











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkIntroduction
The Eastern Tropical Pacific (ETP) is a region that has unique oceanographic and environmental conditions for planktonic and higher trophic level organisms that inhabit it (Chavez et al., 1999; Lavín et al., 2006). This region extends from the Southwestern coast of Baja California to northern Peru and has a strong variability that operates at different scales of space and time in the conditions that characterize it (Amador, Alfaro, Lizano, & Magaña, 2006; Fiedler & Lavín, 2016), such as the interannual variability generated by El Niño-Southern Oscillation (ENSO) event which is probably one of the oceanic-atmospheric processes that has the highest incidence in the ETP, both in its warm phase (El Niño), and in its cold phase (La Niña) (Montealegre, 2007; Fiedler & Lavín, 2016).
One of the most outstanding aspects of ENSO is the unusual variations in the sea surface temperature (SST) (Wang & Fiedler, 2006) since this oceanographic parameter significantly affects the atmosphere and also generates a physiological, ethological and ecological response in marine organisms (O’Connor et al., 2007; Côté & Green, 2015; Gunderson, Armstrong, & Stillman, 2016). The effects of the SST variation caused by ENSO on marine organisms have been historically focused on establishing their relationship with the dynamics of primary consumers and planktonic producers such as phytoplankton, copepods or larvae of commercial interest organisms such as fish larvae, squid and crustaceans (Paredes, Cardoso, & Tarazona, 2004; Masotti et al., 2011; Kozak, Franco-Gordo, Suárez-Morales, & Palomares-García, 2014; León-Chávez, Beier, Sánchez-Velasco, Barton, & Godínez, 2015; Alabia et al., 2016). However, these taxonomic groups directly affect the survival, reproduction, and distribution of higher trophic level organisms like mammals, sea birds, and sea turtles (Ballance, Pitman, & Fiedler, 2006; Polidoro et al., 2012).
The ETP is also one of the most important regions for marine turtles Hawksbill (Eretmochelys imbricata), Leatherback (Dermochelys coriacea), Green (Chelonia mydas), Loggerhead (Caretta caretta) and Olive Ridley (Lepidochelys olivacea), since it shelters extensive nesting beaches and feeding sites, where these species have access to a diversity of marine habitats such as sea grass pastures, mangrove estuaries, open waters and coral and rocky reefs (Seminoff et al., 2012). According to Pritchard and Plotkin (1995), throughout Central America, warming due to El Niño conditions could be partially responsible for declines in the number of nesting females of Lepidochelys olivacea. In this species, changes have also been reported in their post-nesting migratory routes in response to the El Niño effect (Plotkin, 2010).
L. olivacea is considered the most abundant sea turtle in the world, despite being listed as a vulnerable species according to the red list of the International Union for the Conservation of Nature-IUCN (IUCN, 2019). In addition to the ETP, this species is distributed in tropical waters of the Indian and South Atlantic Ocean basins (Barrientos-Muñoz, Ramírez-Gallego, & Páez, 2015). In Colombia, it is the most common nesting turtle of the Pacific (Amorocho, Rodríguez-Zuluaga, Payán, Zapata, & Rojas, 2015), despite this, studies on its reproductive biology in Colombia have been reduced, spaced in time and carried out in a few locations (Amorocho, Rubio, & Díaz, 1992; McCormick, 1996; Martinez & Páez, 2000; Hinestroza & Páez, 2001; Pavía, Rodríguez-Zuluaga, & Amorocho, 2006; Barrientos-Muñoz & Ramírez-Gallego, 2008; Barrientos-Muñoz, Ramírez-Gallego, & Páez, 2014; Payán-Perea, Zorrilla-Arroyave, & Chirimía-Gonzáles, 2016). In this context, it is relevant to evaluate the effect that thermal variations associated with ENSO could have on marine turtle species of the Colombian Pacific, particularly on the different attributes related to the nesting process of L. olivacea in the marine protected area of Gorgona island.
For this purpose, the historical record of the “Marine Turtle Monitoring Program of Gorgona National Natural Park (PNNG)” was used to consolidate the monthly information related to females, nests, eggs and hatchlings of L. olivacea between 2008 and 2018 in Playa Palmeras. This information was correlated with SST of Gorgona island and Panama Bight, and with the indexes of El Niño 1+2, El Niño 3, El Niño 3.4 and El Niño 4 regions, their thermal anomalies, and the ONI and SOI indexes, with the objective of evaluating the effect of the SST on nesting attributes of L. olivacea in Playa Palmeras, Gorgona island, considering the oceanic variability both on a local and regional scale.
