










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.59 n.4 San José Dec. 2011
Walter S. Araújo
Pós-Graduação em Ecologia e Evolução, Instituto de Ciências Biológicas, Universidade Federal de Goiás. Campus Samambaia, 74001-970, Goiânia, Goiás, CP 131, Brazil; walterbioaraujo@yahoo.com.br
Dirección para correspondencia
Abstract
Many hypotheses have been proposed to explain the diversity of gall-midge insects (Diptera: Cecidomyiidae), some of them taking into account plant diversity. This study aims to test the importance of size, age and composition of host plant taxa in the diversity of Cecidomyiidae. For this we used inventories data on the diversity of galling and host plants in Brazil. We found that Asterales, Myrtales and Malpighiales, were the most important orders, with 34, 33 and 25, gall morphotypes, respectively. The most representative host families were Asteraceae (34 morphotypes), Myrtaceae (23) and Fabaceae (22). In general, the order size and the plant family were good predictors of the galling diversity, but not the taxon age. The most diverse host genera for gall-midges were Mikania, Eugenia and Styrax, with 15, 13 and nine galler species, respectively. The size of plant genera showed no significant relationship with the richness of Cecidomyiidae, contrary to the prediction of the plant taxon size hypothesis. The plant genera with the greatest diversity of galling insects are not necessarily those with the greatest number of species. These results indicate that some plant taxa have a high intrinsic richness of galling insects, suggesting that the plant species composition may be equally or more important for the diversity of gall-midges than the size or age of the host taxon. Rev. Biol. Trop. 59 (4): 1599- 1607. Epub 2011 December 01.
Key words: Cecidomyiidae, floristic composition, galls, hypotheses, plant richness, superhosts.
Resumen
Palabras clave: agallas, Cecidomyiidae, composición florística, hipótesis, riqueza de plantas, superhospedeiras.
Gall-midges of the family Cecidomyiidae (Diptera) are the most diverse group of galling insects (Gagné 1994, 2004, Carneiro et al. 2009). These insects are found in all zoogeographical regions and comprise about 5 400 species and 598 genera (Gagné 2004). In the Neotropical region about 170 genera are known, and just over 159 species and 75 genera are recorded in Brazil (Maia 2005).
Forecasts indicate that the number of species of Cecidomyiidae might be about 85 000, representing 64% of the total diversity of galling insects in the world (Espírito-Santo & Fernandes 2007). This can be attributed to adaptive radiation within the group to be largely opportunistic (Price 2005), species within an insect genus can be found in many genera and families of host plants, as occurs in Dasineura Rondani, 1840 (Diptera: Cecidomyiidae) (Gagné 1989). Although gall-midge species are highly specific (Carneiro et al. 2009), the genera do not seem to be, thus promoting the high richness within the group.
Many hypotheses have been proposed to explain the diversity of gall-inducing insects, providing explanations at the intra-specific, interspecific and community levels (Fleck & Fonseca 2007). Some of these hypotheses take into account the possibility that plant diversity influences galling diversity both temporarly and spatially, for example, the plant taxon size hypothesis and plant taxon age hypothesis. The first hypothesis proposes that the most diverse host taxa have a greater galling richness, taking into account that each host species is a potential niche for insects (Mendonça 2007). The second proposes that older host taxa should have more species of gallers, since greater evolutionary time leads to an increase in number of speciation events (Fernandes 1992). Thus, one would expect a positive correlation between the number of species of gall-midges and size and age of the host taxon.
In a pioneer study, Veldtman & McGeoch (2003) proposed that plant community composition is a determining factor in the diversity of gall-inducing insects. According to them, some taxa, known as super-hosts, increase the diversity of galling insects regardless of their size or age. This issue becomes more important at lower taxonomic levels (eg., genera and species), whereas in higher taxa, the composition effect is dissipated by the high number of species. Thus, if composition is an important factor, one would expect no correlation, or a negative correlation, between the size of the plant genera, and the richness of gall-midge, thus contradicting the hypothesis of taxon size, which should be important in explaining diversity at higher taxonomic levels such as family and order. The aim of this study is to answer the following questions: 1) Is the richness of gall-midges proportional to plant taxon size? 2) Do older host taxa have a greater richness of Cecidomyiidae? and 3) Is taxa composition an important factor for the diversity of gall-midges?
