











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkA nivel internacional se ha dicho que la pesca efectuada con redes de arrastre ha sido objeto de señalamientos como causante de alteraciones químicas y biológicas en los fondos marinos (Dernie, Kaiser, Richardson, & Warwick, 2003; Nieto, 2010; Pérez, 2011). Este tipo de pesca es selectiva y ha sido señalada por capturar especies de peces, crustáceos y moluscos no objetivos de la pesquería, con potenciales efectos adversos a sus poblaciones (Thrush & Dayton, 2002; Rábago, López, Valdez, & Nevárez, 2011). Esto provoca cambios en el funcionamiento del ecosistema (Caddy, 2007) y afectación sobre los depredadores (Myers & Worm, 2003). Incluso se ha sugerido que puede colapsar poblaciones de especies de baja tasa de natalidad, como sucede con tiburones y rayas (Márquez, 2002; Morales, 2011).
El Golfo de California (GC) es uno de los ecosistemas marinos de mayor productividad en el mundo y con gran diversidad de especies asociado a la estabilidad del ecosistema, además de la gran variedad topográfica y bati-métrica, que proporciona un mayor número de hábitats para peces y otros organismos marinos (López, Hernández, Herrera, Rábago, & Morales, 2007).
El Norte del Golfo de California (NGC) posee al menos 7 200 km2 (Nelson Ramírez, Arenas, Jacquemine, & Prado, 1980) desde el Delta del Río Colorado hacia la región de las grandes islas: Isla Tiburón y Ángel de la Guardia, con una profundidad promedio de 200 m, de aguas ricas en nutrientes y alta productividad en todo el año (Lluch & Arias, 2000). Es considerado también como uno de los ecosistemas marinos más ricos del mundo (Cudney & Turk, 1998), altamente dinámico y cambiante, lo que hace posible sostener grandes cadenas alimenticias debido al gran potencial alimenticio autotrófico. Más al norte se encuentra la Reserva de la Biosfera Alto Golfo de California y Delta del Río Colorado (RB) decretada en junio de 1993 (Diario Oficial de la Federación, 1993), de la cual 82 % constituye la zona de amortiguamiento, lugar donde existe una gran actividad pesquera multiespecífica.
En el NGC la pesquería de camarones peneidos Litopenaeus stylirostris y Farfantepe-naeus californiensis , genera una fuente importante de ingresos en la comunidad, cerca de 77% de la población vive de la pesca (López, Herrera, Rodríguez, & Hernández, 2010). Esta actividad se desarrolla entre los 9 a 90 m de profundidad, donde opera la flota de arrastre y donde se localiza gran variedad de especies que forman parte de la fauna acompañante del camarón (FAC) compuesta de peces, crustáceos y moluscos (López, Rodríguez, Hernández, & Herrera, 2011).
Con respecto a la comunidad íctica del GC, sobresalen los listados sistemáticos de peces en sistemas lagunares (Rodríguez, Abitia, Galván, & Chávez, 1994), y en otros estudios, se incluyen aspectos de estructura, diversidad, abundancia y variaciones espacio-temporales en comunidades de peces (López et al., 2010). Si bien es cierto, los estudios basados en índices ecológicos nos aproximan a conocer la salud de las comunidades, Jennings y Reynolds (2000) sugieren que la utilización de estos no ha sido suficiente para evaluar impactos provocados por las redes de arrastre en las comunidades béntico demersales, debido a que no han mostrado cambios significativos. Esto hace necesario desarrollar o aplicar otros métodos tales como el uso de las curvas k-dominancia, como una herramienta más recomendable para identificar los niveles de estrés en que se encuentran las comunidades de peces (Jouffre & Inejih, 2005).
