










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.55 n.2 San José Jun. 2007
Reproducción y ultraestructura del huevo y larva de primer estadio de Gyriosomus kingi (Coleoptera: Tenebrionidae) del desierto de Atacama
Jaime Pizarro-Araya1,2,*, Viviane Jerez2 & Jorge Cepeda-Pizarro1
1 Laboratorio de Entomología Ecológica, Departamento de Biología, Facultad de Ciencias, Universidad de La Serena, Casilla 599, La Serena, Chile, Tel.: 56-51-204322, Fax: 56-51-204383, *correo electrónico para correspondencia: japizarro@userena.cl
2 Departamento de Zoología, Facultad de Ciencias Naturales y Oceanográficas, Universidad de Concepción, Casilla 160-C, Concepción, Chile, Tel: 56-41-203884, Fax: 56-41-204805
Recibido 30-I-2004. Corregido 24-V-2006. Aceptado 12-I-2007.
Abstract: Reproduction and ultrastructure of egg and first instar larvae of Gyriosomus kingi (Coleoptera: Tenebrionidae) from the desert of Atacama. Gyriosomus is an important Chilean genus of Tenebrionidae, inhabiting the coastal desert of this country. in this environment, the Gyriosomus-species are both eremic and endemic components of the epigean entomofauna. Despite the ecological importance of these species in the Chilean desert, there is a lack of knowledge on the preimaginal stages of them. in this paper we describe the egg and first instar larval morphology of G. kingi Reed, 1873, along with some bionomic aspects of the female. The eggs and larva were obtained from couples kept under laboratory conditions, after being captured during the spring of year 2002, in coastal sites of National Park Llanos de Challe, located in the southern border of the Atacama Desert (28°01S, 71°03W). The first third of the chorion presents subhexagonal cells lacking aeropyles. The larvae is oligopod and elongated. its cephalic capsule is strongly chitinized with the prothoracic legs longer and thicker than the rest. According laboratory observations, each egg-laying includes seven to ten eggs each time, with a maximum of six egg-laying per female. The eggs are buried into the soil, covered by a mucilaginous layer which creates a clay-sandy film attached to the chorion. Rev. Biol. Trop. 55 (2): 637-644. Epub 2007 June, 29.
Key words: Tenebrionidae, Pimeliinae, insect development, desert soil fauna, Chilean coastal desert.
A pesar de la proliferación de trabajos de tipo ecológicos (Cepeda-Pizarro et al. 1996, 2005 a,b) y sistemáticos (Flores 2000), la mayor parte de la información disponible en la literatura para los Tenebrionidae de Chile está referida a aspectos taxonómicos, con revisiones de tribus y géneros (Kulzer 1954, 1959, Peña 1985, 1995, Flores 1997). Con respecto a la existencia de antecedentes acerca de los estados preimaginales, sólo Artigas y Brañas- Rivas (1973), Cekalovic y Quezada (1973), Cekalovic y Morales (1974) y Pizarro-Araya et al. (2005) han abordado aspectos descriptivos de la morfología larvaria, considerando que la mayoría de la información disponible, está referida a tenebriónidos introducidos con importancia económica (e.g. Tenebrio y Linnaeus, 1758, Gnatocerus Thunberg, 1814, Tenebroides Piller & Mitterpacher, 1783, Tribolium Macleay, 1825) (Artigas 1994). En relación con descripciones de huevos, sólo existen para taxónes de la península ibérica, y que incluyen preferentemente caracteres morfométricos (López-Sánchez et al. 1985 a,b, 1987). Sin embargo, el conocimiento morfológico y bionómico de los estados preimaginales, permite reconocer diferencias poblacionales intraespecíficas (Horsfall et al. 1970), postular hipótesis de inferencias filogenéticas (Stark y Szczytko 1982) o bien sugerir adaptaciones a hábitats particulares (Luff 1981), como ocu rre con tenebriónidos de ecosistemas áridos semiáridos, que presentan características adaptativas para la vida edáfica (Doyen y Tschinkel 1974).
