










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 n.2 San José Apr./Jun. 2014
Imposex en Voluta musica (Caenogastropoda: Volutidae) en el Noreste de la Península de Araya, Venezuela
Imposex in Voluta musica (Caenogastropoda: Volutidae) from Northeastern Península de Araya, Venezuela
Imposex in Voluta musica (Caenogastropoda: Volutidae) from Northeastern Península de Araya, Venezuela
*Dirección para correspondencia:
Abstract
Voluta musica is a dioecious marine gastropod endemic of the South Caribbean. Tributyltin (TBT) and copper (Cu) are potential inducers of imposex, an endocrine disorder by which females develop a penis and/or vas deferens. The goal of this work was to determine the imposex incidence in V. musica populations from Northeastern Península de Araya. For this, we selected three sites (Isla Caribe, Isla Lobos and Bajo Cuspe) and made monthly samplings of 15 snails in each site, during one year, and determined: (1) sizes; (2) sex and imposex incidence and (3) the Relative Penis Length Index (RPLI). We also performed histological analysis of the gonads, and measured TBT and Cu concentrations in sediments from the studied localities. Our results showed that the total number of sampled females affected by imposex was 24.5% at Isla Caribe, 12% at Isla Lobos, and none at Bajo Cuspe. In sediments, Cu was detected mostly in Isla Lobos. The female gonads with imposex did not show any development of male cells in any of the sampled sites. The higher percentage of females with imposex matched with the higher boat traffic locality, and higher TBT level (Isla Caribe). No esterilization was evident in this work, nevertheless, the presence of TBT and Cu in the sediments and females with imposex were considered as a potential threat to V. musica populations in this region. In Venezuela there is no control over this particular issue, possibly because of the lack of information and research in this topic, but certainly, this information will be useful in biodiversity conservation policies.
Key word: Volutidae, histology, tributyltin, copper, antifouling, South Caribbean.
Resumen
Voluta musica es un gasterópodo dioico endémico del Caribe sur. El TBT y el Cu, son potenciales causantes del imposex, fenómeno donde las hembras desarrollan un pene y/o vaso deferente. El objetivo fue determinar la incidencia de imposex en V. musica en el noreste de la Península de Araya. Se seleccionaron tres localidades y se captura-ron mensualmente 15 individuos durante un año para determinar: (1) talla de los individuos; (2) sexo y presencia de imposex; (3) índice Largo Relativo del Pene (RPLI). Se realizó histología de la gónada de los individuos. Se determinó TBT y Cu en el sedimento de cada localidad. En Isla Caribe, el 24.5% de las hembras presentó imposex, y se halló 3.9ngSn/g de TBT; en Isla Lobos, el 12% de las hembras desarrollaron imposex; en Bajo del Cuspe no se observó imposex. Se halló Cu en mayor concentración en Isla Lobos. Las gónadas femeninas con imposex no demos-traron masculinización. El mayor porcentaje de imposex coincide con la localidad de mayor tráfico de embarcaciones y con mayor nivel de TBT (Isla Caribe). No se evidenció esterilización, sin embargo la presencia de TBT, Cu e imposex son potenciales amenazas para las poblaciones de V. musica en la región. Hasta ahora, en Venezuela no se está tomando ninguna medida de control sobre este tema en particular, posiblemente por la escasez de información y orientación de las investigaciones hacia este tema, pero que sin duda se debería tomar en cuenta en las políticas para la conservación de la biodiversidad.
Palabras clave: Volutidae, histología, tributilestaño, cobre, pinturas anti-incrustantes, Caribe Sur.
Voluta musica (Linné, 1758) es un gaster-ópodo marino de la familia Volutidae endémico del Caribe sur. Es una especie gonocórica, de desarrollo directo y maduración tardía (Penchaszadeh & Miloslavich, 2001; Peralta, Miloslavich & Bigatti, 2012). V. musica es considerada una especie en riesgo según el Libro Rojo de la Fauna Venezolana; la última edición del Libro Rojo (2008) establece que “El estado actual de sus poblaciones no es conocido con precisión, sin embargo, se ha señalado como una especie bajo presión considerable ya que se encuentra entre las especies de moluscos venezolanos más buscadas por coleccionistas” (Rodriguez & Rojas-Suarez, 2008). Dada su buena cotización en el mercado internacional de conchas, durante algunos años se sostuvo la explotación inescrupulosa de sus poblaciones silvestres (Rodriguez & Rojas-Suarez, 2008). Actualmente las conchas de V. musica se venden a través de mercados disponibles por internet y su precio varía entre 7 y 1 130US$ por ejemplar, dependiendo del tamaño de la concha, y del patrón de coloración, siendo la mayoría de las conchas provenientes de Venezuela (Conchology, 2013; eBay 2013; Shellmesh, 2013).
Isla Caribe, Isla Lobos y Bajo Cuspe son tres sitios ubicados al noreste de la Península de Araya en Venezuela donde habitan poblaciones de V. musica. Isla Caribe es un pequeño islote donde existe un asentamiento permanente de pescadores artesanales desde hace al menos 30 años, con un tráfico marino relativamente alto llevado a cabo por seis botes de 7m de eslora cada uno, fondeados en la costa a una profundidad máxima de 1.5m y al cual se suman embarcaciones que amarran en profundidades someras en la misma zona por temporadas. El segundo sitio, Isla Lobos, es otro pequeño islote localizado a 2km al oeste del primero. El ecosistema marino es semejante al de Isla Caribe, pero el tráfico marino es relativamente bajo, ya que al sitio llega un bote de 7m de eslora por semana y actualmente no hay asentamientos pesqueros. El tercer sitio, Bajo Cuspe, se encuentra a 3km al norte de Isla Caribe e Isla Lobos, no presenta botes permanentes pero existe cierto tráfico de embarcaciones con fines turísticos o de simple traslado de pasajeros entre las islas y el continente. Todos los botes de estos sitios, utilizados tanto para la pesca como para el traslado turístico, están pintados con pinturas anti-incrustantes a base de Cu y/o TBT, constituyendo un potencial foco de contaminación para la fauna macrobentónica ubicada en la zona de anclaje y desembarque.
