











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
Haemulon plumieri se encuentra distribuida desde el sureste de Estados Unidos hasta Brasil, incluyendo el Mar Caribe y el Golfo de México. En Venezuela es capturada por la pesca artesanal principalmente con nasa, cordel y red. Habita en fondos someros, hasta ~ 40 m de profundidad con formaciones rocosas o coralinas, pero no asociados a ellas. Los ejemplares pequeños de 7 a 9.5 cm de longitud total se capturan en praderas de Thalassia (Cervigón, 1993). H. plumieri es una especie carnívora y los adultos se alimentan principalmente de invertebrados bentónicos, como crustáceos y poliquetos (Manooch, 1976). Es un recurso importante en las pesquerías artesanales del estado Nueva Esparta, e.g., la flota artesanal de El Tirano que faena en el archipiélago Los Frailes desembarcó 35.6 toneladas en la temporada de pesca 2012 representando el 4.11 % de las 51 especies de peces capturados (González, Eslava, Guevara, Díaz, & Rodríguez, 2017).
En el contexto de las pesquerías multiespecíficas no es fácil aplicar dispositivos de gestión y regulación pesquera, aun reconociendo su significación en la sostenibilidad biológica, económica y social. Sin embargo, es necesario realizar estudios de dinámica poblacional con la finalidad de evaluar los recursos, considerando los parámetros de crecimiento, reclutamiento y mortalidad, teniendo en cuenta que el número de individuos en cada clase anual va disminuyendo debido a la mortalidad natural o por pesca, siendo este último parámetro utilizado en la administración pesquera. Básicamente, hay una fracción de individuos que mueren por diversas causas y otra que no mueren y llegan al final del ciclo de vida (Csirke, 1980).
Se han realizado diversos estudios biológico-pesqueros de H. plumieri, principalmente en Norte América. Así, Manooch (1976), determinó la edad, crecimiento y mortalidad, Darcy (1983) realizó una sinopsis de su biología, Murphy, Murie y Muller (1999) y Potts (2000) describieron las características biológico-pesqueras, y estimaron la estructura de edad y tamaño de la población a través del análisis de población virtual (APV), mortalidad por pesca, mortalidad natural y reclutamiento. Silva y Murphy (2001) realizaron un resumen de su biología y pesquería en la costa este de Florida; Potts y Manooch (2001) y Murie y Parkyn (2005) estimaron la edad y crecimiento en Carolina y Florida, respectivamente. Por otra parte, en Brasil, Nomura (1965) analizó la relación longitud-peso, Araújo y Martins (2007) estudiaron edad, crecimiento y mortalidad; Mota y Aragao (1973) y Shinozaki-Mendes, Santander, Silva y Hazin (2013a) establecieron la madurez sexual mediante características macro y microscópica (ovogénesis y espermatogénesis) y tipo de desove; y Shinozaki-Mendes, Santander, Silva y Hazin (2013b) investigaron la variación estacional de las etapas de maduración gonadal, talla de primera madurez sexual (L50) y fecundidad. Así mismo, en Jamaica Billings y Munro (1974) estudiaron la biología, ecología y bionomía; en Puerto Rico, Sadovy, Figuerola y Román (1989) y en Antigua, Constantine (2008) examinaron la edad y crecimiento; en Yucatán, Domínguez-Viveros y Ávila-Martínez (1986) estudiaron la pesquería. Finalmente, Treinen-Crespo, Villegas-Hernández, Guillén-Hernández, Ruiz-Zárate y González-Salas (2012) realizaron un análisis de la forma del otolito como herramienta para la diferenciación poblacional. Es importante señalar que en Venezuela se han realizado estudios sobre la descripción taxonómica de la especie (Cervigón, 1993) y la proporción sexual, talla mínima de madurez, época de reproducción y fecundidad (Palazón-Fernández, 2007). Debido al desconocimiento de la dinámica poblacional de H. plumieri, sobre todo en los actuales momentos en que se ha convertido en un recurso de alta demanda para el consumo por su carne de excelente calidad (Cervigón, 1993), se determinó estimar su crecimiento y mortalidad, para contribuir a la evaluación y correcto manejo pesquero.