Gyriosomus Guérin-Méneville, 1834 es un importante elemento endémico y erémico de la entomofauna epígea de los ecosistemas desérticos del norte de Chile (Pizarro-Araya y Jerez 2004, Pizarro-Araya y Flores 2004), y la mayoría de sus especies presentan irrupciones poblacionales relacionadas con El Niño Oscilación del Sur (ENOS) (Cepeda-Pizarro et al. 2005 a,b). La distribución del género abarca desde el norte de la Reserva Nacional Paposo (25°00S), hasta la precordillera de Rancagua (34º 11S). Gyriosomus kingi Reed, 1873 (Fig. 2e), es una especie endémica del Desierto Costero del Huasco (DCH) (Pizarro-Araya y Jerez 2004), cuyas poblaciones en año con fenómeno ENOS han llegado a representar el 74.4% de la diversidad total de artrópodos (Cepeda-Pizarro et al. 2005 a). A partir de prospecciones realizadas en el DCH en septiembre de 2002 (año ENOS), se recolectó gran número de especímenes de ambos sexos de G. kingi lo que permitió obtener oviposturas y larvas de primer estadio.
Dado que la estructura coriónica y morfología larvaria constituyen elementos de diagnóstico específico, adaptativos y filogenéticos, los objetivos de este trabajo son describir la morfología del huevo y primer estadio larvario de G. kingi, dar a conocer aspectos de su biología reproductiva, y determinar caracteres que permitan postular inferencias filogenéticas.
Materiales y métodos
En septiembre de 2002 se recolectaron parejas de G. kingi en el sector costero del Parque Nacional Llanos de Challe (28°01S; 71°03W), provincia de Huasco, iii Región, Chile. Los insectos se mantuvieron en cajas de crianza hasta la oviposición a temperatura de 17°C mínima y 24°C máxima y un fotoperíodo de 12 h luz y 12 h oscuridad. Fueron alimentados con Erodium cicutarium (L.) (Geraniaceae), Cristaria glaucophylla Cav. (Malvaceae) más un suplemento de dieta artificial.
Huevos y larvas de primer estadio fueron fijados en A.F.A caliente; las fotografías a microscopio electrónico de barrido fueron realizadas, previa deshidratación de huevos y larvas en batería de alcohol, secado a punto crítico y metalizado con oro, según la técnica utilizada por Pizarro-Araya et al. (2005). Para la descripción de la estructura y ornamentación del corión, se utilizó la nomenclatura seguida por Pizarro-Araya et al. (2005). Para la morfología larvaria se siguió a Keleynikova (1963), López-Sánchez et al. (1985 a,b, 1987) y Doyen (1988, 1993).
Resultados
Huevo (Fig. 1a): largo: 4.0 mm (ámbito: 3.7-4.3), ancho: 1.1 mm (ámbito: 1.0-1.2) (n= 10). Coloración blanca, forma elongada y polos redondeados; corión liso, sin aeropilas visibles a microscopía electrónica; el primer tercio del huevo está ornamentado con celdas subhexagonales cuyo ancho aproximado es 17.8 μm (αmbito: 14.8-22.6) (Fig. 1b). El área micropilar está ubicada sobre un levantamiento del polo apical con un diámetro de 27.2 μm (ámbito: 2731). El poro micropilar es poco visible ya que se encuentra cubierto por una secreción y alcanza un diámetro de 1.4 μm (ámbito: 0.5-2.5). La periferia de la micropila y del poro micropilar, presenta numerosas aeropilas con diámetros entre 1 μm y 2 μm (Fig. 1c).
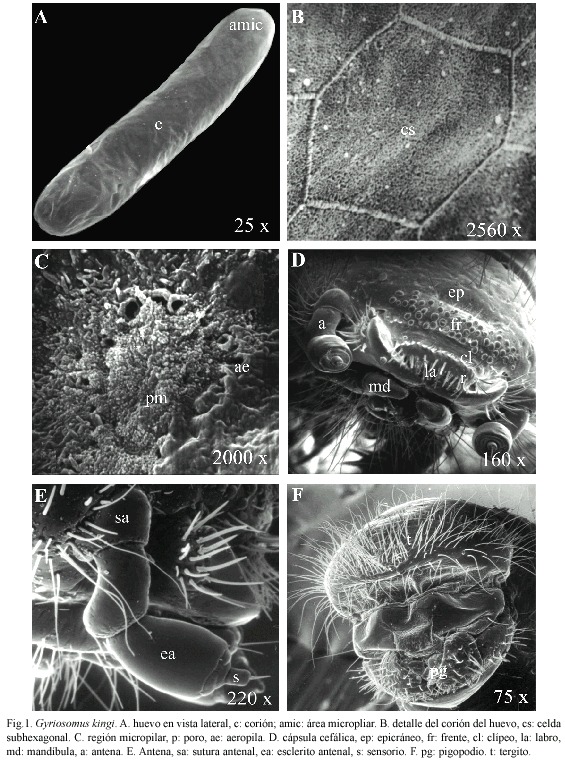