El tributilestaño (TBT) es un compuesto que comenzó a utilizarse en la década de los 60 como biocida en las pinturas anti-incrustantes para embarcaciones, redes para acuicultura y en sistemas de enfriamiento industrial y que fue mundialmente prohibido en el año 2008 (Organización de las Naciones Unidas para la agricultura y alimentación, 2006). Muchos gasterópodos marinos son sensibles al tributil-estaño, el cual se ha identificado como el causante del fenómeno de imposex, un desorden de tipo endocrino que consiste en el desarrollo de caracteres sexuales masculinos en las hembras. De esta manera, las hembras desarrollan un pene, y en algunos casos también un vaso deferente, como una respuesta biológica espe-cífica al contacto con este compuesto (Smith, 1971, 1981). Algunos autores señalan que el cobre también podría inducir la formación de órganos masculinos en hembras (Nias, McKil-lup & Edyvane 1993; Gibbs & Bryan, 1994), sin embargo, esto no ha sido comprobado. El imposex ha sido reportado en los Neogastrop-oda, presentándose en más de 260 especies en el mundo y observándose respuestas a partir de concentraciones de TBT en el agua inferiores a 1ng/L (Smith, 1981; Gibbs, Bryan, Pascoe, & Burt, 1990; Alzieu, 1991; Stewart, de Mora, Jones, & Miller, 1992; Gooding, Gallardo, & Leblanc, 1999; Castro, Cascon, & Fernandez, 2000; Penchaszadeh, Averbuj, & Cledón, 2001; Horiguchi et al., 2003; Bigatti & Penchasza-deh, 2005; Nishikawa, 2006; Oehlmann et al., 2007; Swennen, Sampantarak, & Ruttanadakul, 2009; Bigatti et al., 2009; Castro, Pernia, & Fillmann, 2012a; Castro & Rocha-barreira, 2012b; entre otros). En Venezuela, existe un solo reporte de imposex, las hembras de las especies de gasterópodos de la pesca artesanal Chicoreus brevifrons, Chicoreus margaritensis y por otro lado el gasterópodo Leucozonia nasa presentaron pene y vaso deferente en una zona con alto tráfico marítimo, mientras que en zonas con bajo tráfico marítimo, el porcentaje de imposex fue nulo (Miloslavich, Penchasza-deh, & Bigatti, 2007).
Dadas las actividades antrópicas que se llevan a cabo en Isla Caribe y que implican la presencia de botes permanentes en la misma isla desde hace poco más de tres décadas, este trabajo tiene como objetivo evaluar la incidencia de imposex sobre las poblaciones de V. musica. Dada la vulnerabilidad de esta especie, esta información es esencial para el establecimiento de pautas de manejo y conservación de sus poblaciones en una de las áreas consideradas como prioritarias para la conservación de la biodiversidad marina de Venezuela (Lazo, Klein, & Villalba, 2008).
Materiales y Métodos
Se recolectaron mensualmente entre 10 y 15 individuos de la especie Voluta musica en las localidades de Isla Caribe (10° 41’ 21.63’’ N - 63° 51’ 08.51’’ W), Isla Lobos (10° 41’ 26.37’’ N - 63° 52’ 28.98’’ W), y Bajo Cuspe (10º43’53’’ N - 63º51’10’’ W), entre febrero 2007 y agosto 2008. El bajo número de muestras se debe a la baja densidad de las poblaciones, característico de los integrantes de la familia Volutidae (Peralta et al., 2012; Big-tti & Penchaszadeh, 2005; Cledón, Theobald, Gerwinski, & Penchaszadeh, 2006). Dada la condición de vulnerabilidad de esta especie, los organismos recolectados para este trabajo fue-ron también utilizados para realizar estudios de ciclos reproductivos, dieta y fecundidad (Peral-ta, 2012). Los organismos fueron recolectados entre 1 y 2m de profundidad en las localidades de Isla Caribe e Isla Lobos y a unos 10m de profundidad en la localidad de Bajo Cuspe. Los individuos fueron llevados vivos al laboratorio del Centro de Investigaciones Ecológicas de Guayacán (CIEG-UDO) donde se determinó la talla de cada individuo mediante el uso de un calibre vernier de precisión 0.01mm. Posteriormente se realizó la disección de todos los individuos y se identificó el sexo de cada uno por la presencia de la glándula del albumen y de la cápsula en las hembras y la ausencia de éstas en los machos. La presencia de imposex se determinó a través de la observación de un pene y/o vaso deferente en las hembras y se calculó el % de incidencia en relación al núme-ro de hembras normales. Se calculó el índice del Largo Relativo del Pene (RPLI, por sus siglas en inglés) que se describe como: (promedio del largo del pene en hembras/promedio del largo del pene en machos) x 100, según la ecuación propuesta por Gibbs & Bryan (1994). Se realizó un análisis de correlación de Pearson entre la talla del pene y la longitud del cuerpo tanto para los machos como para las hembras para evidenciar algún efecto relativo a la talla corporal de los ejemplares.
Para comparar la estructura gonádica de los machos y de las hembras con y sin imposex, se disecó una porción cúbica de 2mm de lado de tejido gonadal de 20 individuos de cada categoría (macho, hembra sin imposex, hembra con imposex), para cada uno de los sitios en donde se halló imposex (Isla Caribe, Isla Lobos) y se fijaron en solución de Bouin. Luego de la fijación se procedió a la deshidratación con alcohol creciente; dos baños de alcohol 70º (media hora cada uno), posteriormente etanol 80º durante una hora, etanol 96º durante una hora, etanol 100º durante una hora y finalmente dos baños de xiloxl (media hora cada uno). Una vez deshidratados, los tejidos fueron incluidos en parafina durante 12 horas. Posteriormente, los trozos de tejido fueron colocados en moldes para formar los bloques que luego fueron montados en las bases para el micrótomo donde se procedió a realizar cortes de 5µm de espesor. El material seccionado fue teñido con hematoxilina-eosina para su análisis histológico al microscopio óptico.
Se estimó la concentración de tributilesta-ño y cobre en el sedimento de cada localidad. Para la evaluación de la presencia de TBT se recolectaron de 50 a 100g de sedimento, tomándose las muestras por triplicado y combi-nándose en una muestra compuesta, la cual fue almacenada en frascos de vidrio, protegidas de la luz con una envoltura de papel de aluminio y transportadas en hielo y conservadas a -20ºC hasta su procesamiento (Bigatti et al., 2009). Las muestras fueron analizadas en el Área de Oceanografía Química del Instituto Argentino de Oceanografía (IADO-CONICET, Argen-tina), mediante GC-MS, siguiendo la técnica descrita en Bigatti et al. (2009). El límite de detección de los organoestaños en el presente estudio fue de 0.2ng Sn/g.