A pesar de la importancia pesquera y ecológica del NGC, existe escasa información sobre la ecología de comunidades de peces sujetas a la pesca de arrastre. En cuanto a análisis con agregados comunitarios como curvas k dominancia (Lambshead, Plat, & Shaud, 1983), y aplicaciones de la curva comparativa de Abundancia-Biomasa ABC (Clarke & Warwick, 2001), este estudio representa la primera aproximación para detectar grados de alteración en la comunidad de peces (predicha como cambios en la comunidad por la intensificación de la pesca), donde especies pequeñas de rápido crecimiento y oportunistas se convierten en dominantes, siguiendo a Yemane, Field y Leslie (2005). Por todo anterior, el objetivo de esta investigación fue analizar el grado de estrés, de la comunidad de peces asociados a los fondos blandos, provocados por la pesca de arrastre de camarón en una zona megadiversa como lo es el NGC, con base en la utilización de estos análisis.
Materiales y Métodos
La composición de la FAC de Puerto Peñasco, Sonora, fue evaluada en 119 muestras obtenidas a bordo de la flota camaronera, durante las temporadas de pesca 2009-2010 y 2010-2011. Para esto, se integró un programa de cuatro observadores a bordo de la flota comercial, y adicionalmente, se contó con información de un crucero de veda del camarón recopilada en la misma zona, por el Instituto Nacional de Pesca (INAPESCA) en agosto 2011.
Los 119 arrastres de los 14 viajes fueron realizados en los diferentes caladeros de pesca del NGC (Fig. 1). El tiempo de arrastre varió de una a tres horas, con una velocidad promedio de 5.5 km/hora y profundidades entre 9-90 m. Se utilizaron red de arrastre de relinga superior a 30.48 m, relinga inferior a 38.1 m, el peso de la cadena espantadora fue variable 50-75 kg. La parte delantera con luz de malla de 140-160 mm, cuerpo con malla de 120 mm, la antebolsa de malla 100 mm y bolsa con luz de malla de 90 mm.
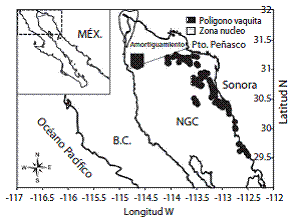
Fig. 1: Área de pesca y recolecta de muestras en el norte Golfo de California durante 2010-2011. Fig. 1: Fishing and sample collection study area in the Northern Gulf of California from 2010-2011.
La duración de los viajes fue de 4 a 15 días, en la bitácora de viaje se registraron los datos propios del lance (fecha, lance, profundidad, área de pesca, velocidad de arrastre, hora inicial-hora final, posición inicial y final del lance, peso y composición de la captura objetivo, captura total de FAC).
Para los análisis, se obtuvieron muestras al azar de aproximadamente 20 kg de la FAC. El tamaño de muestra se determinó siguiendo los criterios estándar (Box, Stuart, & Hunter, 2008). La muestra fue etiquetada y guardada en bolsas de plástico y conservadas en la bodega del barco y posteriormente en bodegas frigoríficas de plantas pesqueras hasta su traslado al laboratorio de Pesquerías del Campus Guay-mas del CIBNOR-Unidad Sonora.
Para la identificación de las especies, se utilizaron las claves de Jordan y Ever-mann (1893-1900), Norman (1934), Curran (1942), Zahuranec (1967), Miller y Lea (1972), Castro-Aguirre (1978), Compagno (1984), Fischer et al (1995), y con bibliografía especializada para algunos grupos: Castro-Aguirre y Espinosa-Pérez (1996) en rayas, McPhail (1958) para la familia Sciaenidae, Ginsburg (1958) en Pleuronectiformes, Rosenblatt y Johnson (1974) para Diplectrum, Walker y Rosenblatt (1988) para Porichthys.
Los nombres comunes fueron tomados de Love, Yoklavich y Thorsteinson (2002) y Robertson y Allen (2002).
Composición de especies: Se integró el listado de las 30 especies de peces bento-demer-sales más frecuentemente capturadas por la red de arrastre de la flota camaronera del norte del Golfo de California en el periodo estudiado. Esta información y sus abundancias, sirvió para obtener la estructura de las comunidades a través de los índices ecológicos aplicados.
