











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
La familia Scolopacidae (Orden: Charadriiformes) constituye una de las familias que cuenta con la mayoría de las especies migratorias neárticas en el Perú (García-Olaechea et al., 2018; Rappole et al., 1993) con 37 especies a nivel nacional (Plenge, 2021). El Humedal Costero Poza de la Arenilla (HCPA) ubicado en el distrito de la Punta, Callao, cuenta con espacios adecuados que sirven para el reposo y alimentación de estas aves acuáticas (Bildstein et al., 1991; Cotillo et al., 2018; Podestá et al., 2017; Podestá & Cotillo, 2016; Pulido Capurro & Bermúdez Díaz, 2018), llegando a sumar un total histórico de 20 especies, superior a otros humedales del Callao (Podestá et al., 2021) como el Área de Conservación Regional Humedales de Ventanilla (ACRHV) que registra 18 especies de escolopácidos (Carazas et al., 2015). Los eventos climáticos podrían afectar la distribución de los organismos acuáticos (Cursach et al., 2011; Jaksic & Fariña, 2010), que sirven de presas a muchas poblaciones de aves, impactando negativamente en sus abundancias y procesos ecológicos como la migración (Crick, 2004; Durant et al., 2004; Lluch-Belda et al., 2000; McKinney et al., 2011). Los eventos El Niño y La Niña afectan a las poblaciones de aves guaneras y su principal recurso alimenticio: la anchoveta (Cursach et al., 2017; Jaksic & Fariña, 2010; Perlaza-Gamboa et al., 2020). Espinosa-Pelaez (2016) menciona que la población de aves guaneras presenta variaciones en su distribución dependiendo de la ocurrencia de El Niño y La Niña. Por otro lado Bouchon-Corrales et al., 2015 mencionan que el evento de 2014 afectó negativamente la abundancia de la anchoveta en el litoral peruano.
Existen diferentes índices para determinar el Niño Costero, siendo uno de ellos el Índice Costero El Niño (ICEN) (Takahashi et al., 2014), que indica la ocurrencia de eventos La Niña o El Niño. La Temperatura superficial del Mar (TSM) es conocida también como una variable predictora para diferentes eventos climáticos globales, entre los cuales uno de los más importantes es el fenómeno del Niño-Oscilación del Sur (ENOS) (Bakun et al., 2015; Jaksic & Fariña, 2010). Sin embargo, la TSM también influye sobre las precipitaciones y la temperatura atmosférica, siendo también un indicador importante de las condiciones ecológicas de diferentes especies marinas (Hernández, 2002; Perlaza-Gamboa et al., 2020). Galbraith et al. (2014) predijeron las vulnerabilidades de las aves playeras frente al cambio climático, ya que estas dependen del recurso alimenticio que proveen los huevos del cangrejo herradura (Limulus polyphemus) en Norteamérica, cuya puesta de huevos podría verse afectada por los cambios de temperatura (Bart et al., 2007; Piersma & Lindström, 2004; Whitney, 2014). La distribución, abundancia y riqueza de aves en los andes tropicales y planos intermareales (especialmente aves playeras) es también afectada por el Fenómeno del Niño (Cárdenas & Hurtado, 2019; Johnston-González et al., 2020). La familia Scolopacidae se encuentra en el apéndice I y II del listado de especies con un estado de conservación desfavorable de la Convención sobre la Conservación de las Especies Migratorias de Animales Silvestres, tratado que busca orientar esfuerzos para la conservación de especies migratorias (CMS, 2020) y alberga especies clasificadas como casi amenazadas (NT) a nivel internacional, entre las que destacan Calidris canutus, Calidris pusilla y Calidris ferruginea (BirdLife International, 2021).
El objetivo principal del presente trabajo fue determinar la relación entre la abundancia y riqueza de especies pertenecientes a la familia Scolopacidae y la TSM e ICEN en el Humedal Costero Poza de la Arenilla durante el período 2013-2019. Conocer esta relación es de vital importancia tanto para la conservación de los escolopácidos como para la protección de los hábitats que utilizan.
