










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.53 n.1-2 San José Jun. 2005
Trophic interactions within the microbial food web in a tropical floodplain lake (Laguna Bufeos,Bolivia)
Danny Rejas 1,2 ,Koenraad Muylaert 3 & Luc De Meester 1
1 Laboratory of Aquatic Ecology, Katholieke Universiteit Leuven, Ch. de Bériotstraat 32, B-3000 Leuven, Belgium
2 Unidad de Limnología y Recursos Acuáticos, Universidad Mayor de San Simón, Casilla 471, Cochabamba, Bolivia; drejas@supernet.com.bo
3 University Gent, Department Biology, Section Protistology & Aquatic Ecology, Krijgslaan 281-S8, 9000 Gent, Belgium
Received 11-VI-2003. Corrected 07-VII-2004. Accepted 07-VII-2004.
Abstract
Whether the primary role of bacterioplankton is to act as "remineralizers" of nutrients or as direct nutritional source for higher trophic levels will depend on factors controlling their production and abundance. In tropical lakes, low nutrient concentration is probably the main factor limiting bacterial growth, while grazing by microzooplankton is generally assumed to be the main loss factor for bacteria. Bottom-up and top-down regulation of microbial abundance was studied in six nutrient limitation and dilution gradient-size fractionation in situ experiments. Bacteria, heterotrophic nanoflagellates (HNF), ciliates and rotifers showed relatively low densities. Predation losses of HNF and ciliates accounted for a major part of their daily production, suggesting a top-down regulation of protistan populations by rotifers. Phosphorus was found to be strongly limiting for bacterial growth, whereas no response to enrichment with Nitrogen or DOC was detected. HNF were the major grazers on bacteria (g=0.43 d-1 ), the grazing coefficient increased when ciliates were added (g=0.80 d-1 ) but decreased when rotifers were added (g=0.23 d-1 ) probably due to nutrient recycling or top-down control of HNF and ciliates by rotifers. Rev. Biol. Trop. 53 (1-2):85-96. Epub 2005 Jun 24.
Key words: Bacterioplankton, microzooplankton, grazing, nutrient, recycling, tropical lake.
Bacteria are thought to play a significant role in the flow of energy and matter through pelagic food webs. First, bacteria are remineralizers of dissolved and particulate organic matter and release inorganic nutrients that can be assimilated by the autotrophic phytoplankton. Second, they convert dead organic matter into living biomass and form a direct source of energy and carbon for higher trophic levels (Elser et al.1995). The major consumers of bacteria are protists like heterotrophic nano-flagellates (HNF) and ciliates, but metazoans like rotifers and cladocerans may also feed on bacteria. The significance of bacteria as contributors of energy and carbon to higher trophic levels in aquatic food webs, however, is controversial (Sanders et al. 1989).
Nutrient to carbon ratios in dissolved or particulate detritus tend to be significantly lower than nutrient to carbon ratios in bacterial cells. Therefore, although bacteria are important reminarelisers of nutrients in pelagic ecosystems, they often take up dissolved inorganic nutrients from the medium in order to sustain growth and to be able to respire detrital organic matter. Studies of bacterial mineral nutrition have suggested that, because of their large surface area to volume ratio, bacteria are excellent competitors with phytoplankton for nutrient elements such as nitrogen and phosphorus (Elser et al.1995) and thus can serve as nutrient "sinks " in aquatic ecosystems (Pomeroy and Wiebe 1988).
The microzooplankton size category (<200 µm) is composed of a diverse assemblage, including protozoa (HNF and ciliates), rotifers and naupliar stages of copepods. A large proportion of the bacterioplanktonic production is believed to be removed by predation by HNF and occasionally ciliated protozoa or mixotrophic algae (Sherr et al. 1986, Sanders et al. 1989, 1992). In natural seawater, feeding by HNF seems to be the main factor controlling bacterial densities, keeping them in relatively low and constant densities from 105 to 107 cells ml-1 (Jiménez-Gómez et al. 1994). Ciliates, on the other hand, are thought to dominate bacterivory in environments rich in detritus (Neill 1994). On their turn, ciliates can also feed on heterotrophic nanoflagellates. Rotifers represent an important part of the zooplankton biomass in most limnetic ecosystems and have been reported to have a variety of trophic modes including bacterivory, herbivory and carnivory (Sanders and Wickham 1993). Their food size spectrum (<1-20 µm) includes the main components of the microbial food web (Arndt 1993).