Larva de primer estadio: Largo 5.2 mm (ámbito: 3.5-6.2 mm) (n= 15). Oligópoda de cuerpo elongado, cilíndrico y con tegumento glabro y débilmente esclerificado a excepción de la cápsula cefálica. Coloración blanquecina. Cápsula cefálica (Cuadro 1), prognata, redondeada; epicráneo punteado, provisto de setas cortas. Sutura epicraneal no evidente, sutura frontal presente y rectilínea, sutura frontoclipeal distinta, ligeramente curvada. Región frontal dorsalmente algo deprimida, provista de numerosos tubérculos dispuestos en hileras horizontales, ocelos ausentes (Fig. 1d). Antena de tres segmentos, segundo segmento elongado, dos veces la longitud del primero, y provisto de un sensorio redondeado y una seta apical (Fig. 1e). Clípeo subrectangular y glabro; labro transverso, separado de la cápsula cefálica; cara dorsal con un par de sensilas y con un pecten formado por una hileras de setas gruesas y de forma lanceolada, cuatro setas a cada lado y un par de setas más finas en el margen externo; margen anterior levemente sinuado y provisto de setas largas y rígidas (Fig. 2a). Mandíbulas subtriangulares bien quitinizadas con dos dientes apicales de ápice obtuso; mola bien desarrollada, ovalada, cóncava y superficie lisa; porción basolateral de la superficie dorsal con un mechón de setas, largas y rígidas. Maxilas con palpos de tres segmentos, último segmento corto y de ápice redondeado; mala redondeada y provista con numerosas setas cortas y gruesas en el margen externo. Palpos labiales de dos segmentos (Fig. 2b). Tórax. Primer segmento torácico más largo que los siguientes y con el margen anterior engrosado (Cuadro 1). Patas protorácicas más desarrolladas que el segundo y tercer par; tarsungulus largo y curvado (Fig. 2c); margen interno de la tibia con tres espinas de aspecto lanceolado; margen anterior y superficie externa con numerosas espinas largas y delgadas, margen interno del fémur con una fila de espinas cortas, gruesas y de ápice obtuso (Fig. 2d). Abdomen. Nueve segmentos revestidos con pilosidad escasa y corta; espiráculos uniforos, segmento abdominal IX elongado; tergito recubierto de sedas largas y delgadas, pigopodio desarrollado (Fig. 1f).

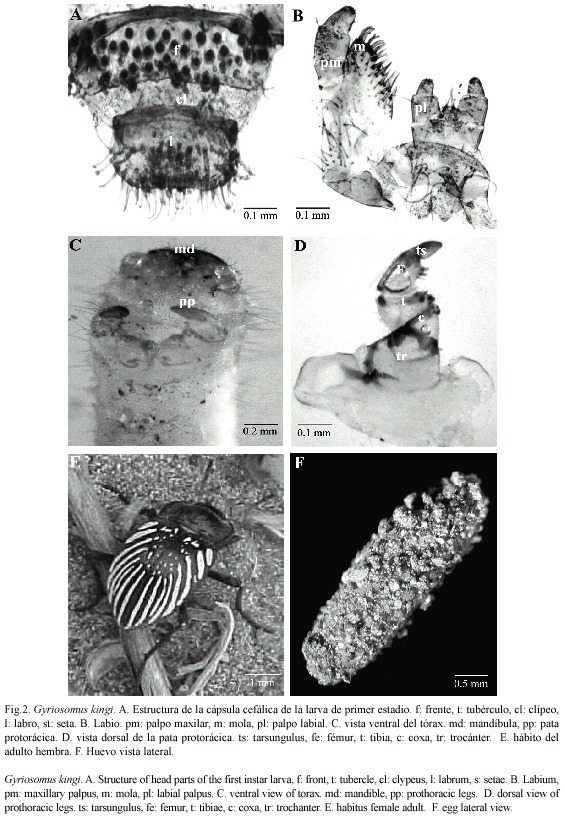
Bionomia: G. kingi (Fig. 2e), está asociada tróficamente a la vegetación arbustiva característica del DCH como Cristaria glaucophylla Cavanilles, 1799 (Malvaceae), Leucocoryne appendiculata Philippi, 1896 (Alliaceae), Adesmia littoralis Burkart, 1967 (Fabaceae) y Alstroemeria leporina Ehr.Bayer et Grau, 1982 (Alstroemeriaceae). Sin embargo, a partir de nuestras observaciones en terreno se estableció que las hembras presentan un marcado canibalismo y depredan larvas de Noctuidae (Lepidoptera). El período reproductivo se inicia a mediados de septiembre con la emergencia de adultos a los que es factible de encontrar desplazándose sobre el suelo o sobre las partes aéreas de plantas (e.g. Skytanthus acutus Meyen, 1834 (Apocynaceae), Nolana paradoxa Lindley, 1825 (Nolanaceae)). Son de hábitos diurnos y en las horas de mayor radiación solar se entierran o se esconden bajo piedras o arbustos. Las cópulas ocurren inmediatamente a la emergencia del estado de pupa y observaciones de laboratorio muestran que las hembras inician las oviposturas a finales de septiembre, ya sea en la superficie del sustrato arenoso, o bien enterrándose a una profundidad que varía entre 10 a 20 cm. Los huevos son colocados en grupos de siete a diez con un máximo de seis posturas por hembra. Los huevos no son visibles externamente ya que están revestidos por una película de arena que se adhiere al corión mediante un mucílago que se seca posteriormente (Fig. 2f).