Material and methods
The database of gall-midges and their host plants was obtained from Carneiro et al. (2009). In this paper, the author conducted a comprehensive review and listed all Brazilian species of gall-midges and their host plants registered in the literature. In addition, I used some data of Cecidomyiidae from a series of inventories of gall diversity in the state of Goiás (Araújo et al. 2007a, b, Santos et al. 2010, Araújo et al., unpublished results), totaling six families and 14 genera of host plants added. Gall-midges were separated by species or, if undetermined, by gall morphotype. Data on the size of plant order, families and genera were obtained from Judd et al. (2002) and Souza & Lorenzi (2008). The age of the taxa was obtained from Wikström et al. (2001). Multiple regression and linear regression analysis was used to correlate gall-midges richness to the size and age of botanical taxa in the software Statistica 7.0 (all data were submitted to normality tests and other assumptions). The analyses were done at the levels of order, family and genus of the host plants. Altogether 233 gall morphotypes were analyzed, belonging to 23 orders, 49 families and 102 genera of plants.
Results
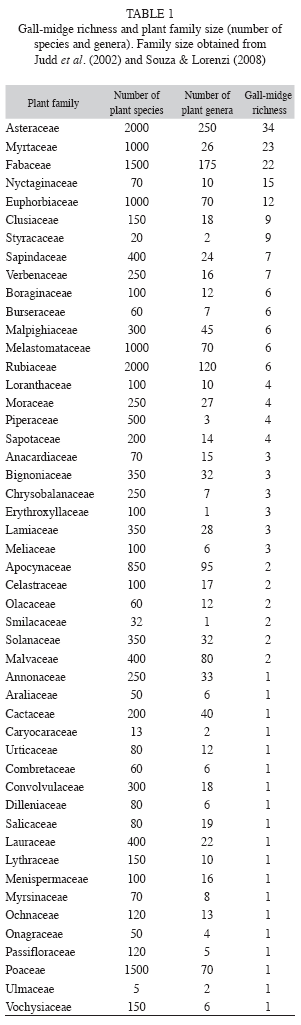
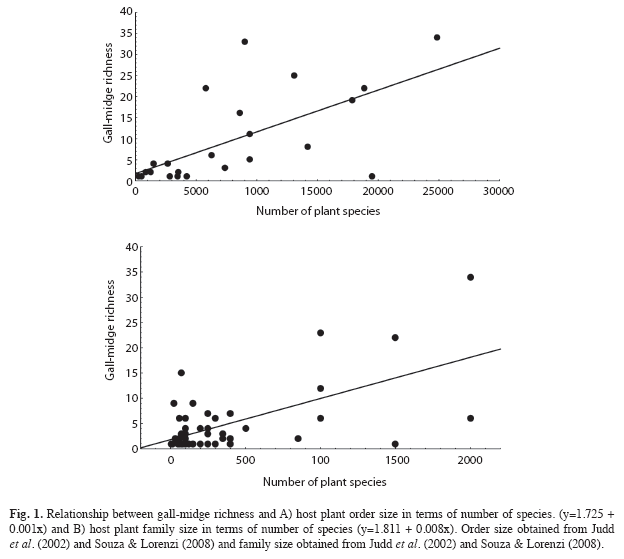
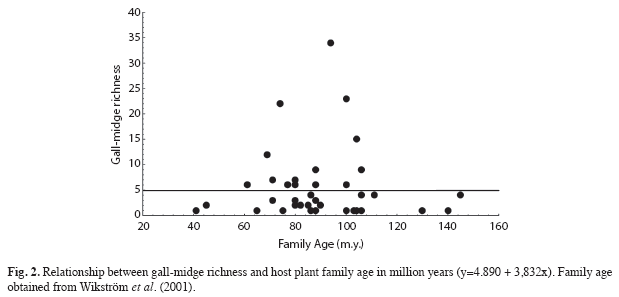
Asterales, Myrtales and Malpighiales were the most important orders of host plant of gall-midges in Brazil, with 34, 33 and 25 galls morphotypes, respectively. The most representative host families were Asteraceae (34 morphotypes), Myrtaceae (23) and Fabaceae (22) (Table 1). Gall-inducing insect richness was positively related with the size of the host plant order, in terms of number of species (r2=0.59, n=23, p<0.01, Fig. 1A), but not with the age of the order (p=0.59). Size of the host plants accounted for 59% of gall-inducing insect richness variation. Gall-midge richness was also positively correlated with the size of the plant family (r2=0.48, n=49, p<0.001, Fig. 1B). However, the age of the plant families were not related to gall-midge richness (p=0.32, Fig. 2).