Para la determinación de la concentración de cobre en el sedimento de las tres localida-des, se recolectaron 100g de sedimento por cada localidad y se almacenaron en frascos de vidrio. En el laboratorio, el sedimento fue molido y tamizado por malla de 1mm para separar cuerpos extraños (piedras mayores). Se realizó una emulsión por triplicado, con agua regia (solución compuesta de ácido nítrico 0.05% p/v y ácido clorhídrico 37%), luego se diluyó con agua destilada, se ajustó el pH a 3 y se extrajeron los metales con APDC (Amonio Pirroli-dona Ditiocarbaminato). La lectura se realizó sobre la fase orgánica con llama aire/acetileno. Las muestras fueron analizadas en el Laboratorio de Química Ambiental del Centro Nacional Patagónico-CONICET (Argentina), siguiendo la técnica descrita en Bigatti et al. (2009).
Resultados
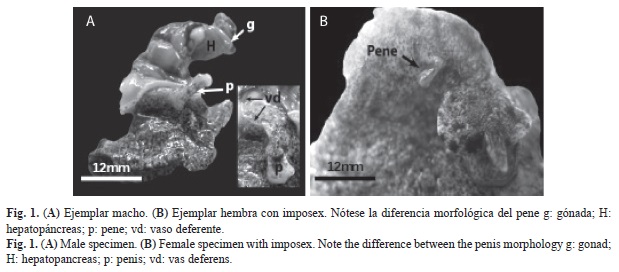
Se registró el fenómeno de imposex por primera vez en V. musica; de 139 hembras capturadas en Isla Caribe, el 24.5% había desarrollado un pene (imposex) cuya longitud promedio total fue de 3.55mm (SD±1.48) (Fig. 1B). Sólo una de las hembras desarrolló el vaso deferente además del pene. En los machos (n=85), solo se observó un individuo con dos penes: el primero de 15mm de longitud y el segundo pene, ubicado por debajo del principal, de 4mm de longitud.
En Isla Lobos, el 12% de las hembras capturadas (n=25) habían desarrollado un pri-mordio peneano (protuberancia) de hasta un máximo de 1mm de longitud (Cuadro 1) y sólo una había desarrollado adicionalmente el vaso deferente. En Bajo del Cuspe, no se observaron alteraciones sexuales en ninguno de los especímenes estudiados (n hembras=81; n machos=38).
El rango de talla de las hembras con alteraciones sexuales halladas en Isla Caribe fue de 60 y 84mm. Adicionalmente, en esta población se observó una correlación entre la talla del pene en las hembras con alteraciones sexuales, y la longitud del cuerpo a partir de los 61mm de longitud corporal (Pearson, r2=0.4639), mientras que en los machos no se observó tal correlación (Pearson, r2=0.0381).
La concentración de TBT encontrada fue de 3.9ngSn/g (SD±3.4) en el sedimento de Isla Caribe, sitio con mayor tránsito de botes pesqueros. No se detectó la presencia de TBT en el resto de los sitios. En los tres sitios muestreados se detectó la presencia de cobre, con una mayor concentración en Isla Lobos (Cuadro 1).
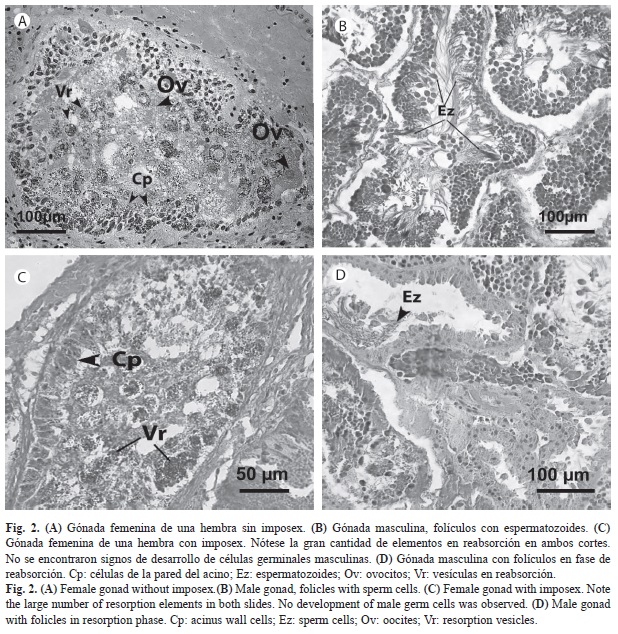
Histología gonadal: Los cortes histológicos de la gónada femenina no demostraron diferencias entre las estructuras gonadales de hembras con y sin imposex. No hay indicios de desarrollo de estructura gonadal masculina en hembras con imposex, ya que la población de hembras con imposex estudiada, no presentó células germinales masculinas asociadas a los acinos. Por otro lado, se observó una intensa reabsorción de oocitos en la mayoría de los cortes de hembras tanto con imposex como sin imposex, así como también un bajo número de ovocitos en cada acino (Fig. 2).