Materiales y métodos
Recolecta de muestras: Los ejemplares de corocoro margariteño Haemulon plumieri fueron obtenidos de las capturas comerciales de la flota artesanal de El Tirano que utilizan nasa hexagonal tipo antillano elaborada con malla de plástico y alambre galvanizado de 5 cm de abertura, sin carnada colocadas en “enyugue” (grupo de tres o más unidas por una cuerda y separadas a una distancia de 15 a 20 m) sobre el fondo marino (González, Eslava, & Guevara, 2006) a una profundidad aproximada de 20 a 40 m. Las faenas de pesca se efectuaron en zonas cercanas al archipiélago Los Frailes, ubicado al noreste de la isla de Margarita, estado Nueva Esparta (11º11’30” N & 63º42’00” W) (Fig. 1).
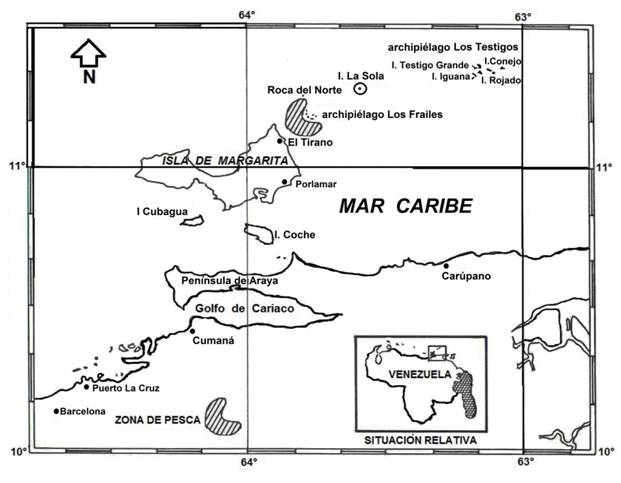
Fig. 1. Fig. 1 Zona de pesca del archipiélago Los Frailes, Venezuela. Modificado de González, Eslava y Guevara (2001). Fishing zone from Los Frailes Archipelago, Venezuela. Modified by González, Eslava y Guevara (2001).
Los muestreos se realizaron semanalmente de enero a diciembre 2014 en el puerto de El Tirano. El número de individuos muestreados mensualmente fluctuó entre 177 y 360. A cada ejemplar se determinó su longitud total (L) en cm, comprendida desde extremo anterior de la boca hasta el extremo posterior de los radios más largos de la aleta caudal, utilizando un ictiómetro marca Wildco modelo 118 con apreciación de 1 mm, y el peso total sin eviscerar (P) en g mediante una balanza electrónica marca Sartorius modelo 1 106 con precisión de 0.01 g y 1 kg de capacidad máxima. La identificación sexual se estableció mediante la observación de las gónadas, según Palazón-Fernández (2007).
Análisis estadístico: Las diferencias estadísticas significativas entre las longitudes por sexo se precisaron con la aplicación de la prueba t-student (Zar, 2009). La relación entre longitud y peso se determinó mediante la ecuación potencial de Ricker (1975): P = a * L b ; donde P es el peso total (g), L es la longitud total (cm), a es la intersección con el eje y, b es la pendiente de la curva que indica la isometría o alometría en el crecimiento. Las constantes a y b fueron estimadas por el método de mínimos cuadrados, previa linealización de los datos mediante una transformación logarítmica (ln). Se aplicó la hipótesis del crecimiento isométrico (Ricker, 1975) mediante la prueba t-student (Walpole, Myers, Myers, & Ye, 2007) con un nivel de significación de α = 0.05.