Índice de Valor Biológico: {XE (tm)6.2.4 Índice de valor biológico (IVB)"}. El índice de Valor Biológico (IVB) de Sanders (1960) descrito Loya y Escofet (1990), tiene la ventaja de combinar la propiedad de abundancia relativa con la constancia de especies y es utilizada para estimar las especies de mayor importancia biológica en la FAC; así las especies que sean más constantes durante los muestreos serán más importantes que aquellas que presenten abundancias altas en menor número de muestreos, este índice se expresa de la siguiente manera

donde: IVB es el Índice de valor biológico, i corresponde a la abundancia relativa de cada especie y j a las estaciones de recolección. Para medir los índices ecológicos de diversidad y riqueza específica se utilizó:
Diversidad de Shannon-Wiener (H'): se expresa de la siguiente manera:

donde: H' = Índice de diversidad de Shannon y Weaver (1963), P= Cociente del número de organismos de la especie i entre el número total de la muestra, ni/NT.
Equitabilidad o Uniformidad J' (Pielou, 1966): J' = H'/ln S donde S = Se considera el número total de especies (S) en una muestra o localidad.
Curvas de abundancia-biomasa (ABC):
Las curvas de abundancia-biomasa (ABC) han sido utilizadas para determinar el nivel de alteración o grado de estrés ecológico de las comunidades (Warwick & Clarke, 2001). Se comparó la disposición de las curvas en un mismo gráfico de abundancia y biomasa de las especies en forma de curvas tipo ^-dominancia (Lambshead et al., 1983; Warwick, 1986). El modelo asume que en condiciones estables, las especies estrategas tipo k, de gran tamaño, longevas, ritmo de crecimiento lento dominan por su biomasa. A medida que en la comunidad se incremente el grado de estrés por perturbaciones de diversa índole, se verá afectada la biomasa de estas especies, tendiendo a disminuir. Por otra parte, empezarán a dominar en abundancia numérica las especies estrategas tipo r (de tallas pequeñas, de rápido crecimiento y vida corta) (Warwick, 1986; Clarke & Warwick, 2001; Yemane et al., 2005). Las especies se ordenan en el eje X (en escala logarítmica) en términos de su porcentaje acumulado de abundancia o biomasa, el estadístico W (Warwick & Clarke, 1994) representa el área de diferencia entre las dos curvas.

donde: S describe el cambio en el número total de especies, Ai es la abundancia total en cada muestra i y Bi es la biomasa total en cada muestra i. Este modelo propone que la curva de bio-masa debe encontrarse por encima de la curva de abundancia (W > 0); para ser considerada un área sin estrés, moderadamente estresada cuando coincidan las dos curvas con valores próximos a cero de (W~0) y fuertemente estresado, cuando la curva de biomasa quede por debajo de la curva de abundancia (W < 0) (Warwick, 1986). Los valores del estadístico W fluctúan entre -1 a 1. De acuerdo al supuesto del modelo, cuando el valor del estadístico W es negativo representa indicios de perturbación, lo que reflejará grado de estrés, y cuando el valor del estadístico es positivo, predice que no existe perturbación, y por lo tanto, la comunidad no presenta grado de estrés.
Para los cálculos de los índices de diversidad se utilizó el programa estadístico PRIMER v 6 (Clarke & Gorley, 2001).
Resultados
Composición de especies: Se obtuvo un total de 32 757 individuos, pertenecientes a dos clases, 26 órdenes, 55 familias, 93 géneros y 148 especies; los peces representaron 70.7% de la FAC (el resto estaba integrado por crustáceos, moluscos y otros). El orden Perciformes fue el más diverso con 18 familias, 25 géneros y 43 especies. Las familias mejor representadas en número de especies fueron: Scianidae con 21, Paralichthydae con 11 y las familias Haemuli-dae y Serranidae con 10 especies cada una. Las especies más abundantes de mayor constancia y frecuencia de ocurrencia (FO), fueron: Pori-chthys analis (Hubbs & Schultz, 1939), Diplec-trum macropoma (Günther, 1864), Urobatis halleri (Cooper, 1863), Pomadasyspanamensis (Steindachner, 1875), Micropogonias megalops (Gilbert, 1890) y Synodus scituliceps (Jordan & Gilbert, 1882). Estas fueron parte de las 15 especies en orden de importancia que aportaron el 72 % de la captura total (Cuadro 1), que a su vez formaron parte de las 30 especies más abundantes y frecuentes que aportaron 90 % de la abundancia relativa acumulada.