Discusión
Los resultados del presente trabajo se pueden comparar con lo reportado para otros volútidos, como en el caso de Adelomelon brasiliana en Mar del Plata (Argentina), donde se encontró que entre 38.9 y 50% de hembras estaban afectadas con imposex en una zona portuaria donde la concentración de TBT era de 160-5 000ngSn/g (Goldberg, Averbuj, Cledón, Luzzato, & Sbarbati-Nudelman, 2004). En otro trabajo realizado con A. Brasiliana, también en Argentina, se obtuvo un 43% de hembras con imposex con RPLI de 39 y valores de TBT de 12.9ngSn/g, en una zona con alta actividad náutica (Bigatti et al., 2009). En el caso del volútido Odontocymbiola magellanica, existen dos trabajos donde se registró que para zonas con baja actividad náutica el imposex fue nulo y en zonas con alta actividad náuticas se hallaron sitios con hasta 100% de imposex, con RPLI de 26.31 y valores de TBT de 1.7ngSn/g (Bigatti & Penchaszadeh, 2005; Bigatti et al., 2009), los cuales son valores semejantes a lo repor-tado en el presente estudio, pero el porcentaje de hembras afectadas fue muy diferente. En el caso de Adelomelon ancilla (Volutidae), en zonas de alta actividad náutica se halló 100% de imposex con un RPLI de 36.64 y TBT de 1.7ngSn/g, mientras que en zonas con baja actividad náutica se halló un 25% de imposex con un RPLI=0.23 (Bigatti et al., 2009). Si bien nuestros resultados no indican proporciones ni intensidades tan alarmantes de imposex en las poblaciones de V. musica (máximo de 25% de hembras afectadas con RPLI de 28.63), sí se encontró que el mayor porcentaje de incidencia de imposex coincide con la localidad de mayor tráfico de embarcaciones y de mayores niveles de TBT en sedimento (3.9ngSn/g en Isla Cari-be), corroborando los resultados de trabajos previos (Nias et al., 1993; Bigatti et al., 2009; Bigatti & Penchaszadeh, 2005; Castro et al., 2012a; Smith, 1981; Strand & Asmund, 2003; entre otros). Por otra parte, es difícil establecer una estricta relación entre el tráfico de embarca-ciones, la concentración de TBT en sedimento, el % de imposex y su intensidad, debido a que la biodisponibilidad del TBT varía de acuerdo al tamaño del grano de sedimento, el contenido de materia orgánica, corrientes, fuentes, sensi-bilidad especie-específica, entre otros.
Existen algunos autores que cuestionan que el imposex ocurra como resultado de con-taminación sólo por TBT y sugieren que es posible que exista una suma de factores que trabajen en sinergia y que generen desordenes fisiológicos en los organismos que bioacumulan determinados compuestos (Garaventa, Faimali, & Terlizzi, 2006). Por ejemplo, en cuanto a los metales trazas como el cobre, se ha reportado que éstos pueden presentar concentraciones relativamente elevadas en los sedimentos superficiales de las zonas costeras alteradas por el hombre, estando su concentración relacionada con el tamaño de las partículas y la cantidad de materia orgánica en el sedimento, alterando así el equilibrio ecológico y biogeoquímico del ecosistema (Sadiq, 1992). En tal sentido, la determinación de metales en los sedimentos de Isla Caribe, Isla Lobos y Bajo Cuspe es un buen indicador del origen de los contaminantes en el medio y de los potenciales impactos que éstos pueden producir en las poblaciones de Voluta musica. Los botes que llevan a cabo las actividades de pesca en la zona de muestreo, son de madera pintada con pinturas a base de cobre desde hace más de 30 años, constituyendo así una posible fuente de este metal. Así, el contenido de cobre presente en los sedimentos superficiales de las tres localidades estudiadas está probablemente determinado por una combinación de factores incluyendo la magnitud de la fuente. A su vez, en la región donde se realizaron los muestreos transitan embarcaciones con fines turísticos que, según datos informales, utilizan pinturas a base de TBT. Con estos antecedentes se ha evidenciado la presencia de TBT y cobre en los sedimentos de los sitios muestreados, sin embargo, es recomendable investigar más sobre las posibles fuentes de estos compuestos. En el caso de Isla Lobos, las altas concentraciones de cobre podrían deberse a una mayor cantidad de materia orgánica contenida en el sedimento fino en comparación con los otros dos sitios. A pesar de que en el presente estudio no se realizaron análisis granulométricos de los sedimentos, Salazar (1997) reporta que tanto en Isla Caribe como en Isla Lobos la disposición de los sedimentos no es homogénea, encontrándose zonas con grava, arena gruesa y fango, dependiendo de la corriente de fondo y la acción de los vientos Alisios (Salazar, 1997).
Según los niveles de calidad de sedimento establecidos por diferentes organismos interna-cionales (EPA, 2011, UK Marine Special Areas of Conservation, 2011) Isla Lobos presentó niveles de cobre elevados (21.9ppm). La EPA (Environmental Protection Agency) establece un límite máximo permisible de cobre en sedimento de 29ppm (EPA, 2011), mientras que la UK Marine SACs Project presenta un reporte sobre calidad del sedimento donde se establece un límite máximo permisible de cobre en sedi-mento de 18.7ppm (http://www.ukmarinesac.org.uk; EPA, 2011). Vale la pena mencionar que los sitios muestreados en el presente trabajo posiblemente reciban impactos de la actividad turística a través de embarcaciones que navegan entre la Península de Araya y la Isla de Margarita, lo cual no pudo ser evaluado en el presente estudio; sin embargo, se recomienda su consideración en futuras investigaciones. Asimismo, se recomienda realizar un monitoreo periódico riguroso de la concentración de metales pesados y TBT, así como otros com-puestos orgánicos persistentes en sedimentos y en animales, y su relación con disrupciones endocrinas presentes en zonas impactadas (zonas portuarias y marinas) para marcar pautas sobre valores perjudiciales o tóxicos.
Desde el punto de vista fisiológico algunos trabajos han estudiado las consecuencias que podría tener sobre un individuo el hecho de estar afectado con imposex. Por ejemplo, en la especie Nucella lapillus (neogasterópodo), se observó, bajo condiciones de laboratorio, que algunas hembras desarrollaron esterilidad a concentraciones de TBT en el agua de 1-2ngSn/L y que a concentraciones de 3-5ngSn/L todas las hembras desarrollaron esterilidad. Cuando los niveles de TBT se mantuvieron a 20ngSn/L el proceso de masculinización avanzó al punto en que la oogénesis fue suprimida y se indujo la espermatogénesis (Gibbs & Bryan, 1994). En el presente estudio con el valor de TBT encontrado en los sedimentos (3.9ngSn/g) no se evidenciaron signos de esterilidad ni en la morfología externa ni en los cortes histológicos de gónadas de hembras con pene. Lo registrado aquí para V. musica difiere además de lo reportado en la especie Acanthina monodon (Huaquín, Osorio, Verdugo, & Collado, 2004) donde en las hembras con imposex, la estructura gonadal sufrió alteraciones con indicios de desarrollo de gametos masculinos.