Parámetros poblacionales: En la estimación de los parámetros poblacionales se utilizaron las longitudes comprendidas entre 14 y 31 cm, debido a que las tallas de 32 a 41 cm estuvieron poco representadas (21 ejemplares). El cálculo de la longitud asintótica (L ∞ ) y el coeficiente de crecimiento o curvatura (k) de la ecuación de crecimiento de von Bertalanffy (1938), se basó en el método indirecto del análisis de la distribución mensual de las frecuencias de longitud de H. plumieri con un rango de 1 cm, utilizando las rutinas del paquete FiSAT (FAO-ICLARM Stock Assessment Tools) (Gayanilo, Sparre, & Pauly, 1996). Se consideró una longitud asintótica preliminar aplicando el Método de Powell (1979) y Wetherall (1986), que permitió determinar el coeficiente de crecimiento mediante la rutina ELEFAN I (Pauly & David, 1981), previa reestructuración de las frecuencias de longitud. Para la optimización de los parámetros de L ∞ y k se empleó el procedimiento de Gulland y Holt (1959) asociado a la misma rutina, previo análisis de la progresión modal utilizando la descomposición de la frecuencia de las distribuciones mensuales de longitud según el método de Bhattacharya (1967). Una vez determinadas las medias de los grupos modales por mes, estas se unieron entre meses para obtener los datos del incremento de crecimiento. Se calculó el to de acuerdo a la fórmula indicada por Pauly (1979): log10(- to) = - 0.3922 - 0.2752 log10L ∞ - 1.038 log10k; donde to es la edad teórica en que el organismo tiene talla cero, L ∞ es la longitud asintótica o longitud máxima promedio y k es el coeficiente de crecimiento o coeficiente de curvatura.
El peso asintótico se estimó mediante la fórmula de Csirke (1980): P ∞ = a * L ∞ b ; donde P ∞ es el peso asintótico o peso máximo promedio. Para el cálculo de la longevidad se utilizó la ecuación de Taylor (1958): A0.95 = to + 2.996 / k donde A0.95 es la edad máxima o tiempo requerido para alcanzar el 95 % de la longitud máxima del pez (L ∞ ). Una vez calculados los parámetros de la ECVB (L ∞ , k y to), se establecieron las curvas de crecimiento ajustadas al modelo de von Bertalanffy (1938) en longitud y peso respectivamente, hasta la edad límite, según las siguientes ecuaciones: L = L∞ [1-e-k (t - to) ]; P = P∞ [1-e -k ( t-to) ]b. Se determinó el índice de desempeño del crecimiento phi prima (Ø′) para comparar los parámetros de crecimiento con los resultados determinados por otros investigadores en diferentes regiones, de acuerdo a la fórmula de Munro & Pauly (1983): Ø′ = log10k + 2 log10L ∞ . Se consideró el criterio de Sparre y Venema (1997), quienes señalaron que el coeficiente de variación (CV) de Ø′ no debe exceder de 4 % para asumir que los resultados son estadísticamente similares.
La mortalidad natural (M) se calculó utilizando la ecuación empírica de Pauly (1980): log10(M) = -0.0066 - 0.279 log10L∞ + 0.6543 log10k + 0.4634 log10T con la temperatura superficial media del agua de 28 °C. La mortalidad total (Z) se calculó mediante el método de la curva de captura linealizada basada en datos de longitud (Pauly, 1983). La mortalidad por pesca (F) se calculó de la diferencia entre Z y M, y la tasa de explotación (E) según Gulland (1971): E = F / Z quien indicó que la tasa de explotación óptima (Eóptima) ocurre cuando E = 0.5/año.