Materials and methods
Study area: Gorgona Island (2°55’45” - 3°00’55” N & 78°09’ - 78°14’30” W) is located in the south of the Colombian Pacific, 30 km from the coast of Cauca and Nariño departments. It is 9 km long and 2.5 km wide (Giraldo, Rodríguez-Rubio, & Zapata, 2008; Amorocho, Rodríguez-Zuluaga, Payán, Zapata, & Rojas, 2015) and the surface waters remain relatively constant throughout the year, with warm temperatures between 26 and 28.6 °C (Giraldo, Valencia, Acevedo, & Rivera, 2012). In 1984, the island and its marine area, were declared Natural National Park, with the aim of preserving species and ecosystems representative of terrestrial and marine flora and fauna of the Pacific. Marine turtles are considered a value object of conservation for the PNNG, reason for which permanent monitoring efforts have been directed since 2008 (Amorocho et al., 2015; Barrientos-Muñoz et al., 2015; Payán-Perea et al., 2016).
In Gorgona Island, sea turtle nesting monitoring is carried out in Playa Palmeras, a sandy beach located at the Southwestern end of the island (2°56’38’’ N - 78°11’53’’ W & 2°56’19’’ N - 78°12’16’’ W). It has a total extension of 1.2 km, being the largest sandy beach on Gorgona. For nesting monitoring, the beach has been divided in six sectors, every 200 meters, numbered from north to south (Payán-Perea et al., 2016).
Data collection: “PNNG Marine Turtle Nesting Monitoring” consists of night patrols between July and November, along Playa Palmeras between 20:00 and 5:00 h if climatic conditions and tides are optimal, in order to intercept nesting turtles and/or search for traces and nests. The staff willing to take data is at least two people per night.
For each turtle found, curved carapace length (CCL) and curved carapace width (CCW) measurements are taken using a tape measure and, straight carapace length (SCL) and straight carapace width (SCW) are taken using a caliper. Subsequently, they are marked with one metal tag in the second scale of the anterior fin, placing the tags marked with an odd number on the left fin and with an even number on the right. After identifying a nest, the eggs are relocated to a hatchery if the nesting site poses a risk of flooding. In each relocated nest, number of eggs is recorded and ten eggs are randomly selected to record the diameter using a caliper and the weight using a 50 g weight. Forty-five days after nesting, the nests are monitored until the emergence of hatchlings. Once they emerge, they are counted and ten are taken at random to register their weight, straight carapace length, and straight carapace width. Finally, the hatchlings are released in the sand.
A database was consolidated from the record sheets of this monitoring program, establishing monthly information on the number of visits to the beach, defined as any evidence of arrival of a turtle (VB), number of nesting females (NNF), number of nests (NN), clutch size, defined as the number of eggs per nest (CZ), incubation days (ID), hatching success (HS), defined as the percentage of live hatchlings over the CZ, diameter (ED) and weight (EW) of eggs, straight carapace length (SCLh), straight carapace width (SCWh) and weight (Wh) of the hatchlings of L. olivacea, from 2008 to 2018, for a total of eleven years of monitoring data. The number of nesting females was estimated from the method of Alvarado and Murphy (1999), where the number of nests is divided by the average number of nests per female per year (2.5 for Lepidochelys sp.).
The SST information for Gorgona (GOR) and Panama Bight (PB) (local scale), was obtained from monthly images of level 3 with 4 km resolution from NASA’s MODIS- Aqua sensor (Ocean Biology Processing Group, 2015). For GOR, the SST information was extracted from the CPC16 station of ERFEN grid (Program for the Regional Study of El Niño Phenomenon in the Southeast Pacific) (3°0’0’’ N - 78°0’0’’ W). For PB, information was extracted from a sampling grid of 86 stations distributed in Panama Bight and separated by 1° of latitude and longitude. The extraction of information from the images was carried out through virtual samplings using ArcGIS 10.5.
For regional SST, monthly information was obtained from climatic variability indexes: El Niño Regions 1+2, 3, 3.4 and 4, their thermal anomalies (ANOM 1+2, ANOM 3, ANOM 3.4 and ANOM 4) and the ONI (Oscillation Niño Index) and SOI (Southern Oscillation Index) indexes from the NOAA (National Oceanic and Atmospheric Administration) Climate Prediction Center, ERSSTv5 dataset (Climate Prediction Center Internet Team, 2017).
