986a; Ninazunta et al., 2016).
El crecimiento del tallo también fue diferente entre los dos géneros. En Chamaedorea, la elongación del entrenudo del tallo se reduce sucesivamente con cada hoja producida cuando inician la reproducción, comportamiento observado en otras especies del género (Souza et al., 2003). Al contrario, en Geonoma la elongación del entrenudo del tallo no cambia sustancialmente cuando inician la reproducción. Estos dos patrones son comunes en palmas de sotobosque y muestran dos estrategias de crecimiento en los individuos con tallo (Souza et al., 2003; Sylvester & Avalos, 2013). En Geonoma, la ausencia de una relación positiva entre el crecimiento y la elongación del entrenudo del tallo puede responder a factores diferentes en las dos especies. Las tasas de saturación fotosintéticas de G. orbignyana son bajas y es posible que se cumplan tempranamente en el ciclo de vida, lo que en consecuencia trae un inicio de la reproducción a baja altura (Rodríguez-Buriticá et al., 2005). En G. undata, el patrón de elongación del tallo observado puede responder a una estrategia de acumulación de biomasa (Sylvester & Avalos, 2013). En esta estrategia se priorizan, inicialmente, coronas más grandes y con más hojas (G. undata fue la especie con más hojas y de mayor tamaño), que longitud del tallo, haciendo una mayor inversión en órganos de captura de energía que puedan mantener los altos costos de formación y mantenimiento del tallo en un ambiente con limitaciones de luz (Sylvester & Avalos, 2013). Esto puede explicar que, en la etapa inicial de formación del tallo, la especie hace un balance energético entre aumentar su área foliar para crecer y reproducirse, e incrementar el tallo para acceder a mejores condiciones de luz (Sylvester & Avalos, 2013).
A nivel de las especies, observamos dos tipos de estrategias. Una en la que los individuos tienen tamaños reducidos en sus estructuras vegetativas y, la otra, en la que alcanzan mayores portes. En el primer grupo encontramos a C. pinnatifrons y G. orbignyana que son palmas que no superan los 7 metros de altura y tienen hojas pequeñas que no cambian mucho de tamaño (Galeano & Bernal, 2010). Estas características son propias de palmas de áreas más sombreadas en el sotobosque, las cuales tienen tasas fotosintéticas y de saturación bajas (Chazdon, 1986a; Hogan, 1988). Así mismo, G. orbignyana y C. pinnatifrons comienzan a reproducirse a baja altura, esto puede indicar que los individuos jóvenes experimentan condiciones lumínicas adecuadas para comenzar su reproducción. El tamaño al cual se alcanza la madurez sexual determina el nivel de sombra que las especies pueden tolerar, y el tamaño de las hojas indican el costo en biomasa que debe ser compensado por las condiciones lumínicas (Chazdon, 1986a). Por el contrario C. linearis y G. undata pueden necesitar una mayor compensación por un incremento en los costos de la biomasa al incrementar su tamaño (Chazdon, 1986a), y que sus estrategias de crecimiento no necesariamente están restringidas al sotobosque, por lo que fue posible observar individuos adultos bajo condiciones de luz directa. Esto hace que C. linearis y G. undata sean palmas con alto potencial para aprovechar las perturbaciones del dosel. Es por esto que las características encontradas en estas poblaciones, pueden ser una evidencia del aprovechamiento de las condiciones lumínicas del sotobosque.
Fecundidad: No encontramos una relación entre la producción de racimos y el tamaño de los individuos en las cuatro especies. Este patrón contradice al encontrado en poblaciones diferentes de G. orbignyana y C. pinnatifrons, en las que se registró que el número de racimos aumenta con el tamaño, posiblemente debido al acceso a mejores condiciones lumínicas (Ataroff & Schwarzkopf, 1992; Rodríguez-Buriticá et al., 2005). Sin embargo, es posible que estas diferencias entre poblaciones se deban a cambios en las dinámicas del dosel y nubosidad entre los bosques. El PN Chicaque, es un bosque nublado tropical, en el que las condiciones lumínicas no presentan una alta fluctuación, y esta condición puede ocasionar diferencias mínimas en la producción de estructuras reproductivas, como se presenta en Astrocaryum mexicanum (Piñero & Sarukhán, 1982). Por otro lado, el tiempo de desarrollo de las estructuras reproductivas hasta tener frutos maduros tiene alta variación entre los individuos y las especies estudiadas, con un tiempo mínimo de 6 a 15 meses. Esta alta variación puede generar que haya una oferta permanente de semillas de las cuatro especies. Es posible que los patrones fenológicos irregulares de las cuatro especies dependan de la estacionalidad local, haciendo que los patrones de fructificación puedan cambiar entre poblaciones.