Materiales y métodos
Área de estudio: El Humedal Costero Poza La Arenilla (HCPA) se encuentra en el distrito de la Punta, Provincia Constitucional del Callao, Perú, entre las coordenadas (12°04’50.32” S - 77°09’15” W & 12°04’18.39” S - 77°09’40.65” W) (Fig. 1A, Fig. 1B). Este humedal artificial costero comprende 18.2 ha y posee dos cuerpos de agua como consecuencia de la existencia de dos bocanas, ubicadas en los extremos este y oeste. Los cuerpos de agua se encuentran separados por una zona de arenilla con extensión variable y dependiente del nivel de la marea. El humedal colinda con el ambiente urbano a través de un malecón que permite la observación de aves y otras actividades (Fig. 1). La arenilla posee una profundidad máxima de 2 m y posee diferentes especies de fauna y flora, pertenecientes a grupos como aves, peces y crustáceos (Sánchez Rivas et al., 2014). Históricamente se han identificado 14 zonas o hábitats (Troll, 2000), habiéndose reducido a 11 en la actualidad (Cotillo et al., 2018).
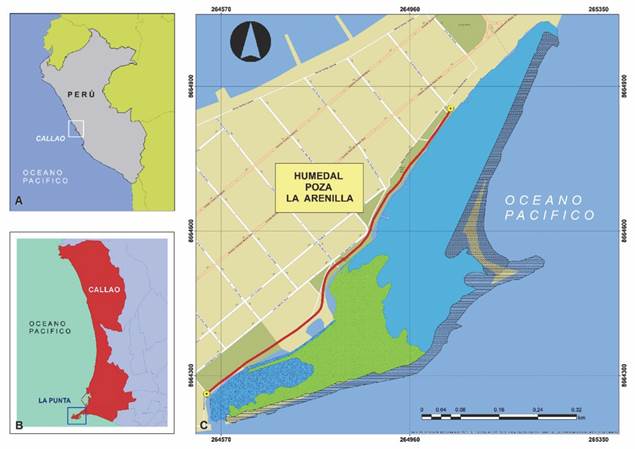
Fig. 1 Área de estudio. A. Ubicación de País. B. Ubicación de la Provincia Constitucional del Callao. C. Ubicación del Humedal Poza La Arenilla, La Punta, Callao: se muestra el transecto evaluado a lo largo del borde de la zona urbana. / Fig. 1. Location of the study area. A. Country Location. B. Location of the Constitutional Province of Callao. C. Location of the Poza La Arenilla Wetland, La Punta, Callao: the transect evaluated along the edge of the urban area is shown.
Obtención de datos abióticos: Los datos mensuales de la TSM fueron obtenidos a través del portal de libre acceso de la IMARPE (Instituto del Mar del Perú - www.satélite.imarpe.gob.pe/uprsig/sst_prov.html). De igual manera, los valores mensuales del ICEN se obtuvieron a través del portal web del Instituto Geofísico del Perú (www.met.igp.gob.pe/datos/icen.txt).
Muestreo de campo: Se realizaron muestreos quincenales entre enero 2013 y enero 2019, en la mañana (7 - 11 am) y la tarde (2 - 4 pm) (N = 292), utilizando el método del conteo total (Alegría Mont, 2018; MINAM, 2015; Sainz-Borgo et al., 2018; Torres et al., 2006) a lo largo de un transecto (Fig. 1C). Se registró la riqueza de especies y sus abundancias. Se utilizaron binoculares (10 x 42 mm) y una cámara fotográfica con lentes de 70-200 mm. El método de conteo por bloque, que consiste en contar el número de individuos de pequeñas submuestras de la totalidad de la bandada para luego estimar la población total (Bibby et al., 2000), se utilizó cuando las bandadas fueron mayores a los 500 individuos. Las abundancias obtenidas según los métodos de conteo total y por bloque fueron utilizadas de manera conjunta para el análisis posterior.