Discusión
El corión del huevo representa una protección para el embrión contra el efecto de factores físicos y biológicos, permitiendo su sobrevivencia y por lo tanto está sometido a fuertes presiones de selección. En este sentido la microescultura reticulada del corión, representa para muchos insectos un mecanismo de defensa contra la pérdida de agua y las aeropilas permiten el intercambio respiratorio entre el huevo y el medio (Stark y Szczytko 1982). Los huevos además pueden incluir recubrimientos elaborados en base a materia fecal y/o alimento regurgitado (Nordell-Paavola et al. 1999).
Los huevos de Gyriosomus no presentan las características antes señaladas, sin embargo, el corión liso y sin aeropilas, está recubierto por una capa protectora de mucílago al momento de la ovipostura, sobre la cual se adhieren gránulos de sedimentos. Esta cubierta está constituida por gránulos de arcilla que representarían un aislamiento térmico frente a las marcadas oscilaciones de temperatura que se presentan en estos ambientes semidesérticos con suelos aridisoles (Rovira 1984).
El origen polifilético de los Tenebrionidae se refleja en una alta diversidad morfológica y etológica entre las tribus, géneros y especies; esta marcada variabilidad ha originado cambios sucesivos en los niveles taxonómicos de la familia (Aalbu et al. 2002, Bouchard et al. 2005). Sin embargo, estas modificaciones se han realizado excluyendo caracteres preimaginales. Al respecto, Doyen (1988) establece la importancia de incorporar estos caracteres para la confirmación taxonómica y sistemática de los taxónes de tenebriónidos, y proponer a partir de ellos inferencias filogenéticas. La larva de primer estadio de G. kingi presenta características morfológicas que permiten corroborar la inclusión de Gyriosomus dentro de Pimeliinae, la subfamilia más diversa de Tenebrionidae y cuyas especies se distribuyen en todas las regiones áridas y semiáridas del mundo (Watt 1974, Matthews 2000). Por otra parte, sus características morfo-adaptativas permiten relacionarla al tipo pedobionta señalado por Keleynikova (1963) y cuya característica más relevante es la presencia de patas protorácicas muy desarrolladas y con función cavadora.
Agradecimientos
Al personal del Laboratorio de Microscopía electrónica de la Dirección de investigación de la Universidad de Concepción. Raúl Briones del Departamento de Zoología, Facultad de Ciencias Naturales y Oceanográficas de la Universidad de Concepción, Chile; Gustavo E. Flores del instituto Argentino de investigaciones de las Zonas Áridas IADIZA-CRICYT, Argentina. Este trabajo fue financiado por los proyectos DIUC N° 203.113.061-1.0, Dirección de investigación, Universidad de Concepción, Concepción, Chile y DIULS N° 220.2.17, Dirección de investigación, Universidad de La Serena, La Serena. Chile.