En cuanto a los patrones de supervivencia de las plántulas germinadas se observan diferencias entre las especies que pueden derivarse de los recursos disponibles de las semillas. Es así como, las semillas de C. linearis, que son más grandes que las de otras tres especies (Galeano & Bernal, 2010), potencialmente pueden hacer uso de los recursos propios sin depender de los factores externos como los picos de precipitación o de radiación. Esto también se sustenta en los ritmos continuos de germinación encontrados en C. linearis, que ocurren entre 60 y 300 días, mientras que en G. undata el tiempo es más restringido, ocurriendo entre 70 y 200 días (Isaza, C. datos sin publicar).
Longevidad de los individuos: Las edades estimadas y el tiempo de inicio de la primera floración fueron diferentes entre géneros. Por un lado, las especies del género Chamaedorea tienen una mayor duración como adultos, etapa que inicia con la reproducción a los 13 años y alcanza edades hasta de 66 años, datos que coinciden con lo reportado para poblaciones de C. pinnatifrons en Venezuela (Ataroff & Schwarzkopf, 1994). En este bosque, las palmas del género Geonoma tienen una mayor duración en la etapa no reproductiva, que se extiende hasta el primer evento de reproducción, aproximadamente a los 81 años. Igualmente, se obtuvo que las Geonomas son plantas longevas, alcanzando edades de hasta 127 años como fue reportado antes para G. orbignyana (Rodríguez-Buriticá et al., 2005). Este trabajo es un primer aporte que demuestra las diferentes estrategias en la inversión de recursos y tiempo de las etapas en el ciclo de vida entre los géneros de la comunidad de palmas de sotobosque.
Estrategias de crecimiento y selección de nicho: Las tasas vitales encontradas nos permiten conocer las estrategias de crecimiento de la comunidad de palmas en un sotobosque, y nos da claves para entender la forma en la que ocurre la selección del nicho al coexistir. Por un lado, los individuos en Chamaedorea priorizan el crecimiento y la reproducción, al pasar la mayor parte de su vida como adultos, además, sus rápidas tasas de crecimiento les permite tener un rápido reemplazo de estructuras dañadas por las dinámicas del sotobosque, lo que vemos reflejado en una menor inversión en tejidos de soporte como coronas grandes o tallos robustos (Cooley et al., 2004). Por otro lado, los individuos en Geonoma priorizan la permanencia de los individuos; observado en los largos ciclos de vida, las bajas tasas de crecimiento y una mayor inversión en estructuras de soporte como los tallos y peciolos robustos (Cooley et al., 2004). Así es como evidenciamos las dos estrategias de las palmas de sotobosque para adaptarse a las condiciones lumínicas limitantes. Además, las diferencias en las tasas vitales abordadas entre los géneros y entre las especies pueden sugerir que, en estas poblaciones, ocurre exclusión competitiva en la selección de nicho, al tener diferentes rasgos para enfrentar limitaciones similares y coexistir (Andersen et al., 2012). Esto también nos permite agrupar a C. pinnatifrons y G. orbignyana como palmas estrictas de sotobosque, y a C. linearis y G. undata como palmas con mayor flexibilidad para ocupar otros hábitats, como bosques secundarios o expuestos a mayor radiación lumínica.
A través del estudio de las tasas vitales, en este trabajo, se presentan las estrategias de crecimiento de Chamaedorea linearis, C. pinnatifrons, Geonoma orbignyana y G. undata en las condiciones limitantes del sotobosque. Estos resultados pueden considerarse como un paso para conocer las estrategias de una comunidad de palmas a sus condiciones ambientales, y esperamos que puedan servir en el diseño de planes de conservación y uso basados en estas estrategias de crecimiento.
Declaración de ética: los autores declaran que todos están de acuerdo con esta publicación y que han hecho aportes que justifican su autoría; que no hay conflicto de interés de ningún tipo; y que han cumplido con todos los requisitos y procedimientos éticos y legales pertinentes. Todas las fuentes de financiamiento se detallan plena y claramente en la sección de agradecimientos. El respectivo documento legal firmado se encuentra en los archivos de la revista.

