










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.54 n.3 San José Sep. 2006
Scanning electron microscopy of the four larval instars of the Dengue fever vector Aedes aegypti (Diptera: Culicidae)
Stefan Schaper1 & Francisco Hernández-Chavarría2
1 Institute for Zoology, Tierärztliche Hochschule Hannover, Germany and Universidad Nacional, Heredia, Costa Rica.
2 Centro de Investigación en Estructuras Microscópicas and Facultad de Microbiología, Universidad de Costa Rica, San José, Costa Rica; hchavarr@cariari.ucr.ac.cr
Received 09-IV-2003. Corrected 16-IX-2005. Accepted 23-III-2006.
Abstract: Aedes aegypti is the main insect vector of Dengue fever and dengue hemorrhagic fever/dengue shock syndrome and represents the only vulnerable element in the control of this disease. Therefore, the identification and quantification of this mosquito is an important task; however, the majority of taxonomic keys are based on the 4th larval instar. For that reason, this study describes the four larval instars of A. aegypti using scanning electron microscopy. Morphological changes during larval development were observed at the pecten, comb scales and the ventral brush of the abdominal segment X; however, the 3rd and 4th instars showed similar structures with only a slight variation. The structures described in this study will be helpful in the identification of the four instars of A. aegypti, a fundamental task for comprehending the natural history of dengue mainly in new territories affected. Rev. Biol. Trop. 54 (3): 847-852. Epub 2006 Sept. 29.
Key words: Aedes aegypti, larval development, morphology, scanning electron microscopy.
Dengue is an arthropod-borne viral disease whose main vector is the mosquito Aedes aegypti. This virus is the etiologic agent of dengue fever and dengue hemorrhagic fever/dengue shock syndrome, whose incidence has shown a constant increase in the last few decades. It is estimated that up to 80 million people are infected annually (Pinheiro and Corber 1997). A. aegypti was reintroduced in Costa Rica at the beginning of the 1990s and the first outbreak of dengue occurred in 1993.
A aegypti has adapted its biological behavior to the human environment, making use of the water that collects in discarded plastic containers and used tires as breeding sites where the mosquito oviposits. From the egg to the adult there are four larval instars and one pupal stage. The larvae change considerably in size during their development; the first instar is only about 1 mm long, whereas the fourth instar is approximately 8 mm long. However, most identification keys are based exclusively on the characteristics of the 4th instar (Clark-Gil and Darsie 1983).
The retrospective studies of specimens preserved in ethanol from water collected at artificial breeding sites are very important in the comprehension of the natural history of dengue in new territories. That research includes identification and differentiation of mosquito larvae found in this same habitat. However, one problem encountered is that it is not always possible to identify all larvae in the sample because many of the individuals are younger instars, which are usually not included in taxonomic keys (Clark-Gil and Darsie 1983).
Scanning electron microscopy (SEM) has not been used extensively in describing the external morphology and fine structure of A. aegypti larvae, although it is frequently employed in the observation of the eggs (Linley 1989a,b, Linley and Clark 1989, Linley and Craig 1994, Service et. al 1997) or adult mosquitoes (Sutcliffe 1994, Bowen 1995). The structures of taxonomical interest that are used in taxonomic keys include the siphon, anal segment and head capsule. Therefore, the aim of this study was a SEM description and a comparison of the structures with taxonomic interest in all instars of A. aegypti.
Materials and methods
The larvae of A. aegypti were obtained from a one year-old laboratory culture (started with specimens from Heredia, Costa Rica). At least five individuals from each larval instar were fixed in 2.5% glutaraldehyde, postfixed in osmium tetraoxide, dehydrated, critical point dried, ion sputtered gold, and examined using a SEM (Hitachi S-570). Observations were made mainly on the head capsule, siphon, segments VIII and X and the mouth brushes.
Results
The structures were found to be similar in all instars, showing equivalent distributions and relative sizes, including the terminal structures of the siphon and setae of the head capsule. The main structural differences were observed in the lateral palatal brushes, comb scales, and pecten.