Índice del valor biológico (IVB): La dominancia numérica estimada a través del IVB mostró que la mayoría de las especies encontradas fueron de hábitat demersal, asociados a fondos blandos (Cuadro 1), Eucinos-tomus dowii (Gill, 1863) fue la especie más importante en el puntaje de IVB = 480.25 con una abundancia relativa AR = 5.00 % y frecuencia de ocurrencia FO % = 36.97 sobrepasando las 15 primeras especies en orden de abundancia; mientras que P. analis, fue la segunda en importancia, obteniendo IVB = 470 en 74 lances y ocupando la tercera posición en FO % = 62.18 pero primer lugar en AR = 12.47 %. La manta redonda U. halleri en tercera posición con IVB = 433.5 pero segundo lugar en FO % = 67.22 % y AR = 6.85 %, S. scituliceps (Jordan & Gilbert, 1882) ocupó el cuarto lugar con IVB = 396.5 con FO = 75 lances con FO % = 63.02 % y una AR = 5.02 %. En quinta posición según el IVB = 393.7, AR = 6.10 % y primer lugar de FO % = 68.91 %, estuvo P. panamensis (Steindachner, 1876) constante en 82 muestras. M. megalops (Gilbert, 1890J presenta una alta abundancia numérica, siendo también de las más constantes, ocupa la octava posición en el IVB = 307.25 con frecuencia de ocurrencia de 68 lances, FO % = 57.14 % y AR = 5.17 % (Cuadro 1).
Cuadro 1: Especies que representaron 90% de la abundancia relativa acumulada e índice de valor biológico IVB en el norte del Golfo de California durante 2010-2011
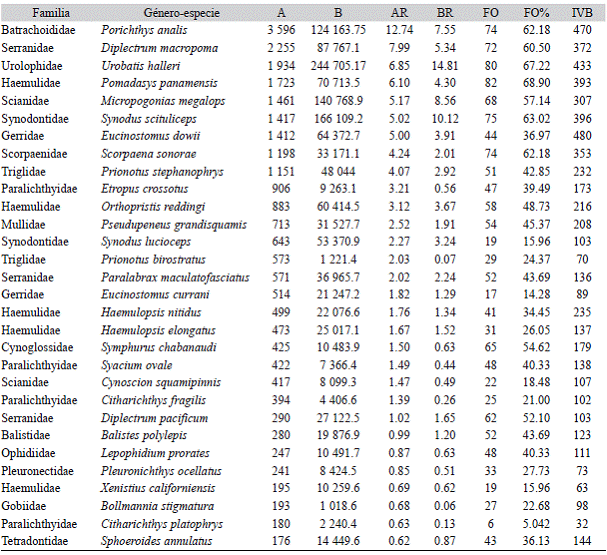
Diversidad: Los valores de los índices ecológicos mostraron una comunidad muy diversa para la región del NGC, así mismo variación entre los meses de captura (Fig. 2). Los valores mensuales mostraron promedio H' = 3.05 (2.72 > H' < 3.25) bits/ind, con el valor más alto de H' = 3.25 ind/bit en octubre 2010, y el valor más bajo en febrero 2010 y enero 2011, con H' = 2.72 ind/bit, seguida de marzo 2010 con H' = 2.92 ind/bit. En el resto de los meses se encontraron valores por encima de 3. El índice de equitatividad J' = 0.71 mostró una comunidad dominada por algunas especies y no se observaron cambios marcados (0.66 < J' > 0.81) (Fig. 2). En general, los índices ecológicos presentan tendencia a decrecer conforme avanzó la temporada de pesca.