Monthly and annual descriptive analyzes of nesting variables were carried out and subsequently, monthly time series were constructed with SST information, indexes and anomalies and the nesting attributes of L. olivacea, taking only the nesting records from July to November. With this set of time series, cross correlations were made to evaluate the relationship between nesting variables and climatic variability indexes, both in simultaneous (lag 0), and at a later time. The nesting variables were lagged in 24 months. The lags with the highest correlations were included in Generalized Additive Models (GAM). All analyzes were done with STATISTICA 7.0 (StatSoft, 2004).
Results
Between 2008 and September of 2018, a total number of 513 L. olivacea arrival records and 486 nests were obtained, for which the NNF was estimated in 194; There were 221 encounters with turtles, of which 169 were females tagged for the first time and 39 were recaptures. The average measurements, in centimeters, of the intercepted females were: CCL = 65.37 ± 2.44; CCW = 69.34 ± 3.20; SCL = 60.83 ± 2.62 and SCW = 54.34 ± 3.02. Fig. 1 shows the average monthly records of VB, NNF and NN, where it is identified that September (month 9), was the month in which the highest average of records was obtained for this three variables (NN = 17.72 ± 6.26, VB = 17.09 ± 8.89, NNF = 7.09 ± 2.50).
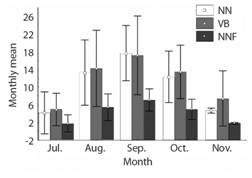
Fig. 1 Monthly nesting records of Lepidochelys olivacea in Playa Palmeras, Gorgona island, during July-November (7-11). Period evaluated: 2008-2018 (11 years). NN = Number of nests/month; NNF = Number of nesting females/month; VB = Number of visits to the beach/month.
There were 32 864 eggs counted corresponding to 347 relocated nests (71.4 % of total). The average CZ was 94.71 ± 19.32 and average ID was 57.03 ± 4.60. The average diameter and weight of 2 968 eggs was: ED = 3.75 ± 0.25 cm and EW = 32.91 ± 3.9 g. Related to hatchlings, there were 17 730 births recorded corresponding to 249 nests (49.17 % of total nests). In these, the total average hatching success was 67.07 %.
In the eleven years period, it was possible to identify interannual variations in all the nesting variables. Fig. 2 and Fig. 3 show the monthly averages of the nesting variables in each year. 2010 was year with the lowest averages of HS (51 %), ED (3.39 ± 0.27 cm), SCLh (3.74 ± 0.19 cm) and SCWh (2.96 ± 0.21 cm); 2008, the one with the lowest CZ (84.78 ± 16.89) and 2015, the lowest ID (54.59 ± 3.99). On the contrary, the years with the highest monthly averages were: 2013 with ED= 3.91 ± 0.20 cm and SCWh= 3.61 ± 0.24 cm; 2014 with HS= 85.73 %, SCLh= 4.34 ± 0.19 cm and EW = 35.23 ± 3.97 g, 2 017 with Wh = 20.35 ± 1.02 g; 2012 the highest CZ (100.42 ± 11.08) and 2009 the highest ID (59.88 ± 5.24).
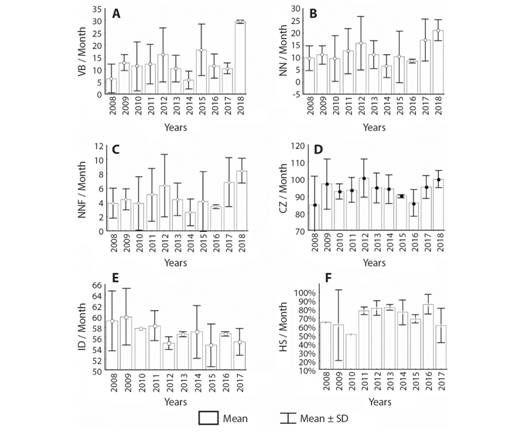
Fig. 2 Annual nesting records of variables related to females and nests of Lepidochelys olivacea in Playa Palmeras, Gorgona Island. Period evaluated 2008-2018 A. Annual average of visits to the beach per month B. Annual average of nests per month; C. Annual average of nesting females per month D. Annual average of size of nests per month; E. Annual average of incubation days per month; F. Annual average hatching success per month.