Es interesante mencionar que en el pre-sente trabajo se observó sólo un macho con dos penes, lo cual difiere de lo reportado sobre imposex en los volutidos Adelomelon ancilla y Odontocymbiola magellanica, en los que se han observado hembras con multiples penes (Bigat-ti et al., 2009). Esta diferencia puede deberse a una mayor contaminación en la zona o a la diferente sensibilidad de las especies estudiadas. El nivel de la respuesta biológica inducida por el TBT en los neogasterópodos presenta varias hipótesis según las especies observadas por los distintos autores, pero en general, el TBT parece ser un disruptor endocrino alterando las concentraciones normales de las hormonas sexuales (Spooner, Gibbs, Bryan, & Goad, 1991; Oberdorster & McClellan-Green, 2000). Se ha reportado que los organoestaños pueden afectar el desarrollo gonadal de las hembras a través de la supresión de la hormona para la ovulación cuya ausencia desencadena la degeneración de los ovocitos maduros (Jong Brink, Boer, & Joosse, 1983). En el presente trabajo se observó una intensa degeneración de ovocitos en todos los cortes histológicos de hembras de tallas adultas, lo que apoyaría la hipótesis planteada por Jong Brink et al. (1983), por lo tanto se sugiere profundizar las observaciones y los análisis en este aspecto y corroborar si la intensa degeneración observada en V. musica se debe a una propiedad intrínseca de la especie o si es una consecuencia del hábitat impactado por el tráfico marítimo y la actividad pesquera.
Dados los antecedentes en cuanto a los efectos nocivos del TBT sobre los organismos marinos, se estableció su restricción en Francia en 1982 en donde se prohibió la aplicación de TBT en pinturas anti-incrustantes en embarcaciones menores a 25m de eslora. Posteriormen-te, la prohibición se extendió hacia el Reino Unido (1987), Estados Unidos (1988), Suecia, Nueva Zelanda (1989), Australia, Japón (1990) y Dinamarca (1991). En 1999 la Organización Marítima Internacional (IMO) recomendó la prohibición de nuevas aplicaciones en embarcaciones a partir del 2003 y la prohibición completa de la presencia de organoestaños (TBT) como biocida en los buques para el 2008 (Fernández, Limaverde, Castro, Almeida, & Wage-ner, 2002). Bajo este escenario, es sorprendente que en Venezuela para el año 2008 aún seguían ingresando al mercado pinturas antiincrustantes con TBT provenientes del extranjero, sin embargo, estas no son utilizadas actualmente por pescadores artesanales (Peralta, 2012). Los resultados provenientes de encuestas realizadas a pescadores artesanales de la Isla de Margarita durante el 2004 indicaron que tres años antes utilizaban pinturas antiincrustantes a base de TBT y que a partir de entonces utilizan pinturas a base de hierro, cobre y oxido de estaño (Miloslavich et al., 2007). Es posible que el TBT hallado en Isla Caribe sea un remanente que todavía está asociado al sedimento desde hace varios años o que actualmente aún exista algún ingreso de TBT al ambiente. Hasta ahora, en Venezuela no se está tomando ninguna medida de control sobre este tema en particular, posiblemente por la escasez de información y orientación de las investigaciones hacia este tema, pero que sin duda se debería tomar en cuenta en las políticas para la conservación de la biodiversidad.
Agradecimientos
Al Fondo Nacional de Ciencia y Tecno-logía (FONACIT) y a la Universidad Simón Bolívar por el financiamiento brindado para los trabajos de campo. A Felicidad, Felipa y Ramón por todo su apoyo en los muestreos en las islas.
Referencias
Alzieu, C. (1991). Environmental problems caused by TBT in France: Assesment, regulations, prospects. Marine Environmental Research, 32(1-4), 7-17. [ Links ]
Bigatti, G., Antelo, C. S., Miloslavich, P., & Penchaszadeh, P. E. (2009). Feeding behavior of Adelomelon ancilla: a predator neogastropod in Patagonia benthic com-munities. The Nautilus, 123, 159-165. [ Links ]
Bigatti, G. & Penchaszadeh, P. E. (2005). Imposex in Odontocymbiola magellanica (Caenogastropoda: Volutidae) in Patagonia. Comunicaciones de la Socie-dad Malacologica del Uruguay, 9, 371-375. [ Links ]
Bigatti, G., Primost, M. A., Cledón, M., Averbuj, A., Theobald, N., Gerwinski, W., Arntz, W., Morriconi, E., & Penchaszadeh, P. E. (2009). Biomonitoring of TBT contamination and imposex incidence along 4700km of Argentinean shoreline (SW Atlantic: From 38S to 54S). Marine Pollution Bulletin, 58, 695-701. [ Links ]
Castro, I., Cascon, H. M., & Fernandez, M. A. (2000). Imposex em Thais haemastoma (Linnaeus, 1767) (Mollusca: Gastropoda) uma indicacao da contami-nacao por organoestanicos na costa do municipio de fortaleza-ceará-Brasil. Arq. Ciencias Marinas, 33, 143-148. [ Links ]
Castro, I., Pernia, F. C., & Fillmann, G. (2012a). Organo-tin contamination in South American coastal areas. Environmental Monitoring and Assessment, 184, 1781-1799. [ Links ]
Castro, I., & Rocha-barreira, C. D. E. A. (2012b). Trans-plant bioassay induces different imposex responses in two species of the genus Stramonita. Marine Biology Research, 8, 331-338. [ Links ]
Cledón, M., Theobald, N., Gerwinski, W., & Penchasza-deh, P. E. (2006). Imposex and organotin compounds in marine gastropods and sediments from the Mar del Plata coast, Argentina. Journal of the Marine Biologi-cal Association of the United Kingdom, 86, 751-755. [ Links ]
Conchology. (2013). Voluta musica. Recuperado de http://www.conchology.com [ Links ]
eBay. (2013). Voluta musica shells. Recuperado de http://www.ebay.com [ Links ]