Curvas comparativas abundancia-bio-masa ABC: De acuerdo a los resultados positivos basados en el valor del estadístico W, obtenido a través de la comparación de las curvas abundancia-biomasa durante los meses de la temporada de pesca 2010: febrero, octubre y noviembre y agosto 2011, la comunidad no presentó grado de estrés (Fig. 3). Sin embargo, en la misma temporada 2010 durante los meses de marzo y septiembre, el estadístico W mostró un valor negativo, al igual que en enero 2011, por lo que, estos valores negativos identificaron periodos con un grado de estrés de nivel moderado. Confirmado por la comparación entre curvas de abundancia y biomasa (Fig. 4), es decir, la abundancia está por encima de la biomasa debido al efecto provocado por perturbaciones de tipo antropogénico y/o natural. Mientras que en los meses de febrero, octubre y noviembre de la temporada 2010 y agosto 2011 la curva comparativa de la biomasa se encuentra por arriba de la curva de la abundancia, que de acuerdo a los supuestos del método ABC, para estos meses representa una comunidad de peces en buen estado.
Discusión
El número de especies observadas en este estudio se considera robusto, debido al tipo e intensidad de muestras analizadas durante el periodo de pesca. La temporada de pesca de arrastre para camarón en embarcaciones mayores inicia a mediados de septiembre y finaliza en marzo, existiendo un periodo de reposo de seis meses. Este periodo de no actividad pesquera, ayuda en la recuperación de los fondos sometidos al arrastre en los caladeros de pesca. Cabe mencionar que durante la temporada de pesca, varios industriales dueños de las embarcaciones cesan actividades para determinar una "autoveda" en función del decaimiento de las capturas y del precio del camarón en el mercado internacional (López et al., 2010). Aunque, si bien es por medidas económicas, esta determinación se suma a la recuperación natural del sistema ante posibles afectaciones antropogénicas en las comunidades de los fondos blandos. Considerando que, el periodo de veda establecido para esta actividad inicia a principios de abril y finaliza a mediados de septiembre. Adicionado a que solo una fracción de la comunidad de peces demersales es explotada en los caladeros de pesca con fondos blandos muy cercanos a la costa, entre 25-40 m. Lo cual, quizá explique una parte de los resultados, es decir, una comunidad con nivel de estrés de tipo moderado. En este estudio participaron 13 embarcaciones en diferentes tiempos, las muestras que fueron analizadas se obtenían de los caladeros de pesca del NGC, fuera del área de amortiguamiento de la RB, debido a la exclusión de las embarcaciones mayores en el área protegida. El estudio registró 148 especies de peces, mayor a lo encontrado por Nava (1994) quien reportó 76 especies de peces provenientes de cruceros comerciales para la misma área de pesca y con lances dentro de lo que hoy es zona de amortiguamiento dentro de la RB. Aunque en dicho estudio, los organismos fueron registrados y agrupados a nivel de familia y género, por lo que hay una pérdida de información a nivel de especies y se descarta comparar en su totalidad.


Fig. 4: Comparison between Abundance-Biomass ABC curves in the Northern Gulf of California from 2010-2011 (black line biomass, dotted line abundance).
Hasting y Findley (2007), registraron 260 especies de peces provenientes de varias fuentes realizadas en zonas libres para la pesca y en la zona de amortiguamiento de la RB, por lo que una gran proporción de especies no fueron observadas en este estudio, debido a la restricción de zonas de pesca para las embarcaciones mayores.
La riqueza fue mayor a la registrada por Calderón (2011) en el norte Golfo de California, donde enlista 134 especies de peces (70 % de las especies totales de la FAC), lo que coincide con el presente estudio. Cabe mencionar que en los resultados de su estudio también fue considerada el área de amortiguamiento de la RB.
Por lo anterior, se cuenta con registros de las mismas especies (Nava 1994) antes del decreto de la reserva (1995) y en la actualidad, lo que permite afirmar que las especies registradas en 1993-1994 siguen siendo las mismas especies componentes de la FAC en el NGC. Especies como S. scituliceps, U. halleri y P. panamensis fueron registradas por Nava (1994) y son especies consideradas de amplia distribución latitudinal y batimétrica del Pacífico Oriental Tropical (POT) (Robertson & Allen, 2002; Castro, Balart, & Arvizu, 1995). Estas especies continúan formando parte de las 30 especies más abundantes de la FAC dentro del Golfo de California (Pérez & Findley, 1985; Valenzuela, Gracia, & Herrera, 1988; López et al., 2007; Madrid, Amezcua, & Morales, 2007; Rábago et al., 2011). Existen además de las especies anteriores, otras muy abundantes que se encuentran restringidas al NGC, entre las que están M. megalops, S. sonorae y P. analis (Nava, 1994; Cudney & Turk, 1998).