Lateral palatal brushes of the 1st and 2nd instars appeared as conglomerates of uniformly shaped filaments. Thus, the palpi of both the palatum and maxillae are evident because the filaments appear as slender and long structures and the palatum with its setae are easily differentiable (Fig. 1). This contrasts markedly with the equivalent structures of the 3rd and 4th instars, in which the arrangement of setae is more complex, scattered in a jumbled fashion. It is difficult to determine where each one originates because the tips of the palatum filaments appear to be intermingled with filaments from other mouth structures and the palatum appears to be wider than in the first two instars (Fig. 2). The palatal brushes of the first two instars showed a relatively straight structure with a row of uniform teeth (Fig. 3). In the 3rd and 4th instars, the setae showed a more complex structure; for example, in the maxillary palpi, the tip of each seta has a distal rake-like structure of five to six pointed teeth (length 8.85±1.4 µm) and is folded approximately 90° with respect to the other undifferentiated teeth on the long stalk of the setae (Fig. 4).
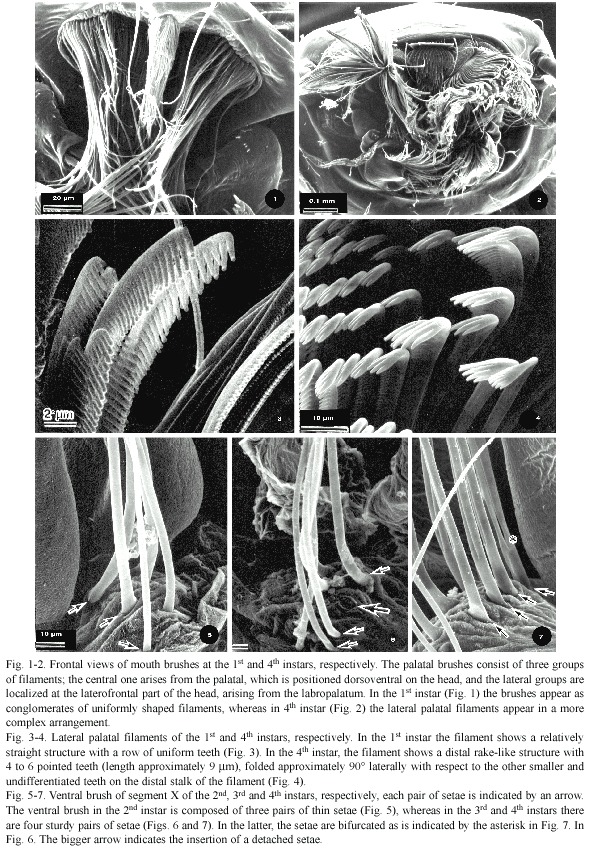
The development of the ventral brush of segment X showed morphological changes. The ventral brush in the 2nd instar was composed of three pairs of thin setae (Fig. 5), although in the 1st instar, only two pairs of setae were observed (not shown). The 3rd and 4th instars showed four pairs of sturdy setae as indicated by the arrows placed at their bases in Fig. 6 and 7; however, in Fig. 6 one of the setae was detached from the specimen (pointed out by a larger arrow). In the 4th instar the setae are bifurcated as is pointed out by an asterisk in Fig. 7.
The number of comb scales and their complexity increased from the 1st to the 4th instar. The first instar had only five scales, each one bearing a fringe of 19-21 pointed and uniformly shaped spines (Fig. 8). The 2nd, 3rd and 4th instars had six to seven, eight, and ten comb scales, respectively (Fig. 9, 10 and 11). In addition, their structures were more complex than those of the first instar, because the comb scales of the later instars showed one long median denticle with a slightly curved and more prominent apex than the others and had a varying number of subapical denticles. In contrast, the 2nd instar, each comb scale had 12-14 subapical denticles, whereas in the 3rd instar the number of subapical denticles was 14-17, and in the 4th there were 10-14 subapical denticles.