EPA. (2011). Sediment Quality Guidelines. Recuperado de http://water.epa.gov/polwaste/sediments/cs/guide-lines.cfm [ Links ]
Fernández, M., Limaverde, A. M., Castro, I., Almeida, A. C. M., & Wagener, A. de L. R. (2002). Ocurrence of imposex in Thais haemastoma: possible evidence of environmental contamination derived from organotin compounds in Rio de Janeiro and Fortaleza, Brasil. Cadernos de Saúde Pública, 18, 463-475. [ Links ]
Garaventa, F., Faimali, M., & Terlizzi, M. (2006). Imposex in pre-pollution times. Is TBT to blame? Marine Pollution Bulletin, 52, 696-718. [ Links ]
Gibbs, P. E. & Bryan, G. W. (1994). Biomonitoring of Tri-butyltin (TBT) pollution using the Imposex response of Neogastropod Molluscs. In K. J. M. Kramer (Ed.). Biomonitoring of Coastal Waters and Estuaries (pp. 205-226). Boca Raton: CRC Press. [ Links ]
Gibbs, P. E., Bryan, G. W., Pascoe, P. L., & Burt, G. R. (1990). Reproductive abnormalities in female Ocenebra crinacea (Gastropda) resulting from tributyltin-induced imposex. Journal of the Marine Biological Association of the United Kingdom, 70, 639-656. [ Links ]
Goldberg, R. N., Averbuj, A., Cledón, M., Luzzatto, D., & Sbarbati-Nudelman, N. (2004). Search for trior-ganotins along the Mar del Plata (Argentina) marine coast: finding of tributyltin in egg capsules of a snail Adelomelon brasiliana (Lamarck, 1822) population showing imposex effects. Applied Organometallic Chemistry, 18, 117-123. [ Links ]
Gooding, M., Gallardo, C., & Leblanc, G. (1999). Impo-sex in three marine gastropod species in Chile and potential impact on muriciculture. Marine Pollution Bulletin, 38, 1227-1231. [ Links ]
Horiguchi, T., Li, Z., Uno, S., Shimizu, M., Shiraishi, H., Morita, M., Thompson, J. A. J., & Levings, C. D. (2003). Contamination of organotin compounds and imposex in molluscs from Vancouver, Canada. Marine Environmental Research, 57, 75-88. [ Links ]
Huaquín, L. G., Osorio, C., Verdugo, R., & Collado, G. (2004). Morphological changes in the reproductive system of females Acanthina monodon (Pallas, 1774) (Gastropoda:Muricidae) asffected by imposex from the coast of central Chile. Invertebrate Reproduction and Development, 46, 111-17. [ Links ]
Jong-Brink, M., Boer, H. H., & Joosse, J. (1983). Oogene-sis, Oviposition and Oosorption, In K. G. A. Adiyodi & R.G. Adiyodi (Eds.). Reproductive Biology of Invertebrates (pp. 297-355). New York: John Wiley & Sons, Press. [ Links ]
Lazo, R., Klein, E., & Villalba, C. (2008). Portafolio de sitios prioritarios para la conservación. In E. Klein (Ed.). Prioridades De PDVSA En La Conservación De La Biodiversidad En El Caribe Venezolano (pp. 42-51). Caracas: Petróleos de Venezuela, S. A. - Universidad Simón Bolívar - The Nature Conservancy. [ Links ]
Miloslavich, P., Penchaszadeh, P. E., & Bigatti, G. (2007). Imposex en gasteropodos de Venezuela. Ciencias Marinas, 33, 319-324. [ Links ]
Organización de las Naciones Unidas para la Agricultura y la Alimentación. (2006). Convenio de Rotterdam sobre el procedimiento de consentimiento funda-mentado previo aplicable a ciertos plaguicidas y productos químicos peligrosos objeto de comercio internacional: Proyecto de documento de orientación para la adopción de decisiones sobre los compuestos de tributilestaño. Recuperado de http://www.fao.org/docrep/011/y5877s/y5877s00.htm [ Links ]
Nias, D., McKillup, C. S., & Edyvane, K. S. (1993). Imposex in Lepsiella vinosa from Southern Australia. Marine Pollution Bulletin, 26, 380-384. [ Links ]
Nishikawa, J. (2006). Imposex in marine gastropods may be caused by binding of organotins to retinoid X receptor. Marine Biology, 149, 117-124. [ Links ]
Oberdorster, E. & McClellan-Green, P. (2000). The neu-ropeptide APGWamide induces imposex in the mud snail, Ilyanassa obsoleta. Peptides, 21, 1323-1330. [ Links ]
Oehlmann, J., Di Benedetto, P., Tillmann, M., Duft, M., Oetken, M., & Schulte-Oehlmann, U. (2007). Endo-crine disruption in prosobranch molluscs: evidence and ecological relevance. Ecotoxicology, 16, 29-43. [ Links ]
Penchaszadeh, P. E., Averbuj, A., & Cledón, M. (2001). Imposex in gastropods from Argentina (South-West-ern Atlantic). Marine Pollution Bulletin, 42, 790-791. [ Links ]
Penchaszadeh, P. E., & Miloslavich, P. (2001). Embryonic stages and feeding substances of the South American volutid Voluta musica (Caenogastropoda) during intracapsular development. American Malacological Bulletin, 16, 21-23. [ Links ]
Peralta, A. C. (2012). Ecología, Reproducción y Amenazas Potenciales concernientes a Voluta musica (Caeno-gastropoda-Volutidae) en el noreste de la Península de Araya, Venezuela. (Tesis doctoral). Universidad Simón Bolívar, Caracas, Venezuela. [ Links ]
Peralta, A. C., Miloslavich, P., & Bigatti, G. (2012). Comparación de la abundancia, estructura de tallas y fecundidad de Voluta musica (Caenogastropoda: Volutidae ) en tres sitios de la costa norte de la Península de Araya , Venezuela. Revista de Biología Tropical, 60, 165-172. [ Links ]
Rodriguez, J. P. & Rojas-Suarez, F. (2008). Libro Rojo de la Fauna Venezolana. Caracas: Provita y Shell Venezuela, S. A. [ Links ]
Sadiq, M. (1992). Toxic metal chemistry in marine environments. New York: Marcel Dekker. [ Links ]
Salazar, J. (1997). Características granulométricas y sedi-mentológicas de los Islotes Caribe y Lobos. In P. R. Villarroel (Ed.). Islotes Caribe y Lobos. Nueva Esparta Venezuela. (p. 253) Nueva Esparta: Universidad de Oriente. [ Links ]