La amplia distribución de algunas de las especies de peces presentes se debe a su adaptación biológica a diversos hábitats y a condiciones ambientales en la zona costera del Golfo de California (Hernández & Solís, 2005; López et al., 2010) considerado como uno de los ecosistemas marinos más ricos del mundo (Cudney & Turk, 1998), por lo que sostiene grandes cadenas alimenticias derivado del gran potencial alimenticio autotrófico. Lo anterior se reflejó en los índices de diversidad de H' = 3.25 bits/ind en la estructura de la comunidad de peces demersales, similares a los valores H' = 3.39 bits/ind reportados en el estudio de Calderón (2011). Aunque en otras áreas del sur del Golfo de California, en un estudio reciente, particularmente en el sur de Sinaloa y norte de Nayarit (Nieto, Zetina, Arreguín, Palacios, & Jordan, 2013) se reportaron valores de H' = 4.1 bits/ind muy superiores a nuestros resultados, y a lo reportado en el estudio de Godínez y González (1999) sobre macroinvertebrados bentó-nicos en las costas de Jalisco y Colima (H' = 2.39) bits/ind en experimentos efectuados en fondos blandos.
Por otro lado, el análisis comparativo entre curvas abundancia biomasa sugiere la presencia de especies pequeñas dominantes, como: P. analis, U. halleri, P. panamensis, Diplectrum macropoma, S. sonorae, Syacium ovale y Prio-notus stephanophrys las cuales explican las diferencias entre las curvas ABC dentro de la comunidad (Clarke & Warwick, 2001), lo que coincide con los resultados en el estadístico W con valores negativos durante los meses de marzo (W = -0.022) y septiembre 2010 (W = -0.02), y enero 2011 (W = -0.042), al mostrar un grado moderado de estrés ecológico. El origen de dichos cambios aún no tiene concenso entre los autores: algunos señalan que podrían ser derivados de las perturbaciones generadas por los efectos de la pesca de arrastre (Kaiser, Collie, Hall, Jennings, & Poiner, 2002; Godí-nez, 2003), porque es considerada la actividad de mayor impacto sobre fondos marinos; esta provoca no sólo mortalidad directa de especies no objetivo, sino también cambios en la diversidad de especies, con efectos indirectos sobre la red trófica por pérdida de depredadores, y destrucción y modificación de hábitat, así como resuspensión de material de fondo provocada por el arrastre continuo (Kaiser et al., 2002; Thrush & Dayton, 2002). En los meses donde el nivel de estrés es bajo, Jouffre y Inejih (2005) mencionan en su estudio que la fuerte resistencia del ecosistema puede ser lo que causa esta discrepancia. Aunque actualmente los investigadores no llegan a un acuerdo respecto al valor negativo del estadístico W para definir una comunidad sobreexplotada (Jouffre & Inejih, 2005), el método logra identificar cambios temporales a escala estacional en las comunidades ícticas (Pérez, 2011).
No obstante lo anterior, es necesario hacer algunos alcances teóricos respecto a las curvas ABC. En primer lugar, pese a que son una herramienta bastante utilizada en la literatura, éstas han sido objeto de una serie de críticas inicialmente por Beukema (1988) quien encontró inconsistencias del método verificadas cuando se presenta una gran dominancia numérica ecológica de especies o formas de pequeño tamaño relativo y de vida corta relacionadas con periodos de reclutamiento. En su estudio sobre contaminación en sedimentos marinos, Burd, Nemee y Brinkhust (1990) encontraron dos limitantes en el método curvas ABC: por un lado asumen que el conjunto faunístico analizado estaba inicialmente en una condición estable o de equilibrio, condición que raramente es efectiva o real en hábitats que son controlados ambientalmente, y la otra limitante refiere a que las estaciones moderadamente contaminadas, muestran análisis muy interesantes, pero con resultados ambiguos.