Se analizó gráficamente la variación anual de la TSM e ICEN, y la variación estacional de la abundancia y riqueza de especies, utilizando la data perteneciente al periodo comprendido entre enero 2013 y enero 2019. Se realizó un análisis de correlación de Spearman entre los valores mensuales de abundancia, riqueza, TSM e ICEN. La prueba de Spearman ha sido utilizada previamente para analizar la correlación entre parámetros a nivel de comunidad biológica y parámetros abióticos (Barona, 2021; Rubio et al., 2011). Para este análisis correlacional se tomó en cuenta el periodo desde enero de 2013 hasta diciembre de 2018.
Se aplicó la prueba ANOSIM para determinar las disimilitudes en las abundancias medias entre estaciones para todo el periodo entre enero 2013 y diciembre 2018. Luego, se aplicó la prueba post-hoc SIMPER, para determinar qué especies son las que más contribuyen con las diferencias en abundancias entre estaciones (Cotillo et al., 2018). Finalmente, se calculó la diversidad beta a través del índice de Whittaker (βw), para analizar tanto el recambio anual para cada estación como el recambio estacional dentro de cada año.
Resultados
Se registraron 14 especies de escolopácidos, todos ellos migratorios neárticos (Tabla 1). Se obtuvieron únicamente dos correlaciones significativas negativas: entre abundancia y TSM (ρ = -0.39, P = 0.0007) y entre riqueza e ICEN (ρ = -0.24, P = 0.04), y se construyeron gráficas de variación temporal para TSM e ICEN (Fig. 2A, Fig. 2B, Fig. 3A, Fig. 3B).
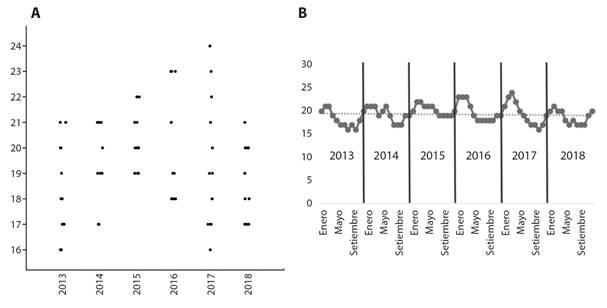
Fig. 2 A. Diagrama de dispersión (jitter-plot) que muestra la variación anual de la TSM y B. variación anual por meses de la TSM en el HCPA, Callao, Perú entre 2013 y 2018. / Fig. 2. A. Scatter diagram (jitter-plot) that shows the annual variation of the TSM and B. annual variation by months of the TSM in the HCPA, Callao, Peru between 2013 and 2018.
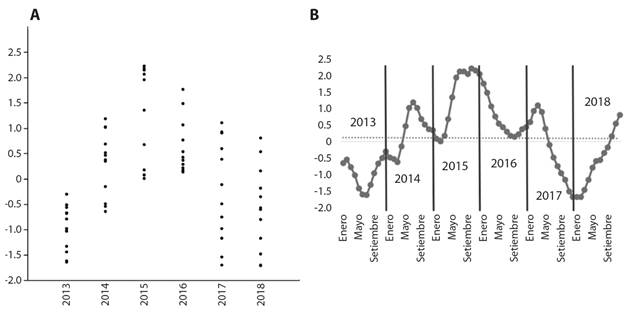
Fig. 3 A. Diagrama de dispersión (jitter-plot) que muestra la variación anual del ICEN y B. variación mensual del ICEN en el HCPA, Callao, Perú entre 2013 y 2018. / Fig. 3. A. Scatter diagram (jitter-plot) that shows the annual variation of the ICEN and B. annual variation by months of the ICEN in the HCPA, Callao, Peru between 2013 and 2018.
Tabla 1 Lista de especies de escolopácidos migratorios neárticos registradas en el HCPA, Callao, Perú entre 2013 y 2019 (N = 292). / Table 1. List of species for nearctic migratory scolopacids registered in HCPA, Callao, Peru, between 2013 and 2019 (N = 292).