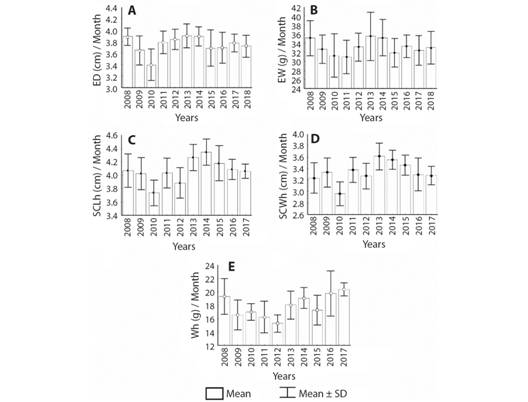
Fig. 3 Annual nesting records of variables related to eggs and hatchlings of Lepidochelys olivacea in Playa Palmeras, Gorgona Island. A. Annual average of egg diameter; B. Annual average of egg weight; C. Annual average of straight carapace length of hatchlings; D. Annual average of straight carapace width of hatclings; E. Annual average weight of hatchlings.
The cross-correlation analysis between the variables of females and nests NN, NHA, VP, TN, DI, EE and climatic indexes El Niño 1+2, 3, 3.4, 4, ONI, ANOM 1+2, ANOM 3, ANOM 3.4 and ANOM 4 and the SST of GOR and PB, indicated significant correlations, of which, the highest correlation since maximum correlation is 1, were negative. That is, when the SST decreased locally and regionally and there were negative anomalies, high values of the nesting variables were found. In the case of SOI, when it was in its positive phase, there was also an increase in the nesting variables. The Niño 1+2 index presented the highest significant negative correlations for these variables, with a 12-month lag, followed by the GOR SST, in which the highest significant lag was 8-9 months. However, high positive significant correlations were also identified in lags 0 and 6 with Niño 1+2. For all cases, with these variables, the lag of highest correlation was equal to or less than 12 months.
For the morphometric variables of eggs and hatchlings, the EW and SCWh had highest negative correlations with all the indexes and SST, except with SOI. For the Wh the correlations were positive and negative with the SOI. The ED had negative correlations, except with Niño 3.4 and finally, with the SCLh both positive and negative correlations were obtained. The greatest lag with these variables was 19 months and the highest correlations were obtained with regional indexes. Fig. 4 shows the values of the highest significant correlations obtained in the cross-correlation tests between the pairs of indexes and SSTs and the nesting variables.
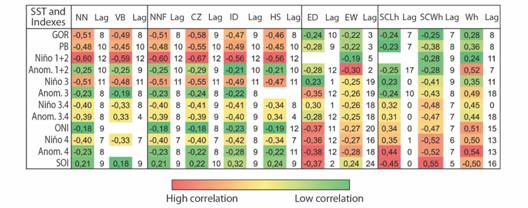
Fig. 4 Highest significant correlations obtained between climatic variability indexes and nesting variables of Lepidochelys olivacea at α = 0.05.
Through GAM (Fig. 5, Fig. 6) it was possible to ascertain the non-linearity between the nesting variables and the climatic indices. In the case of VB (Fig. 5A) and HS (Fig. 5F), low values were recorded both at positive and negative extreme values of ANOM 3.4. A slight tendency to reduction was identified in the NN (Fig. 5B), NNF (Fig. 5C) and the CZ (Fig. 5D), with positive values of ANOM 3.4. For ID (Fig. 5E) the trend seemed to be more stable, that is, the positive values of the index do not drastically reduce the ID. Regarding the morphometric variables of the eggs and hatchlings (Fig. 6), the trend was more pronounced. The tendency to decrease with positive values of the indexes was identified, with the variables ED (Fig. 6A), EW (Fig. 6B), SCLh (Fig. 6C) and SCWh (Fig. 6D). The Wh (Fig. 6E), however, increased with positive values.
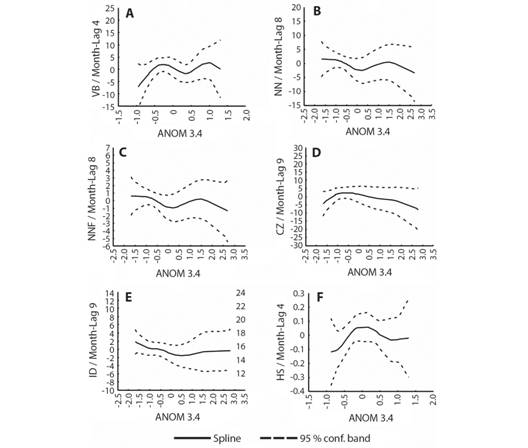
Fig. 5 Results of Generalized Additive Models (GAM) between lagged variables associated with females and nests of Lepidochelys olivacea and Niño 3.4. anomaly; A. Visits to the beach B. Number of nests; C. Nesting females; D. Clutch size; E. Incubation days; F. Hatching success.