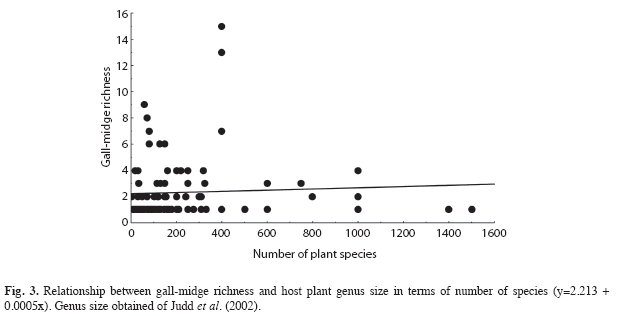
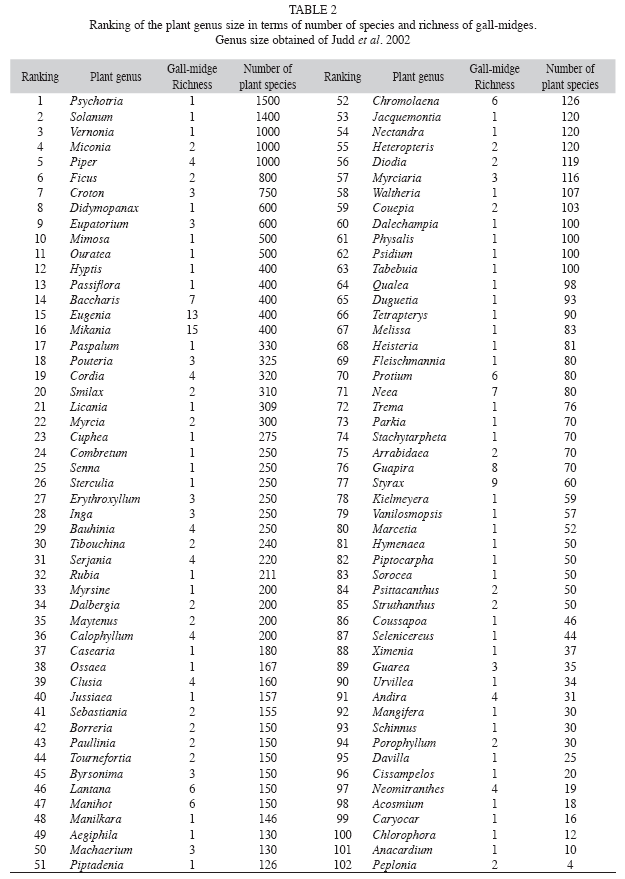
Unlike what was observed for the size of plant order and family, a positive and linear relationship between the size of plant genera and gall-midge richness was not found (r2=0.002, n=102, p=0.60, Fig. 3). The size of plant genera ranged from four to 1 500 species (mean 218.9±271.9). The most diverse host genera for gall-midges were Mikania, Eugenia and Styrax, with 15, 13 and nine galler species, respectively (Table 2). These genera occupy only the 15th, 16th and 77th positions in genus size ranking. The three plant genera with the most species of plants were Psychotria, Solanum and Vernonia, but, these genera had only one species of gall-midge each (Table 2).
Discussion
The diversity patterns of gall-midges observed in this study are supported by previous investigation where the size of plant taxon was positively correlated with the richness of gall-inducing insects (Fernandes 1992, Gonçalves- Alvim & Fernandes 2001, Mendonça 2007). As expected, the most diverse plant taxa in terms of number of species had a greater diversity of gall-midges. At the family level, in addition to the number of species, the number of genera was also a predictor of diversity of Cecidomyiidae.
The plant taxon size hypothesis has been tested mainly for the taxonomic level of family (also known as the plant family size hypothesis) (Fernandes 1992), while at the order level, the hypothesis has rarely been tested (for example, Mendonça 2007). Gonçalves-Alvim & Fernandes (2001) found a positive relationship between family size and gall richness in the Cerrado of Southeast of Brazil, where Fabaceae was the most important family. Fabaceae is the most species rich family in the Brazilian Cerrado, peaking at 777 species (Mendonça et al. 1998), and is also registered as the host family with the greatest number of Neotropical cecidomyiid galls (Gagné 1994). Araújo et al. (unpublished results) also found Fabaceae to be important in areas of the Cerrado in Goiás State in Midwestern Brazil, so that when the family was excluded from the analysis the positive relationship between family size and richness gall-inducing insects disappeared.