Shellmesh. (2013). Volutidae shells. Recuperado de http://shellmesh.com/products-page/volutidae [ Links ]
Smith, B. (1971). Sexuality in the American mud snail, Nassarius obsoletus (Say). Proceedings of the Mala-cological Society of London, 39, 377-378. [ Links ]
Smith, B. (1981). Male characteristics on female mud snails caused by antifouling bottom paints. Journal of Applied Toxicology, 1, 22-25. [ Links ]
Spooner, N., Gibbs, P. E., Bryan, G. W., & Goad, L. J. (1991). The effect of tributyltin upon steroid titres in the female dogwhelk, Nucella lapillus, and the development of imposex. Marine Environmental Research, 32, 37-49. [ Links ]
Stewart, C., de Mora, S. J., Jones, M. R. L., & Miller, M. C. (1992). Imposex in New Zealand Neogastropods. Marine Pollution Bulletin, 24, 204-209. [ Links ]
Strand, J., & Asmund, G. (2003). Tributyltin accumulation and effects in marine molluscs from West Greenland. Environmental Pollution, 123, 31-37. [ Links ]
Swennen, C., Sampantarak, U., & Ruttanadakul, N. (2009). TBT-pollution in the Gulf of Thailand: a re-inspection of imposex incidence after 10 years. Marine Pollution Bulletin, 58, 526-32. [ Links ]
UK Marine Special Areas of Conservation. (2011). Summary of saltwater sediment guidelines. Recuperado de http://www.ukmarinesac.org.uk/activities/water-quality/wq4_3_1.htm. [ Links ]
Bigatti, G., Antelo, C. S., Miloslavich, P., & Penchaszadeh, P. E. (2009). Feeding behavior of Adelomelon ancilla: a predator neogastropod in Patagonia benthic com-munities. The Nautilus, 123, 159-165. [ Links ]
Bigatti, G. & Penchaszadeh, P. E. (2005). Imposex in Odontocymbiola magellanica (Caenogastropoda: Volutidae) in Patagonia. Comunicaciones de la Socie-dad Malacologica del Uruguay, 9, 371-375. [ Links ]
Bigatti, G., Primost, M. A., Cledón, M., Averbuj, A., Theobald, N., Gerwinski, W., Arntz, W., Morriconi, E., & Penchaszadeh, P. E. (2009). Biomonitoring of TBT contamination and imposex incidence along 4700km of Argentinean shoreline (SW Atlantic: From 38S to 54S). Marine Pollution Bulletin, 58, 695-701. [ Links ]
Castro, I., Cascon, H. M., & Fernandez, M. A. (2000). Imposex em Thais haemastoma (Linnaeus, 1767) (Mollusca: Gastropoda) uma indicacao da contami-nacao por organoestanicos na costa do municipio de fortaleza-ceará-Brasil. Arq. Ciencias Marinas, 33, 143-148. [ Links ]
Castro, I., Pernia, F. C., & Fillmann, G. (2012a). Organo-tin contamination in South American coastal areas. Environmental Monitoring and Assessment, 184, 1781-1799. [ Links ]
Castro, I., & Rocha-barreira, C. D. E. A. (2012b). Trans-plant bioassay induces different imposex responses in two species of the genus Stramonita. Marine Biology Research, 8, 331-338. [ Links ]
Cledón, M., Theobald, N., Gerwinski, W., & Penchasza-deh, P. E. (2006). Imposex and organotin compounds in marine gastropods and sediments from the Mar del Plata coast, Argentina. Journal of the Marine Biologi-cal Association of the United Kingdom, 86, 751-755. [ Links ]
Conchology. (2013). Voluta musica. Recuperado de http://www.conchology.com [ Links ]
eBay. (2013). Voluta musica shells. Recuperado de http://www.ebay.com [ Links ]
EPA. (2011). Sediment Quality Guidelines. Recuperado de http://water.epa.gov/polwaste/sediments/cs/guide-lines.cfm [ Links ]
Fernández, M., Limaverde, A. M., Castro, I., Almeida, A. C. M., & Wagener, A. de L. R. (2002). Ocurrence of imposex in Thais haemastoma: possible evidence of environmental contamination derived from organotin compounds in Rio de Janeiro and Fortaleza, Brasil. Cadernos de Saúde Pública, 18, 463-475. [ Links ]
Garaventa, F., Faimali, M., & Terlizzi, M. (2006). Imposex in pre-pollution times. Is TBT to blame? Marine Pollution Bulletin, 52, 696-718. [ Links ]
Gibbs, P. E. & Bryan, G. W. (1994). Biomonitoring of Tri-butyltin (TBT) pollution using the Imposex response of Neogastropod Molluscs. In K. J. M. Kramer (Ed.). Biomonitoring of Coastal Waters and Estuaries (pp. 205-226). Boca Raton: CRC Press. [ Links ]
Gibbs, P. E., Bryan, G. W., Pascoe, P. L., & Burt, G. R. (1990). Reproductive abnormalities in female Ocenebra crinacea (Gastropda) resulting from tributyltin-induced imposex. Journal of the Marine Biological Association of the United Kingdom, 70, 639-656. [ Links ]
Goldberg, R. N., Averbuj, A., Cledón, M., Luzzatto, D., & Sbarbati-Nudelman, N. (2004). Search for trior-ganotins along the Mar del Plata (Argentina) marine coast: finding of tributyltin in egg capsules of a snail Adelomelon brasiliana (Lamarck, 1822) population showing imposex effects. Applied Organometallic Chemistry, 18, 117-123. [ Links ]
Gooding, M., Gallardo, C., & Leblanc, G. (1999). Impo-sex in three marine gastropod species in Chile and potential impact on muriciculture. Marine Pollution Bulletin, 38, 1227-1231. [ Links ]
Horiguchi, T., Li, Z., Uno, S., Shimizu, M., Shiraishi, H., Morita, M., Thompson, J. A. J., & Levings, C. D. (2003). Contamination of organotin compounds and imposex in molluscs from Vancouver, Canada. Marine Environmental Research, 57, 75-88. [ Links ]
Huaquín, L. G., Osorio, C., Verdugo, R., & Collado, G. (2004). Morphological changes in the reproductive system of females Acanthina monodon (Pallas, 1774) (Gastropoda:Muricidae) asffected by imposex from the coast of central Chile. Invertebrate Reproduction and Development, 46, 111-17. [ Links ]
Jong-Brink, M., Boer, H. H., & Joosse, J. (1983). Oogene-sis, Oviposition and Oosorption, In K. G. A. Adiyodi & R.G. Adiyodi (Eds.). Reproductive Biology of Invertebrates (pp. 297-355). New York: John Wiley & Sons, Press. [ Links ]