Resumen
Gyriosomus es un importante género chileno de Tenebrionidae, habitante del desierto costero del país. En este ambiente, las especies de Gyriosomus son un componente erémico y endémico de la entomofauna epígea. A pesar de la importancia ecológica que estas especies tienen en el desierto chileno, poco se conoce respecto de la morfología de los estados preimaginales. En este trabajo se describe la morfología del huevo y de la larva de primer estadio de G. kingi Reed, 1873, y se entregan algunos antecedentes acerca de la bionomía de la hembra. Los huevos y las larvas fueron obtenidos a partir de parejas mantenidas en condiciones de laboratorio. Los adultos fueron capturados durante la primavera del año 2002, en sitios costeros del Parque Nacional Llanos de Challe, localizado en la frontera sur del Desierto de Atacama (28°01S; 71°03W). El primer tercio del corión del huevo presenta celdas subhexagonales sin aeropilas. La larva es oligópoda y elongada. Su cápsula cefálica está fuertemente quitinizada y sus patas protorácicas son más largas y más gruesas que las restantes. Según observaciones de laboratorio, cada ovipostura incluye de siete a diez huevos cada vez, con un máximo de seis oviposturas por hembra. Los huevos son enterrados en el suelo, recubiertos con un mucílago que genera una película de arena-arcilla adherida al corión.
Palabras clave: Tenebrionidae, Pimeliinae, ontogenia de insectos, fauna del suelo desértico, desierto costero chileno.
Referencias
Aalbu, R.L., C.A. Triplehorn, J.M. Campbell, K.W. Brown, R.E. Somerby & D.B. Thomas. 2002. Tenebrionidae Latreille 1802. Chapter 106, p. 463-509. In R.H. Arnett., M.C. Thomas, P.E. Skelley & H. Frank (eds.). American Beetles. CRC, Boca Raton, Florida, EEUU. [ Links ]
Artigas, J. 1994. Entomología económica. Ediciones Universidad de Concepción, Concepción. Chile. 943 p. [ Links ]
Artigas, J. & P. Brañas-Rivas. 1973. Descripción de la larva y pupa de Praocis curta Solier (Coleoptera: Tenebrionidae). Bol. Soc. Biol. Concepción. XLVI: 64-74. [ Links ]
Bouchard, P., J.F. Lawrence, A.E. Davies & A.F. Newton. 2005. Synoptic classification of the world Tenebrionidae (insecta: Coleoptera) with a review of family-group names. Ann. Zool. 55: 499-530. [ Links ]
Cekalovic, T. & A. Quezada. 1973. Descripción de la larva, pupa y distribución geográfica de Emmallodera multipunctata curvidens Kulzer, 1955 (Coleoptera: Tenebrionidae). Anal. inst. Patag. 4: 417-422. [ Links ]
Cekalovic, T. & E. Morales. 1974. Descripción de la larva y pupa de Oligocara nitida Solier, 1848 (Coleoptera: Tenebrionidae). Bol. Soc. Biol. Concepción. XLVIII: 173-177. [ Links ]
Cepeda-Pizarro, J.G., H. Vásquez, H. Veas & G. Colon. 1996. Relaciones entre tamaño corporal y biomasa de Tenebrionidae (Coleoptera) de la estepa costera del margen meridional del desierto chileno. Rev. Chil. Hist. Nat. 69: 67-76. [ Links ]
Cepeda-Pizarro, J., J. Pizarro-Araya & H. Vásquez. 2005a. Composición y abundancia de artrópodos del Parque Nacional Llanos de Challe: impactos del ENOS de 1997 y efectos del hábitat pedológico. Rev. Chil. Hist. Nat. 78: 635-650. [ Links ]
Cepeda-Pizarro, J., J. Pizarro-Araya & H. Vásquez. 2005b. Variación en la abundancia de Arthropoda en un transecto latitudinal del desierto costero transicional de Chile, con énfasis en los tenebriónidos epígeos. Rev. Chil. Hist. Nat. 78: 651-663. [ Links ]
Doyen, J.T. 1988. Descriptions of some phylogenetically important larvae of Tenebrionidae (Coleoptera). Coleop. Bull. 42: 285-301. [ Links ]
Doyen, J.T. 1993. Cladistic relationships among Pimeliinae Tenebrionidae (Coleoptera). J. N. y. Entomol. Soc. 101: 443-514. [ Links ]