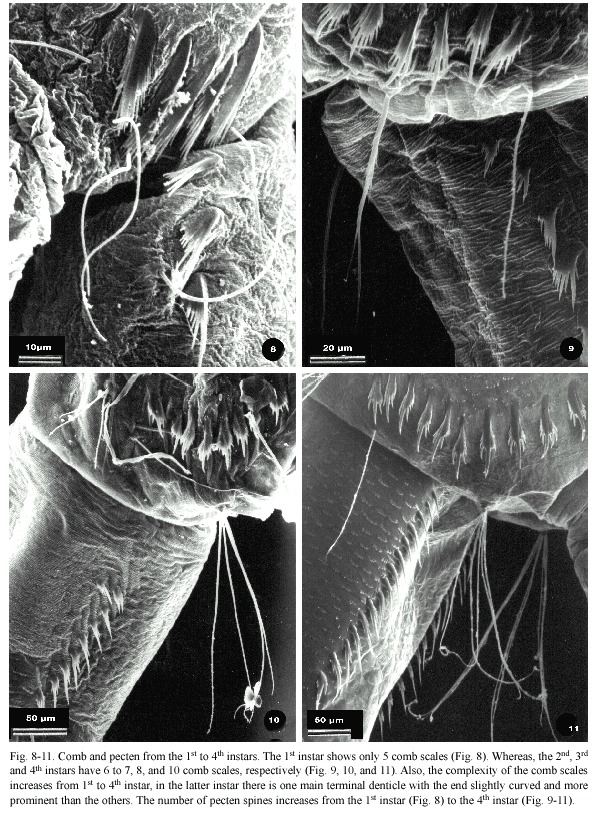
The pecten also showed modifications at different larval stages with regards to the number and complexity of their spines. In one specimen analyzed at 1st instar the pecten was formed by only two pecten spines, bearing an arrangement of seven small denticles, while the other instars usually showed more than ten denticles. In the pecten spines at the 2nd instar one
of the denticles was more prominent with four to five subapical denticles. This pattern was more evident in the mature instars; thus, the 3rd instar had 11 spines with two to four subapical denticles, and the 4th instar had 15-16 spines, each composed of one prominent denticle and two to three subapical denticles (Fig. 8-11).
Discussion
The mouth brushes of A. aegpti showed the most evident transformations during larval development from the 1st to the 4th instar. One modification was an increase in the number of lateral palatal brush filaments. In the 1st and 2nd instars, individual filaments of the palatum and maxillary palpi were evident because each structure had fewer filaments than those of the 3rd and 4th instars. The filaments of the former instars were also very simple, whereas in the later instars (3rd and 4th) they appeared as a tangle of long structures, obstructing the view of individual mouth parts. Furthermore, the filaments of these brushes have a terminal rake-like structure, composed of four to five big teeth, located at an angle of approximately 90° with respect to the rest of the smaller teeth in the filament. This development may be important in the feeding of larvae since it could provide for better grasping of food particles, including bacterial films and protozoa (Rashed and Mulla 1990). The development of these filaments might be correlated with a change in diet during larval maturation (Rashed and Mulla 1990). It is assumed that 1st and 2nd instar larvae feed more on bacteria, whereas 3rd and 4th instars ingest large organic particles and can be seen feeding on surface films in culture containers. Dahl et al. (1993) observed differences in food selection among different instars; however, they found that the younger instars showed preference for larger particles. For this reason, the morphological change is unlikely to be related to a behavioral change during larval development; nevertheless, the big filaments on the lateral palatal brush could enable the larvae to capture and feed on proportionally larger food items.
The structural modifications of the comb and pecten are also important due to their taxonomical interest. There were fewer comb scales in the 1st and 2nd instars; however, they bear the same appearance and distribution as in later instars. At the high resolution of the SEM, the comb scales of the pecten at the 1st instar appeared as a fringe of small denticles; however, using light dissection microscopy, this structure appeared as a single spine. The development of the pecten showed an increase in the number and complexity of the pecten spines.
Size comparisons of the different structures analyzed were not done because those dimensions may have been altered during processing, particularly during the critical point drying, as has been previously described by various authors (Eskelinen and Saukko 1983, Beckett et al. 1984).
In summary, structural changes during larval development are evident, mainly in the mouth parts. The 3rd and 4th instars showed similar structures with only a slight variation in the number of elements. The structures described in this paper should be helpful for more accurate identification of any of the four instars of A. aegypti, a significant task when retrospective analysis of preserved water samples containing different immature forms of mosquitoes is needed.