| Especie | Nombre común | Estado de conservación |
| Numenius phaeopus | Zarapito Trinador | LC |
| Arenaria interpres | Vuelvepiedra rojizo | LC |
| Calidris canutus | Playero de Pecho Rufo | NT |
| Calidris virgata | Chorlo de las Rompientes | LC |
| Calidris himantopus | Playero de Pata Larga | LC |
| Calidris alba | Playero Arenero | LC |
| Calidris pusilla | Playerito Semipalmado | NT |
| Calidris minutilla | Playerito Menudo | LC |
| Calidris mauri | Playerito Occidental | LC |
| Phalaropus tricolor | Faláropo Tricolor | LC |
| Actitis macularius | Playero Coleador | LC |
| Tringa melanoleuca | Playero Pata Amarilla Mayor | LC |
| Tringa semipalmata | Playero Ala Blanca | LC |
| Tringa flavipes | Playero Pata Amarilla Menor | LC |
LC: Preocupación menor; NT: Casi amenazada.
También se observó una tendencia al descenso gradual en cuanto a la riqueza y abundancia para el periodo 2013-2019 (Fig. 4A, Fig. 4B). La prueba ANOSIM arrojó diferencias estacionales en las abundancias (R = 0.445, P = 0.0001), y la prueba SIMPER arrojó que la mayor diferencia en las abundancias entre estaciones fue entre otoño y primavera (disimilitud de Bray-Curtis = 81.57 %), y la menor entre invierno y primavera (disimilitud de Bray-Curtis = 49.86 %) (Tabla 2).
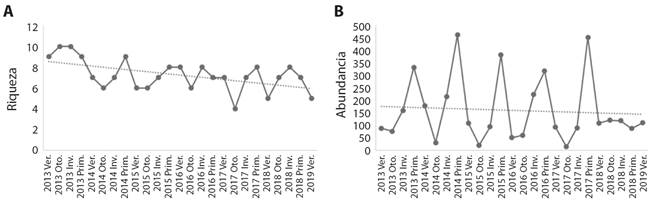
Fig. 4 A. Variación estacional de la riqueza y B. la abundancia para el periodo 2013-2019 en La Arenilla. / Fig. 4. A. Seasonal variation of richness and B. abundance for the period 2013-2019 in La Arenilla.
Tabla 2 Diferencias estacionales de las especies de la familia Scolopacidae registradas en el HCPA durante el periodo 2013-2019. / Table 2. Seasonal differences of the species of the Scolopacidae family registered in the HCPA during the period 2013-2019.
| Verano | Otoño | Invierno | Primavera | |
| Verano | - | Numenius phaeopus (31.7 %) 67.43 | Calidris alba (32.9 %) 50.93 | Calidris alba (62.7 %) 62.35 |
| Otoño | - | - | Calidris alba (36.1 %) 68.13 | Calidris alba (56.8 %) 81.57 |
| Invierno | - | - | - | Calidris alba (62.5 %) 49.86 |
| Primavera | - | - | - | - |
Para cada comparación pareada entre estaciones se muestra la especie que más contribuye con las diferencias y el porcentaje de contribución a dicha diferencia (a través del análisis SIMPER). El texto en negrita muestra la disimilitud de Bray-Curtis (a través del análisis ANOSIM).
El análisis SIMPER mostró que, en términos generales, Calidris alba es la especie que más contribuye con las diferencias observadas entre todas las estaciones, a excepción de otoño y verano, en donde la especie que más contribuye a las diferencias es Numenius phaeopus (Tabla 2).
El recambio anual global para cada estación a través del índice beta de Whittaker (βw) arrojó que el recambio general de especies entre todos los veranos desde 2013 a 2019 fue de 0.64, entre todos los otoños desde 2013 al 2018 fue de 0.69, entre todos los inviernos desde 2013 al 2018 fue de 0.4, y entre todas las primaveras desde 2013 al 2018 fue de 0.25. Adicionalmente, se calculó también el índice de Whittaker para medir el recambio de un año a otro para cada estación (Tabla 3).