In this study, Asteraceae was the host family with most species of Cecidomyiidae. This family is one of the most diverse not only in Brazil but also throughout the neotropics (Judd et al. 2002), being recorded in Southern Brazil as the most important host family for gall inducers (Mendonça 2007). According to Gagné (1994) this family is the second largest host family of gall-midges in the Neotropical region.
However, unlike taxon size, taxon age was not a good predictor of diversity of Cecidomyiidae. Older orders and families did not have a greater diversity of gall-midges. The age hypothesis suggests that geologically older taxa offer more opportunities for the host plant change of galling insects and thus accumulate a greater number of species over time (Fernandes 1992, Mendonça 2007). However, contrary to what the hypothesis predicts, some relatively young taxa, such as Asteraceae and Fabaceae, have a higher galling insect diversity than older host plant taxa (Fernandes 1992). In fact, the adaptive opportunism of Cecidomyiidae can explain this (Price 2005). There seems to be a remarkable capacity for host shifting across families and genera, resulting in a wide range of galled plants and adaptive radiation within some families/genera can then proceed (Price 2005). As previously mentioned, the most diverse taxa have higher rates of speciation and synchrony (Price 2002). This makes measures of plant diversity, local richness of host plants (Cuevas-Reyes et al. 2004) or taxon size (Araújo et al. unpublished results), in terms of number of genera or species, better predictors of gall-midge diversity than taxon age.
All of these patterns observed for the size of the plant orders and families and flora composition, indicate the importance of plant community to galling diversity (Veldtman & McGeoch 2003). However, there is a chance that these observations may be sampling artifacts, as argued by Fleck & Fonseca (2007). According to them, the most diverse taxa are probably sampled more and super-hosts may be the result of an over-sampling. On the other hand, if this were true, major genera have a higher diversity of gall-midges and superhosts taxa would also be the most diverse in species number. Further studies are necessary to address these issues at local and regional scales. Moreover, these questions should be examined in other groups of galling insects. Apparently, the plant taxon size hypothesis as well as the plant composition hypothesis, are good predictors of gall-midges diversity.
Acknowledgments
I am very grateful to V.C. Maia, M.S. Mendonça Jr. and M.A.A. Carneiro for their important suggestions and criticisms of the manuscript; to N. Coelho for English revision and to Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) for the scholarship grant.
References
Araújo, W.S. & B.B. Santos. 2009. Efeitos da sazonalidade e do tamanho da planta hospedeira na abundância de galhas de Cecidomyiidae (Diptera) em Piper arboreum (Piperaceae). Rev. Bras. Entomol. 53: 300-303. [ Links ]
Araújo, W.S., V.L. Gomes-Klein & B.B. Santos. 2007a. Galhas entomógenas associadas à vegetação do Parque Estadual da Serra dos Pireneus, Pirenópolis, Goiás, Brasil. R. Bras. Bioci. 5: 45-47. [ Links ]
Araújo, W.S., B.B. Santos, H.D. Ferreira & T.C. Lousa. 2007b. Ocorrência de galhas entomógenas na vegetação do Campus da UFG, em Goiânia, Goiás. R. Bras. Bioci. 5: 57-59. [ Links ]
Blanche, K.R. & M. Westoby. 1995. Gall-forming insect diversity is linked to soil fertility via host plant taxon. Ecology 76: 2334-2337. [ Links ]
Carneiro, M.A.A., C.S.A. Branco, C.E.D. Braga, E.D. Almada, M.B.M. Costa, V.C. Maia & G.W. Fernandes. 2009. Are gall midge species (Diptera, Cecidomyiidae) host-plant specialists? Rev. Bras. Entomol. 53: 365-378. [ Links ]
Cuevas-Reyes, P., M. Quesada, P. Hanson, P. Dirzo & K. Oyama. 2004. Diversity of gall-inducing insects in a Mexican tropical dry forest: the importance of plant species richness, life-forms, host plant age and plant density. J. Ecol. 92: 707-716. [ Links ]