Acknowledgments
The authors are grateful to Jorge D. García and Belinda Dick for their suggestions and critical review of the manuscript. We also thank Leda Muñoz for her support. This study was funded by the Vicerrectoría de Acción Social, the Vicerrectoría de Investigación de la Universidad de Costa Rica, and PNUD.
Resumen
Aedes aegypti es el principal insecto vector de la fiebre del dengue y del dengue hemorrágico/síndrome del choque por dengue y es el único elemento atacable para el control de esta virosis. La identificación y cuantificación de éste es una tarea importante; no obstante, la mayoría de las llaves taxonómicas se basan en el cuarto estadio larval. Por esta razón, en este trabajo se describen los cuatro estadios larvales de A. aegypti los cuales fueron examinados mediante microscopia electrónica de rastreo. Los cambios morfológicos ocurridos durante el desarrollo larval fueron observados en el pecten, las escamas del peine, el cepillo ventral del décimo segmento. El 3ero y 4to estadios larvales mostraron estructuras similares con sólo ligeras variaciones. Las estructuras descritas en este artículo permiten identificar cualquiera de los cuatro estadios larvales de A. aegypti, lo cual representa una tarea importante en la comprensión de la historia natural del dengue en los nuevos territorios afectados.
Palabras clave: Aedes aegypti, desarrollo larval, morfología, microscopia electrónica de rastreo.
References
Beckett, A., N.D. Read & R. Porter 1984. Variation in fungal spore dimensions in relation to prepraratory techniques for light microscopy and scanning electron microscopy. J. Microsc. 136: 87-95. [ Links ]
Bowen, M.F. 1995. Sencilla basiconia (grooved pegs) on antennae of female mosquitoes: electrophysiology and morphology. Entomol. Exp. Appl. 77: 233-238. [ Links ]
Clark-Gil, S. & R.F. Darsie. 1983. The Mosquitoes of Guatemala, their identification and binomics. Mosquito Syst 15: 151-284 [ Links ]
Dahl, C., G. Sahl, J. Grawe, A. Johannisson & H. Amenus. 1993. Differential particle uptake by larvae of three mosquito species (Diptera: Culicidae). J. Med. Entomol. 30: 537-543. [ Links ]
Eskelinen, S. & P. Saukko. 1983. Effects of glutaldehyde and critical point drying on the shape and size of erythrocytes in isotonic and hypotonic media. J. Microsc. 130: 63-71. [ Links ]
Linley, J.R. 1989a. Scanning electron microscopy of the egg of Aedes (Protomacleaya) triseriatus (Diptera: Culicidae). J. Med. Entomol. 26: 474-478. [ Links ]
Linley, J.R. 1989b. Comparative fine structure of the eggs of Aedes albopictus, Ae. aegypti and Ae.bahamensis (Diptera: Culicidae). J. Med. Entomol. 26: 510-521. [ Links ]
Linley, J.R. & G.G. Clark. 1989. Egg of Aedes (Gymnometopa) mediovittatus (Diptera: Culicidae). J. Med. Entomol. 26: 252-255. [ Links ]
Linley, J.R. & G.B. Jr Craig. 1994. Morphology of long- and short-day eggs of Aedes atropalpus and A. epactius (Diptera: Culicidae). J. Med. Entomol. 31: 855-867. [ Links ]
Pinheiro, F.P. & S.J. Corber. 1997. Global situation of dengue and dengue haemorrhagic fever, and its emergence in the Americas. World Health Stat. Q. 50: 161-169. [ Links ]
Rashed, S.S. & M.S. Mulla. 1990. Comparative functional morphology of the mouth brushes mosquito larvae (Diptera: Culicidae). J. Med. Entomol. 27: 429-439. [ Links ]
Service, M.W., D. Duzak & J.R. Linley. 1997. SEM examination of the eggs of five British Aedes species. J. Am. Mosq. Control Assoc.13:47-65. [ Links ]
Sutcliffe, J.F. 1994. Sensory bases of attractancy: Morphology of mosquito olfactory sencilla: a review. J. Am. Mosq. Control Assoc. 10: 309-315. [ Links ]