Godínez (2003) menciona que los cambios espaciales como la batimetría, también pueden ser considerados como perturbación, y que podría ser erróneo concluir que la pesca es la que produce el efecto negativo, en donde la dominancia de especies pequeñas puede interpretarse equivocadamente como un disturbio en la comunidad. Thrush, Hewit, Gibbs, Lundquist y Norkko (2006) sugieren que la comunidad puede responder a niveles de estrés de acuerdo a la intensidad del disturbio, que podrían estar siendo provocados por factores independientes de la pesca, como cambios estacionales en el reclutamiento, o cambios en los ciclos de vida de las especies. Sin embargo, en el estudio de Nieto (2010) sobre la aplicación del método ABC, llega a la conclusión que la pesca de arrastre no fue altamente significativa para concluir que el nivel de deterioro sobre las comunidades de fondos blandos sea atribuible a la pesca.
Aunque en el trabajo de Pérez (2011) que comparó área de arrastre con otra de no perturbación, particularmente para macroinvertebra-dos de la FAC concluye que las curvas ABC detectaron un alto grado de estrés en las zonas donde operó la flota de arrastre camaronera, ya que la abundancia estuvo siempre por encima de la biomasa, parcialmente diferente a lo encontrado en el NGC.
De lo anterior se desprende que, si bien los resultados de las curvas comparativas ABC denotan comunidades en buen estado y moderadamente alteradas, lo que coincide con los índices bioecológicos, éstos deben realizarse continuamente como sugirieron Aderlini y Wear (1992) para suministrar una evaluación más exacta ante posibles disturbios físicos, biológicos o de otra naturaleza. Así mismo, deberán ser combinados con información ambiental y esfuerzo pesquero, para poder asociar a uno u otro como causa principal, sobre todo considerando lo altamente dinámico que es el NGC (Kurczyn, Beier, Lavin, & Chaigneau, 2012).
Así mismo, la carencia de estudios similares para el área de estudio dificulta la comparación de los resultados aquí presentados, limitando la deducción sobre la riqueza y la abundancia de especies. Por otro lado no se identifican cambios en la composición y bioma-sas de las especies más abundantes y frecuentes anteriormente registradas. Sin embargo, esas 30 especies siguen soportando presión pesquera principalmente en los estratos de profundidad 25-40 m, por lo que es necesario realizar estudios directos sobre estas poblaciones.
De acuerdo a la pesca de arrastre efectuada por barcos camaroneros en el Norte Golfo de California, como factor detonante para evidenciar cambios en la estructura de la comunidad de peces y en la composición íctica estimadas por el método curvas comparativas abundancia biomasa ABC a través de su estadístico W, mismos que mostraron valores negativos, se presentó estrés moderado en la comunidad, aunque ambiguo y no explicado totalmente por la actividad de pesca. Las variaciones en la diversidad y abundancias son explicadas por el patrón espacial y temporal de las especies, asociada a la estabilidad y resiliencia del ecosistema.
Los resultados en general fueron explicados por el comportamiento que sigue el periodo de captura y la forma en que operan las embarcaciones.
Es crucial estimar la composición de las especies que forman parte de la (FAC), para identificar aquellas especies que están siendo afectadas en su contribución a los atributos comunitarios como la riqueza, diversidad y abundancias relativas debido a que se encuentran ligados directamente con el funcionamiento del ecosistema.
Es necesario continuar con los análisis espacio temporales entre los diferentes caladeros de pesca del área de estudio, con base en la información de cruceros de investigación y viajes comerciales, hasta lograr una serie robusta de listados ictiofaunísticos, para su manejo, y poder detectar niveles de perturbación en combinación con otros métodos de análisis multi-factoriales para la evaluación de comunidades biológicas, que permitan evidenciar cambios en la estructura comunitaria.
Se recomienda efectuar un análisis de esfuerzo pesquero de las especies comerciales y relacionarlo con información biológica de las poblaciones de peces sujetas a la pesca objetivo, integrando estudios de dinámica pobla-cional para identificar cambios en las medidas de longitud, talla de primera madurez sexual, patrón de reclutamiento, etc.

