











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkClams of the genus Donax are worldwide the dominating group of the invertebrate community on sandy beaches; of the 64 species that have been described until now, about 75 % are to be found in tropical waters, around 22 % lives in temperate areas and a very small amount inhabits coastal upwelling regions (McLach-lan & Brown, 2006). As primary consumers, donacids provide a significant abundance and biomass to the ecosystem, being also important for recreational and/or commercial fisheries in many countries (McLachlan et al., 1996).
Donax striatus Linnaeus, 1767, lives in the intertidal zone of sandy beaches of the Caribbean and the South-eastern Atlantic Ocean from Gibara Bay in Cuba to North-eastern Brazil (Wade, 1967). Despite its large distribution range and important role for nature and human (McLachlan et al., 1996) only a few studies concerning this species have been carried out by now. Wade (1967) suggested that variations in shell colour and morphometric is associated with ecological factors such as beach characteristics, food availability and population density. The morphology of the spermatozoa and aspects about spermatogenesis were described by Matos, Matos, Corral, and Azebedo (1995). The structure of a D. striatus population in Gibara (Northeastern Cuba) was analysed by Ocaña, Fernández, Silva, González, and García (2010). Furthermore, merely two theses from Venezuela (Farache, 1980; Pigallet de Mahieu, 1984) and two published studies from Brazil (Rocha-Barreira, Batista, Monteiro, & Franklin-Junior, 2002) and Venezuela (Delgado, Reverol, Godoy, & Severeyn, 2003) are available dealing with data of population abundance, structure and growth of D. striatus. Besides these studies, biological information of D. striatus is scarce and there is no report on secondary production of this species.
A population of D. striatus at Las Balsas beach has been subjected to a recreational exploitation during many decades according local people. Therefore the aim of the present investigation is to estimate the growth parameters and secondary production of D. striatus from Las Balsas beach in order to provide information for an optimized management of this resource.
Materials and Methods
Study area: Las Balsas is a sandy beach located towards the South part of Gibara Bay in Holguín province, at North-eastern Cuba (21°05'39" N - 76°07'55" W). The beach extends over 1 600 m in length, the mean width of the beach face is 8.8 m and the slope is less than 6 %. The sediment is of terrestrial origin and medium grain size. The tidal cycle in the bay is semidiurnal with a maximum tidal range of 0.6 m. The beach is located between two estuaries of the rivers Gibara at the East and Cacoyugüin at the West, and affected by freshwater seepage, which increases the turbidity of the sea water remarkable during the rainy season (June-October). The sublittoral zone is shallow with a mean depth of 0.5 m to a distance of 150 m from the shoreline.
Sampling: Sampling was carried out monthly from February 2008 to November 2009 during low tide according to a stratified design (Schoeman, Wheeler, & Wait, 2003; Ocana et al., 2010). Three fixed stations were selected at 300 m intervals along the beach. At each station, five strata were set up perpendicular to the shoreline from the low water mark to the drift line. Distance between consecutive strata varied 1.5-2 m depending on the beach width. On each strata of each station, three replicated samples were taken at 1 m intervals by means of a PVC corer of 0.025 m2 area and 30 cm height, introduced in the sand to 20 cm depth (Sample Unit, SU). A total of 45 SU (three stations x five strata x three replicates) were taken each month. The sediment was then sieved on a 1 mm mesh. Collected individuals of D. striatus were stored in a plastic bag and kept on ice. In the laboratory, shell length (L) of individuals was measured to the nearest 0.1 mm, using a digital calliper. Individuals were grouped in 1 mm size class intervals to obtain monthly length frequencies histograms. A sub-sample of 390 specimens, covering all size classes, was used to determine length-weight relationships, first measuring the total mass with shell (TM, g) in a 1 mg resolution scale, thereafter removing soft tissues to determine wet mass (WM, g) after blotting on filter paper.
Growth, longevity and mortality: A series of 20 length-frequency distributions were used for growth estimations, to which a general von Bertalanffy growth function (VBGF) (von Bertalanffy, 1938) was fitted throughout the ELEFAN routine of the FiSAT II program package (Gayanilo, Sparre, & Pauly, 2005): L t=Lx [1-e(-K(t-to)], where L t is L (mm) at time t, L x is the asymptotic maximum anterior-posterior shell length, K is the growth coefficient and to is the theoretical age at zero length.
Preliminary estimates of L x and K, were used to identify the VBGF that best fits the monthly size-frequency data, using the Rn value as a criterion of fit, using response surface analysis of ELEFAN I.
To compare growth parameters with other Donax species the growth performance index (Φ´) (Pauly & Munro, 1984) Φ´=2log(L∞)+log(K) and the overall growth performance index (OGP) of the length-weight relationship (Pauly, 1979) OGP=log, were calculated.