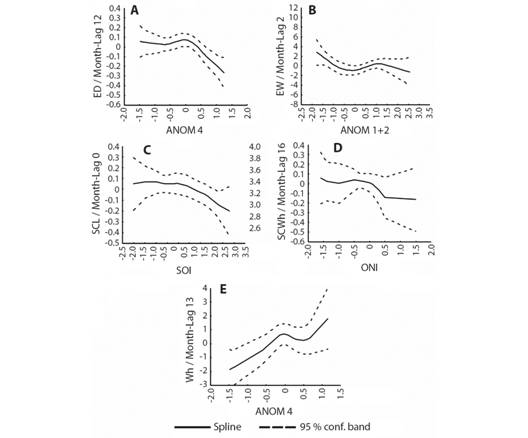
Fig. 6 Generalized Additive Models (GAM) between lagged variables related to eggs and hatchlings of Lepidochelys olivacea and climatic variability indexes. A. Eggs diameter and Niño 4 anomaly; B. Eggs weight and Niño 1+2 anomaly; C. Straight carapace length of hatchlings and SOI index; D. Straight carapace width of hatchlings and ONI; E. Weight of hatchlings and Niño 4 anomaly.
Discussion
The measurements of L. olivacea females found in the PNNG monitoring correspond with those reported on other beaches in the Colombian Pacific (Barrientos-Muñoz & Ramírez-Gallego, 2008; Hinestroza & Páez, 2001; Barrientos-Muñoz et al., 2014). Globally, the CCL of the nesting females of L. olivacea varies between 65.9 and 73.1 cm and those of the populations, between 60.8 and 83 cm, being the smallest marine turtle living species (Varo-Cruz, Monzón-Argüello, Carrillo, Calabuig, & Liriz-Loza, 2015). In general, in the Colombian Pacific the nesting season occurs from July to November, with a peak of nesting in september, as well as Gorgona island and there have also been few recaptures considering that this species, unlike others, nests annually and between two and three times per season (Amorocho et al., 1992, Amorocho et al., 2015; Barreto-Sánchez, 2011; Barrientos-Muñoz et al., 2015).
Regarding the relationship between SST and marine turtles, Mazaris, Kornaraki, Matsino and Margaritoulis (2004) identified through different models that this variable does represent an important factor for the reproductive behavior of sea turtles, in such a way that sea water temperature is a dynamic stimulus that affects processes associated with reproduction. This relationship has also been reported in C. mydas, (Solow, Bjorndal, & Bolten, 2002), C. caretta, (Chaloupka, Kamezaki, & Limpus, 2008; Mazaris, Kallimanis, Sgardelis, & Pantis, 2008), D. coriacea (Saba et al., 2007, Saba et al., 2008) and E. imbricata (Del Monte-Luna, Guzmán-Hernández, Cuevas, Arreguín-Sánchez, & Lluch-Belda, 2012), demonstrating that these effects are independent of the species and the turtle diet.
Particularly, Chaloupka et al. (2008)using 51-year-old nesting databases, concluded that regardless of whether the populations of C. caretta are growing or decreasing the general pattern is that there is an inverse correlation between the SST of the feeding areas and the abundance of the nesting population, even Del Monte-Luna et al. (2012) reproduced the same relationship with E. imbricata on a regional geographic scale. In the present study, this pattern was not only evident in the number of nests and nesting females, but also in the clutches sizes and in the egg diameter and weight of L. olivacea nesting in Playa Palmeras. It should be mentioned that the positive values of SOI, unlike the other indexes and anomalies coincide with abnormally warm waters in the ETP (Climate Prediction Center Internet Team, 2005), so the correlations found with this, correspond to the trend found with the other indexes.
Because of the energy demand experienced by mature females before reproduction that must be greater to carry out vitellogenesis, migration to nesting beaches and the nesting process itself (Hamann, Limpus, & Whittier, 2002; Wallace, Kilham, Paladino, & Spotila, 2006), a smaller egg size and weight could be consequence of a lower food availability, accompanied by an increase in the metabolic rate and consequently an accelerated vitellogenesis due to warmer waters in the feeding areas (Wallace & Jones, 2008). Generally, females require at least one year to store enough fat deposits to carry out this process (Miller, 1997), so the lags that resulted from the highest correlations (≤ 12 months) that were also consistent with the published by Chaloupka et al. (2008) and Mazaris et al. (2008), could reflect the required migration time between feeding and nesting areas.