Resultados
Se examinó un total de 2 967 ejemplares (2 084 juveniles y 883 adultos) con longitudes entre 14 y 41 cm (Media = 20.58 cm DE ± 2.42) y pesos que fluctuaron entre 45 y 890 g (Media = 131.22 g DE ± 60.23). El mayor número de individuos se registró en agosto, mientras que el menor en diciembre. La menor longitud se encontró en enero y la mayor en octubre (Tabla 1). No se encontraron diferencias estadísticamente significativas para la longitud entre hembras y machos (ts= 0.093; P > 0.05), por lo que se determinó el crecimiento y la mortalidad para los sexos combinados. En los meses estudiados, excepto febrero, H. plumieri presentó la mayor frecuencia de longitud entre 18 y 23 cm, siendo la moda de 20 cm.
TABLA 1 TABLE 1 Ejemplares recolectados de Haemulon plumieri, enero-diciembre 2014 en el archipiélago Los Frailes, Venezuela Haemulon plumieri specimens collected during January-December 2014 from Los Frailes Archipelago, Venezuela
| Mes | n | Longitud total (cm) | Peso total (g) | ||||
| Min-Max | Media | DE± | Min-Max | Media | DE± | ||
| Enero | 241 | 14-38 | 20.30 | 2.39 | 48-801 | 126.71 | 61.65 |
| Febrero | 252 | 16-37 | 19.87 | 4.53 | 45-800 | 135.59 | 116.62 |
| Marzo | 273 | 16-31 | 19.11 | 1.94 | 57-493 | 101.9 | 41.50 |
| Abril | 234 | 17-26 | 19.33 | 1.29 | 58-240 | 100.78 | 21.98 |
| Mayo | 335 | 16-31 | 20.22 | 2.08 | 60-468 | 123.96 | 50.58 |
| Junio | 204 | 16-27 | 20.62 | 1.56 | 60-332 | 131.26 | 30.94 |
| Julio | 206 | 17-35 | 20.34 | 2.21 | 69-639 | 130.48 | 55.93 |
| Agosto | 360 | 17-32 | 21.17 | 2.12 | 67-343 | 138.49 | 43.66 |
| Septiembre | 249 | 16-26 | 20.13 | 1.76 | 58-244 | 115.27 | 34.66 |
| Octubre | 205 | 16-41 | 21.94 | 3.86 | 72-890 | 161.33 | 127.72 |
| Noviembre | 231 | 16-33 | 22.28 | 2.43 | 61-499 | 159.15 | 65.44 |
| Diciembre | 177 | 16-33 | 21.68 | 2.91 | 64-528 | 149.78 | 72.05 |
| Total | 2 967 | 20.58 | 2.42 | 131.22 | 60.23 | ||
n = número ejemplares examinados; Min = valor mínimo; Max = valor máximo; Media = valor medio; DE = desviación estándar.
n = number of specimens examined; Min = minimum value; Max = maximum value; Media = mean value; DE = standard deviation.
Crecimiento
La relación longitud-peso quedó establecida como P = 1.4×10-3 * L2.99 siendo la constante b igual a 3 (ts = -0.302; P >0.05), demostrando así un crecimiento isométrico (Fig. 2), lo que indica que los ejemplares, tienen un crecimiento individual donde las proporciones geométricas del cuerpo no cambian (crecimiento homogéneo).
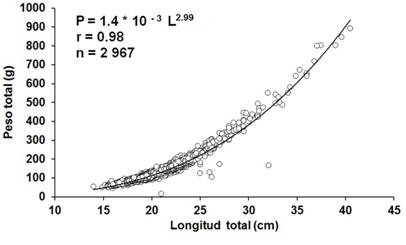
Fig. 2. Fig. 2 Relación longitud total - peso total de Haemulon plumieri del archipiélago Los Frailes, Venezuela de enero a diciembre 2014. Total length - total weight relationship of Haemulon plumieri from Los Frailes Archipelago, Venezuela between January and December 2014.