Espírito-Santo, M.M. & G.W. Fernandes. 2007. How many species of gall-inducing insects are there on Earth and where are they? Ann. Entomol. Soc. Am. 100: 95-99. [ Links ]
Fernandes, G.W. 1992. Plant size family and age effects on insular gall-forming species richness. Global Ecol. Biogeog. 2: 71-74. [ Links ]
Fernandes, G.W., M.A.A. Carneiro, A.C.F. Lara, L.A. Allain, G.I. Andrade, G.R. Julião, T.C. Reis & I.M. Silva. 1996. Galling insects on neotropical species of Baccharis (Asteraceae). Trop. Zool. 9: 315-332. [ Links ]
Fleck, T. & C.R. Fonseca. 2007. Hipóteses sobre a riqueza de insetos galhadores: uma revisão considerando os níveis intra-específico, interespecífico e de comunidade. Neotrop. Biol. Conserv. 2: 36-45. [ Links ]
Gagné, R.J. 1989. The plant-feeding midges of North America. Ithaca, Comstock, USA. [ Links ]
Gagné, R.J. 1994. The gall midges of the region neotropical. Ithaca, Comstock, USA. [ Links ]
Gagné, R.J. 2004. A catalog of the Cecidomyiidae (Diptera) of the world. Mem. Entomol. Soc. Wash. 25: 1-408. [ Links ]
Gonçalves-Alvim, S.J. & G.W. Fernandes 2001. Biodiversity of galling insects: historical, community and habitat effects in four neotropical savannas. Biodivers. Conserv. 10: 79-98. [ Links ]
Judd, W.S., C.S. Campbell, E.A. Kellogg, P.F. Stevens & M.J. Donoghue. 2002. Plant Systematics: A Phyogenetic Approach. Sinauer, Sunderland, Massachussetts, USA. [ Links ]
Maia, V.C. 2005. Catálogo dos Cecidomyiidae (Diptera) do Estado do Rio de Janeiro. Biota Neotrop. 5: 1-15. [ Links ]
Mendonça, R.C., J.M. Felfili, B.M.T. Walter, M.C. Silva- Júnior, A.V. Rezende, T.S. Filgueiras & P.E. Nogueira. 1998. Flora vascular do Cerrado, p. 289-556. In S.M. Sano & S.P. Almeida (eds.). Cerrado: ambiente e flora. Embrapa/CPAC, Brasília, Brasil. [ Links ]
Mendonça, M.S. 2001 Galling insect diversity patterns: the resource synchronization hypothesis. Oikos 95: 171-176. [ Links ]
Mendonça, M.S. 2007. Plant diversity and galling arthropod diversity - searching for taxonomic patterns in an animal-plant interaction in the Neotropics. Bol. Soc. Arg. Bot. 42: 347-357. [ Links ]
Oyama, K., C. Scareli-Santos, M.L. Mondragón-Sánchez, E. Tovar-Sánchez & P. Cuevas-Reyes. 2006. Morphological variations of gall-forming insects on different species of oaks (Quercus) in Mexico, p. 259-269. In M. Kappelle (ed.). Ecology and conservation of neotropical montane oak forests. Springer, Berlin, Germany. [ Links ]
Price, P.W. 2002. Species interactions and the evolution of biodiversity, p. 3-25. In C.M. Herrera & O. Pellmyr (eds.). Plant-animal interactions. An evolutionary approach. Blackwell, Oxford, England. [ Links ]
Price, P.W. 2005. Adaptive radiation of gall-inducing insects. Basic Appl. Ecol. 6: 413-421. [ Links ]
Santos, B.B., H.D. Ferreira & W.S. Araújo. 2010. Ocorrência e caracterização de galhas entomógenas em uma área de floresta estacional semidecídua em Goiânia, Goiás, Brasil. Acta Bot. Bras. 24: 243-249. [ Links ]
Souza, V.C. & H. Lorenzi. 2008. Botânica Sistemática: Guia ilustrado para identificação das famílias de Angiospermas da flora brasileira, baseado em APG II. Nova Odessa, São Paulo, Brasil. [ Links ]
Veldtman, R. & M.A. McGeoch. 2003. Gall-forming insect species richness along a non-scleromorphic vegetation rainfall gradient in South Africa: the importance of plant community composition. Austral Ecol. 28: 1-13. [ Links ]
Weis, A.E. & W.G. Abrahamson. 1986. Evolution of hostplant manipulation by gall makers: ecological and genetic factors in the Solidago-Eurosta system. Am. Nat. 127: 681-695. [ Links ]
Wikström, N., V. Savolainen & M.W. Chase. 2001. Evolution of the angiosperms: calibrating the family tree. Philos. T. Roy. Soc. Lon. B. 268: 2211-2220. [ Links ]
Yukawa, J. 2000. Synchronization of gallers with host plant phenology. Popul. Ecol. 42: 105-113. [ Links ]
Received 25-X-2010. Corrected 27-II-2011. Accepted 30-III-2011.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}