Tabla 3 Recambio anual por estación de la comunidad de escolopácidos en el HCPA durante el periodo 2013-2019. Los valores mostrados corresponden al índice de Whittaker. / Table 3. Annual turnover by season of the scolopacid community in HCPA during the period 2013-2019. The Whittaker index values are shown.
| Verano | Otoño | Invierno | Primavera | |
| 2013-2014 | 0.25 | 0.38 | 0.18 | 0.11 |
| 2014-2015 | 0.08 | 0.33 | 0 | 0.06 |
| 2015-2016 | 0.29 | 0.5 | 0.2 | 0.2 |
| 2016-2017 | 0.33 | 0.6 | 0.2 | 0.07 |
| 2017-2018 | 0.17 | 0.45 | 0.2 | 0.07 |
| 2018-2019 | 0.2 | - | - | - |
En cuanto al recambio estacional dentro de cada año medido a través del índice de Whittaker, fue 0.37 para el 2013, 0.38 para el 2014, 0.19 para el 2015, 0.93 para el 2016, 0.54 para el 2017, y 0.48 para el 2018. También se calculó el índice de Whittaker para medir el recambio de una estación a la siguiente para cada año (Tabla 4).
Tabla 4 Recambio estacional por año de la comunidad de escolopácidos en el HCPA durante el periodo 2013-2019. Los valores mostrados corresponden al índice de Whittaker. / Table 4. Seasonal turnover per year of the scolopacid community in HCPA during the period 2013-2019. The Whittaker index values are shown.
| 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | |
| Ver-Oto | 0.16 | 0.38 | 0 | 0.71 | 0.27 | 0.5 |
| Oto-Inv | 0.3 | 0.23 | 0.23 | 0.43 | 0.64 | 0.33 |
| Inv-Prim | 0.16 | 0.13 | 0.07 | 0.2 | 0.33 | 0.07 |
| Prim-Ver | 0.11 | 0.25 | 0.14 | 0.47 | 0.07 | 0.17 |
Discusión
Se observaron 14 especies de escolopácidos migratorios neárticos durante el período de estudio en el HCPA. Esto demuestra la importancia del humedal como refugio para estas aves, siendo comparable a lo observado en otros humedales de la costa peruana: San Pedro de Vice con 12 especies (García-Olaechea et al., 2018), Pantanos de Villa con 12 especies (Torres et al., 2006), humedales de Etén con 14 especies (Angulo-Pratolongo et al., 2010) y Humedales de Ventanilla con 18 especies (Carazas et al., 2015). Los cambios en la temperatura y la disponibilidad de alimento modifican el comienzo y final de las migraciones (Jonzén et al., 2006). Esto significa que temperaturas anormales podrían afectar la época de inmigración y emigración de las aves debido a la alteración de sus hábitats. Para Países Bajos se ha reportado correlaciones negativas entre la distancia de migración y la temperatura debido al cambio climático (Chen et al., 2011; Visser et al., 2009). La disminución gradual de la riqueza de especies observada para HCPA podría deberse al incremento sostenido en el ICEN, observado desde 2013 hasta 2015.
Los resultados del presente estudio muestran que se produjo una ligera alza en la TSM durante 2015 y 2016 en comparación a otros años, particularmente durante los meses de invierno y primavera, aunque en 2017 se observó una amplitud térmica anual más marcada (Fig. 2A). También para 2015 y 2016 se observa un aumento en el ICEN, aunque ello no parece haber afectado notoriamente a la riqueza y abundancia de escolopácidos. Además, los valores de ICEN de 2015 fueron los mayores de todo el periodo (Fig. 3A). Sin embargo, para todo el periodo evaluado (2013-2019) existe una débil correlación inversa entre la abundancia y TSM, y entre la riqueza e ICEN. Además, la disminución gradual observada tanto en la abundancia como en la riqueza de especies aparentemente tiene una relación baja o no significativa con los factores abióticos abordados en el presente estudio, pero dada la tendencia a largo plazo de incremento de la TSM y de intensificación de los eventos El Niño y La Niña (medidos a través del ICEN), la disminución observada en la abundancia y riqueza podrían ser parte de un efecto negativo a largo plazo sobre las comunidades de aves (Sutherland et al., 2012), como ocurre en latitudes polares y subpolares, donde existe una tendencia de disminución de las abundancias poblacionales de varias especies de aves como respuesta al cambio climático y a las presiones antropogénicas (Raya Rey et al., 2014).