Los grupos modales por mes no estuvieron bien definidos. Se realizó una primera estimación de la longitud asintótica (L∞) igual a 30.95 cm (Fig. 3A), que fue utilizada como dato de entrada en la rutina ELEFAN I para la estimación de k. Luego de varias corridas, se obtuvo una primera valoración de k = 0.67/año, evidenciándose el inicio de la curva de crecimiento en longitud en mayo. La bondad de ajuste entre la L∞ y el k fue de Rn = 0.271. Posteriormente se aplicó la rutina de Gulland y Holt (1959), la cual proporcionó valores adicionales de L∞ y k, los que fueron descartados por no corresponder a las características biológicas de la especie (Fig. 3B). Se estimó el to= -0.24 años, lo que permitió finalmente determinar la edad límite A0.95 = 4 años. Se obtuvo un peso asintótico de 418.17 g, utilizando las constantes a = 1.4×10-3 y b = 2.99 de la ecuación de la relación longitud-peso y el parámetro preliminar de crecimiento L∞= 30.95 cm.
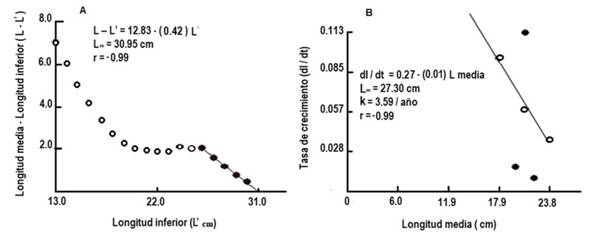
Fig. 3A. Fig. 3A Estimación de la longitud asintótica (L∞) aplicando el modelo de Powell (1979) y Wetherall (1986). B. Estimación de la longitud asintótica (L∞) y el coeficiente de crecimiento (k) aplicando el modelo de Gulland & Holt (1959) de Haemulon plumieri del archipiélago Los Frailes, Venezuela, enero-diciembre 2014. Valores considerados en la regresión lineal en A ( y ( en B. Estimation of Asymptotic length (L∞) applying the Powell (1979) and Wetherall (1986) model. B. Estimation of Asymptotic length (L∞) and growth coefficient kapplying the Gulland & Holt (1959) model of Haemulon plumieri from Los Frailes Archipelago, Venezuela between January and December 2014. Values considered in the linear regression in A ( and ( in B.
La curva de crecimiento en longitud describió una curva de tipo exponencial, acelerado en el primer año de vida y luego disminuyó gradualmente hasta alcanzar la longitud máxima teórica (Fig. 4A). La curva de crecimiento en peso mostró un comportamiento sigmoideo lento en el primer año de vida, luego en forma exponencial hasta los dos años y posteriormente, una disminución paulatina hasta alcanzar su peso asintótico (Fig. 4B). El coeficiente de variación de Ø′ por el método directo (CV = 5.26 %) indica que existe variabilidad de los parámetros de crecimiento de H. plumieri en diferentes regiones, mientras que por el método indirecto (CV = 0.55 %) no existe variación (Tabla 2).
Fig. 4A. Fig. 4A Curva de crecimiento en longitud de Haemulon plumieri según el modelo de von Bertalanffy (1938). B. Curva de crecimiento en peso de Haemulon plumieri según el modelo de von Bertalanffy (1938) del archipiélago Los Frailes, Venezuela de enero a diciembre 2014. Growth curve in length of Haemulon plumieri, according to the von Bertalanffy (1938) model. B. Growth curve in weight of Haemulon plumieri, according to the von Bertalanffy (1938) model from Los Frailes Archipelago, Venezuela between January and December 2014.