Lazo, R., Klein, E., & Villalba, C. (2008). Portafolio de sitios prioritarios para la conservación. In E. Klein (Ed.). Prioridades De PDVSA En La Conservación De La Biodiversidad En El Caribe Venezolano (pp. 42-51). Caracas: Petróleos de Venezuela, S. A. - Universidad Simón Bolívar - The Nature Conservancy. [ Links ]
Miloslavich, P., Penchaszadeh, P. E., & Bigatti, G. (2007). Imposex en gasteropodos de Venezuela. Ciencias Marinas, 33, 319-324. [ Links ]
Organización de las Naciones Unidas para la Agricultura y la Alimentación. (2006). Convenio de Rotterdam sobre el procedimiento de consentimiento funda-mentado previo aplicable a ciertos plaguicidas y productos químicos peligrosos objeto de comercio internacional: Proyecto de documento de orientación para la adopción de decisiones sobre los compuestos de tributilestaño. Recuperado de http://www.fao.org/docrep/011/y5877s/y5877s00.htm [ Links ]
Nias, D., McKillup, C. S., & Edyvane, K. S. (1993). Imposex in Lepsiella vinosa from Southern Australia. Marine Pollution Bulletin, 26, 380-384. [ Links ]
Nishikawa, J. (2006). Imposex in marine gastropods may be caused by binding of organotins to retinoid X receptor. Marine Biology, 149, 117-124. [ Links ]
Oberdorster, E. & McClellan-Green, P. (2000). The neu-ropeptide APGWamide induces imposex in the mud snail, Ilyanassa obsoleta. Peptides, 21, 1323-1330. [ Links ]
Oehlmann, J., Di Benedetto, P., Tillmann, M., Duft, M., Oetken, M., & Schulte-Oehlmann, U. (2007). Endo-crine disruption in prosobranch molluscs: evidence and ecological relevance. Ecotoxicology, 16, 29-43. [ Links ]
Penchaszadeh, P. E., Averbuj, A., & Cledón, M. (2001). Imposex in gastropods from Argentina (South-West-ern Atlantic). Marine Pollution Bulletin, 42, 790-791. [ Links ]
Penchaszadeh, P. E., & Miloslavich, P. (2001). Embryonic stages and feeding substances of the South American volutid Voluta musica (Caenogastropoda) during intracapsular development. American Malacological Bulletin, 16, 21-23. [ Links ]
Peralta, A. C. (2012). Ecología, Reproducción y Amenazas Potenciales concernientes a Voluta musica (Caeno-gastropoda-Volutidae) en el noreste de la Península de Araya, Venezuela. (Tesis doctoral). Universidad Simón Bolívar, Caracas, Venezuela. [ Links ]
Peralta, A. C., Miloslavich, P., & Bigatti, G. (2012). Comparación de la abundancia, estructura de tallas y fecundidad de Voluta musica (Caenogastropoda: Volutidae ) en tres sitios de la costa norte de la Península de Araya , Venezuela. Revista de Biología Tropical, 60, 165-172. [ Links ]
Rodriguez, J. P. & Rojas-Suarez, F. (2008). Libro Rojo de la Fauna Venezolana. Caracas: Provita y Shell Venezuela, S. A. [ Links ]
Sadiq, M. (1992). Toxic metal chemistry in marine environments. New York: Marcel Dekker. [ Links ]
Salazar, J. (1997). Características granulométricas y sedi-mentológicas de los Islotes Caribe y Lobos. In P. R. Villarroel (Ed.). Islotes Caribe y Lobos. Nueva Esparta Venezuela. (p. 253) Nueva Esparta: Universidad de Oriente. [ Links ]
Shellmesh. (2013). Volutidae shells. Recuperado de http://shellmesh.com/products-page/volutidae [ Links ]
Smith, B. (1971). Sexuality in the American mud snail, Nassarius obsoletus (Say). Proceedings of the Mala-cological Society of London, 39, 377-378. [ Links ]
Smith, B. (1981). Male characteristics on female mud snails caused by antifouling bottom paints. Journal of Applied Toxicology, 1, 22-25. [ Links ]
Spooner, N., Gibbs, P. E., Bryan, G. W., & Goad, L. J. (1991). The effect of tributyltin upon steroid titres in the female dogwhelk, Nucella lapillus, and the development of imposex. Marine Environmental Research, 32, 37-49. [ Links ]
Stewart, C., de Mora, S. J., Jones, M. R. L., & Miller, M. C. (1992). Imposex in New Zealand Neogastropods. Marine Pollution Bulletin, 24, 204-209. [ Links ]
Strand, J., & Asmund, G. (2003). Tributyltin accumulation and effects in marine molluscs from West Greenland. Environmental Pollution, 123, 31-37. [ Links ]
Swennen, C., Sampantarak, U., & Ruttanadakul, N. (2009). TBT-pollution in the Gulf of Thailand: a re-inspection of imposex incidence after 10 years. Marine Pollution Bulletin, 58, 526-32. [ Links ]
UK Marine Special Areas of Conservation. (2011). Summary of saltwater sediment guidelines. Recuperado de http://www.ukmarinesac.org.uk/activities/water-quality/wq4_3_1.htm. [ Links ]
*Correspondencia a:
Ana Carolina Peralta. Universidad Simón Bolívar, Departamento de Estudios Ambientales, Centro de Biodiversidad Marina, Laboratorio de Biología Marina, Apdo. 89000 Caracas 1086-A Venezuela. Correo electrónico aperalta@usb.ve
Patricia Miloslavich. Universidad Simón Bolívar, Departamento de Estudios Ambientales, Centro de Biodiversidad Marina, Laboratorio de Biología Marina, Apdo. 89000 Caracas 1086-A Venezuela. Correo electrónico pmilos@usb.ve
Gregorio Bigatti. LARBIM. Centro Nacional Patagónico (CENPAT) – CONICET, Puerto Madryn, Argentina. Correo electrónico gbigatti@cenpat.edu.ar
1. Universidad Simón Bolívar, Departamento de Estudios Ambientales, Centro de Biodiversidad Marina, Laboratorio de Biología Marina, Apdo. 89000 Caracas 1086-A Venezuela. Correo electrónico aperalta@usb.ve, pmilos@usb.ve
2. LARBIM. Centro Nacional Patagónico (CENPAT) – CONICET, Puerto Madryn, Argentina. Correo electrónico gbigatti@cenpat.edu.ar
Recibido 26-iv-2013. Corregido 04-Xi-2013. Aceptado 02-Xii-2013.


{kind=link}
{kind=link}