The theoretical life span tmax was estimated by the inverse von Bertalanffy growth equation (Taylor 1958): tmax=[lnL95%-ln(L∞-L95%)]/K, where L95% represents 95 % of the maximum L recorded during field sampling. Mortality rate (Z) of the population was estimated using the exponential extinction model, using the length-converted catch curve of the cumulative data of samplings: Nt=Nt-1(e-Z*Δt), where Nt is the number of individuals in time t and Z is the curvature parameter of the equation.
Biomass and production: Length-weight relationships, using TM (g) and WM (g), were determined by nonlinear regression analysis using the exponential equation: TM, WM = a*Lb, where a and b are constants. Monthly abundance (ind./m2, mean ± standard error) was estimated extrapolating the number of clams collected in each of the 45 SU. Total annual production P (g/m2.yr), considering TM and WM, was computed using all pooled samples by the mass specific growth rate method (Crisp, 1984) from the data of abundance, size-class frequencies, VBGF parameters and length-weight relationships: P = YN*M*G,, where N i is mean abundance (ind./m2) in length class i, M i is the mean individual weight in length class i and G i is the mass-specific growth rate: G % = ^*K*((Lw/Ll.)-1)/|yr], where b is the exponent of length-weight relation an L i is the mean length of individuals at size class i. In addition, mean annual biomass was estimated by: B = HN*M i and annual renewal rates (P/B) of the population were calculated from total annual production and mean annual biomass. Logistical issues impeded me to determine ash-free dry mass (AFDM), so to compare my results with other Donax populations, annual renewal rates were converted to ash-free dry mass (AFDM) using the conversion factor AFDM/WM = 5.8 % (Ricciardi & Bourget, 1998).
Results
Growth, longevity and mortality: A total of 5 471 specimens were collected during the sampling period. The smallest recorded individual measured L= 2.7 mm and the largest L= 33.3 mm. Growth parameters, using ELEFAN I routine best fits (Rn= 0.15), were as follows: L m = 36.1 mm, K= 0.8/yr and t0 = 0.2/yr; these parameters were used to construct the growth curve of D. striatus population at Las Balsas beach (Fig. 1). The growth performance resulted in values of 0'= 3.02 and OGP= 4.57. The estimated theoretical life span was 2.4 yrs.
The instantaneous mortality rate was 3.07/ yr calculated from the length-converted catch curve for the study period; the regression equation is as follows: ln{NIM)= 10.72-3.07/ (R 2= 0.98). In 2008 mortality was 3.30/yr (R2 = 0.97), and in 2009 the estimated mortality rate was 2.85/yr (R2= 0.98).
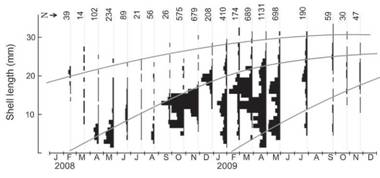
Fig. 1: Growth curves (grey lines) of D. striatus estimated by the FISAT II program from monthly length-frequency data (black histograms) for the period February 2008 to November 2009.
Biomass and production: In order to estimate the production of the Cuban D. striatus population, b values of the length-weight relationships were 3.26 and 3.18 for TM and WT, respectively (Fig. 2).
In 2008, abundance of D. striatus (mean ± se) ranged between 17.1 ± 3.7 and 770.7 ± 443.1 ind/m2. In 2009 the lowest abundance was 34.4 ± 15.2 and the highest 892.5 ± 34.4 ind/m2. During 2009 biomass and production was more than twice higher in comparison with 2008 (Table 1).
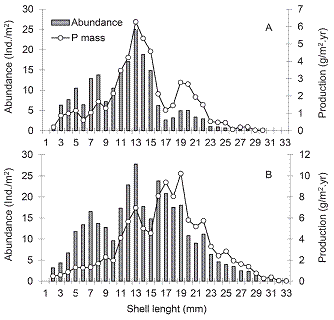
Individual production showed highest values in the 24 mm shell size (3.74 g/m2.yr) and 25 mm (0.71 g/m2.yr), considering TM and WM respectively (Fig. 3). During 2008 the 13 mm size class contributed with the highest density and production (Fig. 4A), whereas in 2009 individuals of 13 mm contributed with the higher abundance too, but the higher production were reported for the 19 mm size class (Fig. 4B). Considering AFDM, the renewal rate was 5.11 in 2008 and 3.47 in 2009.
Table 1: Biomass (B) and production estimation (P) of D. striatus from Las Balsas beach in 2008 and 2009

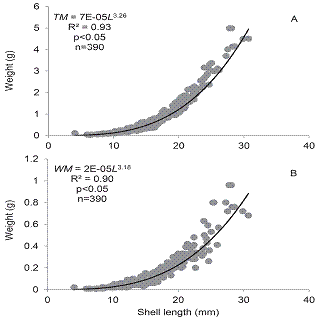
Fig. 2: Length-weight relationship and model fit considering total mass (A) and wet mass (B) of D. striatus from Las Balsas beach.