TABLA 2 TABLE 2 Parámetros de crecimiento de Haemulon plumieri estimados por diferentes autores Growth parameters of Haemulon plumieri estimated by different authors
| Autor | Región | Sexo | L∞ (cm) | k (año) | Ø′ | A0.95 (años) | |||
| Método directo (otolitos) | |||||||||
| Potts & Manooch (2001) | Sureste de Florida (EUA) | Combinados | 32.70 | 0.19 | 2.30 | 15 | |||
| Norte y sur de Carolina (EUA) | Combinados | 59.10 | 0.08 | 2.44 | 13 | ||||
| Murie & Parkyn (2005) | Costa del Golfo de Florida (EUA) Norte-Central Central | Hembras Machos Hembras Machos | 31.70 34.50 29.50 31.90 | 0.35 0.41 0.30 0.36 | 2.54 2.68 2.41 2.56 | 18 | |||
| Araújo & Martins (2007) | Costa Central de Brasil (Brasil) | Combinados Hembras Machos | 31.22 29.80 33.10 | 0.48 0.59 0.35 | 2.67 2.71 2.58 | 28 | |||
| Constastine (2008) | Antigua (Costa sur oriental) | Combinados | 28.20 | 0.36 | 2.45 | 14 | |||
| Coeficiente de Variación (CV) | 5.26 | ||||||||
| Método indirecto (frecuencia de tallas) | |||||||||
| Billings & Munro (1974) | Arrecife de Port Royal (Jamaica) | Combinados | 42.00 | 0.37 | 2.81 | 8 | |||
| Domínguez-Viveros & Ávila-Martínez (1986) | Banco de Campeche (México) | Combinados | 45.43 | 0.29 | 2.78 | 11 | |||
| Este estudio | Archipiélago Los Frailes (Venezuela) | Combinados | 30.95 | 0.67 | 2.80 | 4 | |||
| Coeficiente de Variación (CV) | 0.55 | ||||||||
L∞ = longitud asintótica; k = coeficiente de crecimiento; Ø′ = índice de desempeño de crecimiento; A0.95 = edad máxima.
L∞ = asymptotic length; k = growth coefficient; Ø′ = growth performance index; A0.95 = max age.
Mortalidad
La mortalidad natural M se estimó en 1.36/año, y la mortalidad total Z = 3.31/año (Fig. 5), en consecuencia, se obtuvo una mortalidad por pesca de F = 1.95/año y la tasa de explotación E = 0.59/año.
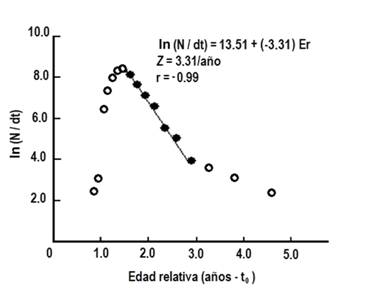
Fig. 5. Fig. 5 Estimación de la mortalidad total (Z) de Haemulon plumieria partir del método de la curva de captura linealizada del archipiélago Los Frailes, Venezuela de enero a diciembre 2014. ( Valores considerados en la regresión lineal. Estimation of total mortality (Z) from a linearized catch curve method of Haemulon plumieri from Los Frailes Archipelago, Venezuela between January and December 2014. ( Values considered in the linear regression.
Discusión
Las longitudes encontradas en el presente trabajo para H. plumieri variaron entre 14 y 41 cm, lo cual difiere con lo determinado por Potts y Manooch (2001) en el norte y sur de Carolina (17.3-51.2 cm), Murie y Parkyn (2005) en la costa oeste de Florida (7-41 cm), Araújo y Martins (2007) en la costa central de Brasil (21.8-37.8 cm), Palazón-Fernández (2007) en la isla de Margarita (18.3-38.9 cm), Constantine (2008) en la costa sur oriental de Antigua (15.9-30.9 cm) y Shinozaki-Mendes et al. (2013a) en el estado de Ceará, Brasil (13.5-34.3 cm). Los disímiles rangos de longitudes observadas en las distintas regiones quizás estén dados por los artes de pesca utilizados en las capturas exploratorias y comerciales, como línea de mano, palangre, artes de enmalle, arrastre y nasa (Holden & Raitt, 1975), y también por las características ecológicas de la zona de pesca. Cervigón (1993) mencionó que el mayor ejemplar observado de H. plumieri en el archipiélago Los Roques fue de 38.5 cm de longitud total, la cual es menor a la longitud total de 41 cm encontrada en el archipiélago Los Frailes utilizando el mismo arte de pesca, lo que hace suponer que las longitudes máximas también están condicionadas a las zonas de pesca.