La baja diferencia en la disimilitud de las abundancias entre invierno y primavera puede deberse a registros más frecuentes de “singletons” (especie con registro único) como Calidris himantopus (García-Olaechea et al., 2018; Podestá et al., 2017). Si bien estas especies poco abundantes no contribuyen significativamente a los cambios generales de la abundancia, sí contribuyen a los cambios en riqueza en humedales (García-Olaechea et al., 2018). También resulta importante que en Poza de la Arenilla se han evidenciado registros de especies de escolopácidos migratorios neárticos en invierno y primavera, épocas en que normalmente no se espera encontrarlos (Cotillo et al., 2018; Podestá et al., 2017).
Por otro lado, Calidris alba es la especie que más contribuye con las diferencias observadas entre estaciones, lo cual ya había sido previamente notado en HCPA por Podestá et al. (2017). Algo similar ocurre en el hemisferio norte, como es el caso del Golfo de Santa Clara (Sonora, México) en donde C. alba se encuentra entre las especies de aves más abundantes, pero con alta fluctuación en abundancia entre una estación y la siguiente, lo cual se debe parcialmente a la perturbación por bañistas (Carmona & Hernández-Alvarez, 2017).
Verano y otoño son las estaciones que mostraron el mayor recambio de especies para todo el periodo de evaluación, lo cual se debe al momento en que las aves playeras migran hacia Norteamérica. Algo similar se observa para el recambio anual para cada estación: en verano y otoño se observan cambios de mayor magnitud que en invierno y primavera. Los mayores recambios estacionales anuales se dan en los tres últimos años del período evaluado, siendo particularmente alto para el 2016, cuando se da el mayor recambio de especies de todo el período: βw = 0.71 entre verano y otoño. Esto puede deberse a la influencia de la variación de TSM sobre la dinámica poblacional de diferentes especies marinas que sirven de alimento a los escolopácidos (Perlaza-Gamboa et al., 2020; Sydeman et al., 2012). Los cambios en temperatura producidos por el ENOS también constituyen la principal causa en la demora en la inmigración y emigración de las aves, lo cual se traduce en una variación en el recambio de especies (Johnston-González et al., 2020; Perlaza-Gamboa et al., 2020). El aumento de temperaturas relacionado tanto a El Niño como al cambio climático, además del descenso de las mismas por eventos La Niña modifica el comportamiento y fisiología de las aves marinas y su distribución. Esto podría tener consecuencias en el retraso y reducción de nidadas (Guerra et al., 1988), cambios en su alimentación, comportamiento agonístico entre diferentes individuos e incluso alta mortalidad de las especies más sensibles/especializadas (Marra et al., 2005).
Durante El Niño de 1983, en el litoral costero chileno, varias especies de gaviotas mostraron retrasos en ciclos gonadales, muda, y falta de apareamiento, interrumpiéndose así la reproducción de Leucophaeus modestus, disminuyendo su población drásticamente. También afectó a Engraulis ringens (Anchoveta) y Emerita analoga (Muy Muy) que se encuentran entre los principales alimentos de L. modestus (Guerra et al., 1988). Además, los eventos El Niño de 1983 y 1998 afectaron sobremanera a las especies de aves marinas peruanas, siendo las más afectadas las endémicas de la Corriente de Humboldt (Sula variegata, Pelecanus thagus, Phalacrocorax boungavilii, Sphenicus humboldtii) (Apaza & Figari, 1999).
En conclusión, la variación en TSM e ICEN serían importantes factores implicados en la reducción de la abundancia y riqueza de especies de escolopácidos en el HCPA, ya que los cambios a mediano y largo plazo de los factores ambientales térmicos interfieren de forma negativa principalmente sobre sus recursos alimenticios, patrones de migración, reproducción, entre otros.
Declaración de ética: los autores declaran que todos están de acuerdo con esta publicación y que han hecho aportes que justifican su autoría; que no hay conflicto de interés de ningún tipo; y que han cumplido con todos los requisitos y procedimientos éticos y legales pertinentes. Todas las fuentes de financiamiento se detallan plena y claramente en la sección de agradecimientos. El respectivo documento legal firmado se encuentra en los archivos de la revista.