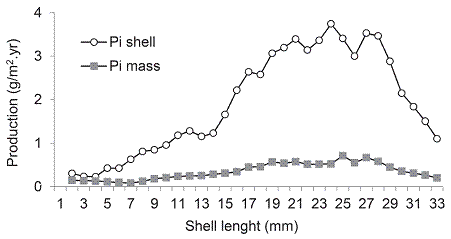
Fig. 3: Individual somatic production for different length classes of D. striatus from Las Balsas beach. Pi shell: individual somatic production for total mass with shell, Pi mass: individual somatic production for wet mass.
Discussion
The maximum shell length (L= 33.3 mm) of the Cuban D. striatus population, recorded during this study, is the largest currently known length of this species. Wade (1967) commented that D. striatus seldom reaches 35 mm but the populations studied by this author did not show evidence of individuals larger than L= 30 mm. In this population there is low abundance of specimens reaching L > 30 mm, one of the reasons being probably that they are fished throughout the year, and then a higher probability of fishing mortality in the selection by size is combined with natural mortality.
The estimated K value for this population is higher than the one reported by Farache (1980) in a Venezuelan beach (0.29/yr) but lower than that estimated in a Brazilian beach (1.16/yr) by Rocha-Barreira et al. (2002). The growth rate calculated for this population is relatively high. Herrmann et al. (2009) made a compilation of several studies on different species of Donax to compare growth among them in different climatic areas. According to these data, D. striatus from Las Balsas beach had higher growth performance than populations of other Donax species inhabiting the same climate region. These values match with other species of the same genus located in temperate zones, but that have reach longer shell sizes; this could be explained by the estimation method used in this study (Ocana, Apin, & Cala, 2013). Growth estimates based on length-frequency data may be biased for tropical species which present continuous recruitment along the year (Sparre & Venema, 1997).
Most species of Donax have longevity of one or two years, whereas mortality is often high, particularly in tropical species (Ansell, 1983). The life span of 2.4 yrs calculated in this study is higher than precedent values found in other populations of the same species and even higher than D. denticulatus that has the same distribution range. Other populations of Donax in the Caribbean (Wade, 1968; Velez, Venables, & Fitzpatrik, 1985; McLachlan et al., 1996; Rocha-Barreira et al., 2002; Marcano, Prieto, Larez, & Salazar, 2003; Ocana et al., 2013) do not last two years. Mortality rate obtained for D. striatus was higher than other tropical species such as D. incarnatus (Thippeswamy & Mohan Joseph, 1991) and D. dentifer (Riascos & Urban, 2002), was similar compared with D. obesulus (Velarde & Agui-lar, 2008), and lower than a population of D. denticulatus from South-eastern Cuba (Ocana et al., 2013). It is necessary to remark that this study estimated the total mortality rate for the population of Las Balsas beach. Because there are no official catch statistics, the fishing mortality could not be estimated. High mortality could be related with the direct impact of removing organisms, and the alteration of habitat features due to human trampling that could affect young individuals as suggested by Rocha-Barreira et al. (2002).
Abundance varies between years, thus mean annual biomass and secondary production were more than double in 2009 when compared with 2008. This result suggests that there is not a clear seasonal pattern of abundance in this population. Herrmann et al. (2009) found that abundance of D. hanleya-nus varied between years at two of the three studied beaches, therefore it could be possible that the inter-annual variation of abundance is population-specific instead of species-specific. In this population there is a coincidence that size classes with higher abundance had higher contribution to the total production in the studied period. In 2009 the abundance of individuals with L > 15 mm increased in relation with 2008, this result could be explained by clima-tological events occurred during 2008 (Ocana et al., 2010) that might cause higher mortality of adults individuals or their migration to the subtidal zone, so they could not be collected in my sampling in 2008.
Values of biomass and production are higher than those obtained for other tropical species such as D. incarnatus, D. spiculum (Ansell, McLusky, Stirling, & Trevallion, 1978) and D. dentifer (Velarde & Aguilar, 2008). Secondary production of D. striatus is higher than the overall median estimated by Petracco, Cardoso, and Turra (2013) to bivalves of sandy beaches. High renewal rate matches with values of some tropical species according to Herrmann et al. (2009), nevertheless is lower than P/B rate reported for D. incarnatus, D. spiculum (Ansell et al., 1978) and for D. denticulatus (Velez et al., 1985; Ocana et al., 2013). The rapid growth and high turnover rate of D. striatus suggest a rapid recovery of the population. These results support the idea that this beach clam is an important resource at Las Balsas beach. Thus its exploitation must continue with caution, and only at the level of a recreational fishery.