El coeficiente de regresión b = 2.99 de la relación longitud-peso de H. plumieri, indica un crecimiento isométrico, es decir, existe mayor relación entre la longitud y el peso, por lo que el peso del cuerpo aumenta a un ritmo igual a la longitud (crecimiento homogéneo); diferente al crecimiento alométrico, que a medida que crecen los individuos modifican sus proporciones corporales, como es el caso (b = 2.86) encontrado por Nomura (1965) en el noroeste de Brasil, Billings y Munro (1974) en Jamaica (b = 2.93), Potts y Manooch (2001) en el sureste de Florida (b = 2.73) y Murie y Parkyn (2005) en la costa central de Florida (b = 2.75) y para la región centro-norte (b = 2.88). Sin embargo, Potts y Manooch (2001) reportaron en el norte y sur de Carolina (b = 3.05). En la isla de Margarita, Palazón-Fernández (2007) obtuvo un crecimiento alométrico diferente en adultos (b = 2.84) y juveniles (b = 3.17). Estos resultados indican la variabilidad de la relación longitud-peso en diferentes regiones, posiblemente, por la disparidad en la estructura de tallas de la población, debido al sexo, madurez sexual, épocas del año y alimentación (Tresierra, Culquichicón, & Solano, 2013).
Una explicación de que no se haya presentado una progresión modal clara de la frecuencia de longitudes en el tiempo, es que H. plumieri es una especie tropical que se reproduce durante todo el año, con picos de máxima intensidad en febrero-abril y agosto-octubre (Palazón-Fernández, 2007). La longitud asintótica (L∞) determinada para H. plumieri presentó una considerable variación porque estuvo sujeto a las regiones y estructura de longitudes de las muestras, e.g., por el método directo (otolitos) la mayor fue de 59.1 cm en Carolina (Potts & Manooch, 2001) y la menor de 28.2 cm en Antigua (Constantine, 2008); mientras que por el método indirecto (frecuencia de tallas) la mayor fue de 42 cm en Jamaica (Billings & Munro, 1974) y la menor de 30.95 cm en el presente trabajo.
El coeficiente de crecimiento determinado (k = 0.67/año) indica que esta especie tiene un crecimiento moderado (Sparre & Venema, 1997), siendo su estimación mayor al obtenido por Billings y Munro (1974) en aguas de Jamaica (k = 0.37/año) y al determinado por Potts y Manooch (2001) en el norte y sur de Carolina (k = 0.08/año). Por lo general, los peces tropicales presentan valores más altos de k que los peces de aguas frías por su mayor tasa metabólica (Sparre & Venema, 1997). En el presente trabajo, la fracción adulta > 31 cm no apareció regularmente en las capturas durante el periodo de muestreo, lo cual sería la causa principal de la subestimación de la longitud asintótica (L∞) y la sobreestimación del coeficiente de crecimiento (k). Los valores calculados del índice de crecimiento (Ø’) de H. plumieri presentan amplia diferencia, e.g., el coeficiente de variación de Ø′ determinado por el método directo (CV = 5.26 %) supera el 4 %, lo que indica la existencia de variabilidad del patrón de crecimiento en las diferentes regiones; mientras que por el método indirecto (CV = 0.55 %) se demuestra lo contrario. Estas divergencias del índice de desempeño (Ø′) determinado por el método de los otolitos, puede deberse a problemas en la lectura e interpretación de los anillos de crecimiento que comprometen las estimaciones de la longitud asintótica (L∞) y el coeficiente de crecimiento (k). Por otro lado, Sparre y Venema (1997) señalan que los parámetros de crecimiento dentro de una misma especie, pueden variar de una población a otra, o también cohortes sucesivas pueden crecer de forma diferente según las condiciones ambientales. Algo similar ocurre con la edad máxima teórica calculada de 13 a 28 años por el método de los otolitos y de 8 a 11 años por el método de frecuencia de tallas, particularidad etaria de peces de vida larga, cuando su longitud máxima (L∞) es de 59.10 cm en el norte y sur de Carolina (Potts & Manooch, 2001). Estos resultados difieren con el valor de 4 años obtenido en este trabajo que corresponde a una especie de vida corta.