Doyen, J.T. & W.F. Tschinkel. 1974. Population size, microgeographic distribution and habitat separation in some tenebrionid beetles (Coleoptera). Ann. Entomol. Soc. Am. 67: 617-626. [ Links ]
Flores, G.E. 1997. Revisión de la tribu Nycteliini (Coleoptera: Tenebrionidae). Rev. Soc. Entomol. Argent. 56: 1-19. [ Links ]
Flores, G.E. 2000. Cladistics analysis of the Neotropical tribe Nycteliini (Coleoptera: Tenebrionidae). J. N. y. Entomol. Soc. 108: 13-25. [ Links ]
Guérin-Méneville, F.E. 1834. Matériaux pour une classification des Mélasomes. Magazine du Zoologie 3e. année: 1-39. [ Links ]
Horsfall, W.R., F.R. Voorhess & E.W. Cupp. 1970. Eggs of floodwater mosquitoes. XIII. Chorionic sculpturing. Ann. Entomol. Soc. Am. 63: 1709-1716. [ Links ]
Keleynikova, S.i. 1963. Larval types of palaeartic darkling beetles (Coleoptera, Tenebrionidae). Entomol. Rev. 42: 292-296. [ Links ]
Kulzer, H. 1954. Neunter Beitrag zur Kenntnis der Tenebrioniden (Col.) Eine Studie über die Tribus Nycteliini. Ent. Arb. Mus. G. Frey. 5: 14-267. [ Links ]
Kulzer, H. 1959. Neue Tenebrioniden aus Südamerika (Col.) 18 Beitrang zur Kenntnis der Tenebrioniden. I Die Gattung Gyriosomus Guérin. (Nycteliini). Ent. Arb. Mus. G. Frey. 10: 523-547. [ Links ]
López-Sánchez, S., A. de los Santos & C. Montes. 1985a Estudio morfológico de la forma larvaria de Tentyria placyteps Stev., (1829). Eos. LXI: 173-182. [ Links ]
López-Sánchez, S., A. de los Santos & C. Montes. 1985b Estudio morfológico de los estados preimaginales de Micrositus ulyssiponensis Germ. 1824 (Coleoptera: Tenebrionidae). An. Biol. Univ. Murcia. 3: 95-102. [ Links ]
López-Sánchez, S., A. de los Santos & C. Montes. 1987 Estudio morfológico de los estados preimaginales de Pimelia costata Waltl. 1835 (Coleoptera: Tenebrionidae). Nouvelle Revue d´entomologie. Tome 4, Fascicule 3: 299-311. [ Links ]
Luff, M.L. 1981. Diagnostic characters of the eggs of some Carabidae (Coleoptera). Ent. Scand. Suppl. 15: 317-327. [ Links ]
Nordell-Paavola, A., S. Nokkala, S. Koponen & C. Nokkala. 1999. The utilization of chorion ultrastructure and chorion polypeptide analysis in recognizing taxonomic units in North European Galericini (Col., Chrysomelidae), p. 95-104. In Cox M.L. (ed.) Advances in Chrysomelidae Biology 1. Backhuys Leiden, Holanda. [ Links ]
Matthews, E.G. 2000. Origins of Australian arid-zone tenebrionid beetles. invertebr. Taxon. 14: 941-951. [ Links ]
Peña, L.E. 1985. Revisión del género Psectrascelis Fairm. (Coleoptera: Tenebrionidae). Rev. Chil. Ent. 12: 15-51. [ Links ]
Peña, L.E. 1995. Revisión del género Physogaster Guérin, 1834 (Coleoptera: Tenebrionidae: Physogasterini). Gayana Zool. 59: 119-130. [ Links ]
Pizarro-Araya, J. & G.E. Flores. 2004. Two new species of Gyriosomus Guérin-Méneville from the Chilean coastal desert (Coleoptera: Tenebrionidae: Nycteliini). J. N. y. Entomol. Soc. 112: 121-126. [ Links ]
Pizarro-Araya, J. & V. Jerez. 2004. Distribución geográfica del género Gyriosomus Guérin-Méneville, 1834 (Coleoptera: Tenebrionidae): Una aproximación biogeográfica. Rev. Chil. Hist. Nat. 77: 491-500. [ Links ]
Pizarro-Araya, J., V. Jerez & J. Cepeda-Pizarro. 2005. Descripción de huevos y larvas de primer estadio del género Gyriosomus Guérin-Méneville, 1834 (Coleoptera: Tenebrionidae: Nycteliini). Gayana 69: 277-284. [ Links ]
Reed, E.C. 1873. Descriptions of new species of Coleoptera from Chili. Ent. Monthl. Mag., Vol. 9: 207-209. [ Links ]
Rovira, A. 1984. Geografía de los Suelos. Colección Geográfica de Chile, inst. Geográfico Militar, Santiago, Chile. 180 p. [ Links ]
Stark, B.P. & S.W. Szczytko. 1982. Egg morphology and phylogeny in Pteronarcyidae (Plecoptera). Ann. Entomol. Soc. Am. 75: 519-529. [ Links ]
Watt, J.C. 1974. A revised subfamily classification of Tenebrionidae (Coleoptera). New Zealand J. Zool. 1: 381-452. [ Links ]