In this context, possible deficiencies in nutrition, caused by decreases in food availability during 2009-2010 El Niño event, could have caused the low egg sizes recorded in Playa Palmeras (Gorgona island, Colombia) in 2010. This event not only was characterized by strong SST anomalies in the Central Pacific, but also was the event with the fastest transition from El Niño to La Niña registered until then (Kim, Yeh, Kim, Kug, & Kwon, 2011). However, the abnormally cold temperatures that occurred in 2010, did not produce the largest eggs nor the highest records in the other nesting variables in the following year.
Attributes such as nest size and number of nests are affected by a variety of factors, such as the size and physiological condition of the nesting female, and environmental constraints in nesting and feeding habitats (Miller, 1997). In this sense, the SST could be associated with these variables, however, others such as the hatching success, the incubation days and the physical state of the hatchlings, can change enormously depending on the beach, the sand type, the predation, the tidal regime, rains, sun exposure, etc. (Del Monte-Luna et al., 2012; Varo-Cruz et al., 2015) and probably, for this reason, the trend observed in this study with these variables were not marked and in some cases, contradictory. Additionally, it is very likely that longer time series of nesting records will be needed to establish this type of relationship.
The highest correlations for the variables related to females and nests were presented with the Niño 1+2 region, with a lag of twelve months. That is, the nesting season attributes would be related to the temperatures that occur in September of the previous year. However, high positive correlations were also found with this index, in other lags. This could be explained by the fact that in this zone there are high seasonal amplitudes in the SST (± > 3 °C) and marked seasonal and interannual variations (Fiedler & Lavín, 2016) so that individuals who remain in colder than optimal conditions could increase their reproductive output as the temperature increases, and those in optimal conditions may experience a decrease as the temperature increases (Mazaris et al., 2008). Another explanation would be that the variables studied would be determined by many factors and the SST is only one of them.
In general, nesting variables had higher correlations with regional SSTs, which is consistent with that reported by Del Monte-Luna et al. (2012) for Hawksbill turtles in the Atlantic. On the other hand, warmer temperatures in the SST of local waters would be more related to earlier beginnings of nesting seasons, which was not evaluated in this investigation. In addition to this, large-scale indexes tend to be better predictors of ecological processes than local variables (Hallett et al., 2004).
Moreover, through satellite telemetry, Chambault et al., (2016) studied the foraging habits of L. olivacea and documented that this species prefers to feed in cold waters, above or below the thermocline, in such a way that the deeper the thermocline is, olive ridleys must invest a greater amount of time and energy in submerging, so they could benefit from the cooler waters and the more shallow thermocline that occur in the Niño 1+2 region (Fiedler & Lavín, 2016) and the results obtained would suggest that they would be using this region, or part of it, as an important feeding area, so drastic changes in the SST and therefore in the thermocline of Niño 1+2 region , would be influencing the preparation prior to reproduction and nesting.
It has also been documented that seabirds that feed in this area (coast of Peru), experience reproductive failures and mortality during El Niño events due to food shortages, unlike those that feed in warmer areas of ETP, which are not severely affected (Ballance et al., 2006). It should be noted that the species and communities that inhabit the ETP have evolved to persist through the almost regular perturbations imposed by the ENSO events (Wang & Fiedler, 2006) and according to Paine, Tegner and Johnson (1998) this type of disturbances, when occurring in a typical range, do not cause major changes in species in the long-term, regardless how large and infrequent they turn out to be.
However, under the current climate change scenario, an increase in the occurrence and intensity of ENSO events has been projected, linked to a deepening and flattening of the thermocline in the equatorial Pacific (Ashok & Yamagata, 2009; Yeh et al., 2009) so, in the long term, this could negatively affect the reproductive aspects of L. olivacea, in Playa Palmeras and possibly, in other beaches of the ETP. With the results obtained in the present study, the need for information on the distribution and population trends at broad geographic and temporal scales is evident; therefore, it is important to perpetuate and promote long-term capture and tagging monitoring such as that carried out in the PNNG, as they are important for the quantification of changes in nesting variables and provide the basis for making decisions in regarding the management and conservation of sea turtles, specially Olive Ridleys in the Pacific.
Ethical statement: authors declare that they all agree with this publication and made significant contributions; that there is no conflict of interest of any kind; and that we followed all pertinent ethical and legal procedures and requirements. All financial sources are fully and clearly stated in the acknowledgements section. A signed document has been filed in the journal archives.