Las tasas de mortalidad natural (M), por pesca (F) y total (Z) del presente estudio fueron mayores a las estimadas por Domínguez-Viveros y Ávila-Martínez (1986) en el Banco de Campeche, México (M = 1.02/año, F = 0.60/año, Z = 1.6 /año), Potts (2000) en el norte y sur de Carolina (M = 0.30/año, F = 0.23/año, Z = 0.53/año) y sureste de Florida (M = 0.30/año, F = 0.33/año, Z = 0.63/año), y Araújo y Martins (2007) en la costa central de Brasil (M = 0.15/año, F = 0.66/año, Z = 0.21/año). Sin embargo, los valores de mortalidad por pesca y total fueron menores a las determinadas por Billings y Munro (1974) en el arrecife de Port Royal, Jamaica (F = 5.13/año, Z = 6.90/año). Sánchez y Medina (2008) indican que la expresión M es alta cuando los peces tienen crecimiento rápido y habitan ambientes cálidos. Por otra parte, las variaciones en los valores, posiblemente son causados por factores biológicos independientes de la pesca, como enfermedades, depredación, competencia, estrés del desove, inanición y vejez (Sparre & Venema, 1997). Billings y Munro (1974) señalan que la mortalidad natural de los Hemúlidos es causada principalmente por depredación, enfermedad y vejez. La mortalidad por pesca es un parámetro importante porque puede ser controlado y regulado por el hombre; la captura o número de peces muertos por la pesca es uno de los pocos, y a veces el único dato de la población que puede ser medido y observado directamente, y puede ser relacionado posteriormente con el número de ejemplares vivos (Csirke, 1980). Es importante acotar que en las regiones costeras de Venezuela no existe una pesca dirigida al H. plumieri. En este caso, los ejemplares analizados fueron obtenidos con nasa, siendo una de las 51 especies demersales capturadas en el archipiélago Los Frailes, lo que estaría ocasionando un fuerte impacto en la pesca, cuyo efecto directo podría causar cambios en la abundancia y también en la estructura de tallas, con una reducción en las tallas máximas (Sparre & Venema, 1997).
La tasa de explotación (E) estimada de H. plumieri en el presente trabajo (E = 0.59/año) fue mayor a la determinada por Domínguez-Viveros y Ávila-Martínez (1986) en México (E = 0.36/año), Potts (2000) en el norte y sur de Carolina (E = 0.43/año) y sureste de Florida (E = 0.52/año), y Araújo y Martins (2007) en Brasil (E = 0.28/año); pero menor a la estimada por Billings y Munro (1974) en Port Royal, Jamaica (E = 0.74/año). Basados en las tallas mínimas de madurez sexual de 21.3 cm en hembras y 27.1 cm en machos (Palazón-Fernández, 2007) y el alto porcentaje de juveniles (70.24 %) en la composición de las capturas, se infiere que H. plumieri del archipiélago Los Frailes está siendo sobre explotada. Pauly (1983) explica que la sobrepesca puede ocurrir cuando los juveniles de las especies comerciales pasan a ser disponibles a la pesquería, es decir, los reclutas son capturados antes de alcanzar la talla necesaria. Teniendo en cuenta las características multiespecíficas de la pesquería artesanal y los antecedentes similares con Orthoristis ruber (Guerrieri, Eslava, González, & Guevara, 2015) y Canthidermis sufflamen (Alarcón, Eslava, González, & Guevara, 2017), se sugiere aumentar la abertura de malla y disminuir el número de nasas permitidas por pescador que faena en el archipiélago Los Frailes.