Microzooplankton may play an important role in the transfer of energy and matter because of its "trophic repackaging" of bacteria into larger "animal" cells that can be captured by zooplankton (Sherr and Sherr 1988). The importance of bacterial production and microbial consumers to macrobial food webs is still not resolved, especially in fresh water environments. Many authors seem to agree that high metabolic rates dissipate almost all microbial organics prior to consumption by macrobes and that bacteria and their microbial consumers serve mostly to remineralize inorganic nutrients subsequently used by primary producers (Currie 1990). In this view, bacterial and protists biomasses may provide emergency rations or casual by-catch for suspension feeders, except under "unusual "conditions (e.g. detritally driven ecosystems). The opposing perspective suggests that major uncertainties still remain, particularly at small spatial and temporal scales (e. g. hotspots of production, mutualistic facilitation) and that productive freshwaters may be especially conductive to high fluxes of microbial production to higher trophic levels, despite apparent low efficiency (Neill 1994).
Since the development of the microbial loop concept in the 1980s, large research efforts have been devoted to the study of bacteria, HNF and ciliates in a variety of aquatic ecosystems. Most studies, however, were carried out in the marine environment or in lakes at high latitudes. Very few studies on the microbial food web have been done in the tropics (Schmidt 1969, Rai and Hill 1984, Anesio et al. 1997, Erikson et al. 1998). Here we report the functioning of the microbial food web in a tropical floodplain lake, Laguna Bufeos (Bolivia). The aims of this study were to estimate the densities of microplanktonic organisms (bacteria, HNF, ciliates and zooplankton) and to contribute to the understanding of the factors controlling these densities. We carried out experiments to estimate growth rates and grazing/predation losses among the different trophic levels within the microbial food web (top-down control), and to determine the nutrient limitation of bacterial growth (bottom-up control)under experimental conditions.
Material and methods
Study site: Experiments were carried out in Laguna Bufeos, an oxbow lake in the flood-plain of the Ichilo River (16º 43 S -64º 70 W) at 174 m above sea level in the province Carrasco, Department of Cochabamba, Bolivia. Average depth of the lake is 2.36 -4.35 m during low-water and high-water season, respectively (Maldonado et al.1996). The climate is tropical, with mean annual temperatures exceeding 25 °C and a yearly precipitation of 2000-2500 mm. There is a short dry season of ome to four months during the period from July to September. Temperature variations are very small, except during some days in the dry season, when sub-Antarctic winds can cause minimum temperatures of 14° C. Mean Secchi depth is 61 cm and the zooplankton assemblage is composed of rotifers (86% in numbers) of the genus Brachionus and Polyarthra, cladocerans (7%) of the genus Moina and Bosminopsis, and copepods (7%) mainly cyclopoids in young stages (Acosta unpubl.). All experiments were performed between September 23 and 2, 1998.
Theoretical background: Two techniques were applied to investigate trophic interactions among components of the microbial food web. The first technique used was size fractionation, in which prey organisms are separated from their predators by filtration over a mesh or filter that retains all predators but allows all prey cells to pass through (Zimmermann 1996). Growth rates (k ) of the prey organisms are estimated from changes in abundance in the predator-free filtrate assuming an exponential growth response:
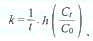
where Ct and Co are prey densities in the filtrate at the end and the beginning of the incubation, respectively, and t is the incubation time. The difference in growth response of the prey organisms in the filtered and unfiltered water corresponds to the grazing rate
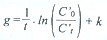
where C t and C o are prey densities in the unfiltered water at the end and the beginning of the incubation period, respectively.
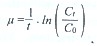
where Ct and Co are prey densities in the filtrate at the end and the beginning of the incubation period, respectively, and t is the incubation time.
Once growth and grazing rates k and g are known, one can calculate doubling per day = k potential production
ln2
Pp= t(Co · ek) - Co; actual production 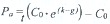 ; initial standing stock grazed d-1 (%) =
; initial standing stock grazed d-1 (%) =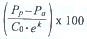
and potential production grazed d-1 (%) =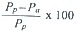 using equations reported by Gifford (1988).
using equations reported by Gifford (1988).
Experimental setup: Bacterial nutrient limitation: In order to test for nutrient limitation of bacteria, nutrient addition assays were set up. The water used for the assays was filtered through 2 µm polycarbonate membrane filters in order to reduce abundance of bacterial grazers. Four treatments were prepared: (1) addition of inorganic phosphorus (2.6 µM Na3 PO4), (2) addition of inorganic nitrogen (11 µM NH4Cl), (3) addition of organic matter as glucose (0.2 mg l-1 ), and (4) addition of all the former nutrients. Three replicates for each treatment were prepared. Filtered water was incubated in situ in 100 ml polycarbonate bottles for a period of 24 hr. Three replicate samples of the 2 µm filtered water were collected at the beginning of the incubation to determine initial bacterial numbers. From each treatment, a bacteria sample was taken at the end of the incubations.
Growth and grazing losses of bacteria: Three parallel dilution experiments were carried out to measure growth rates of bacteria and grazing rates on bacteria by grazers belonging to different size-fractions. First, water was filtered over nylon meshes with pore sizes of 10, 30 and 200 µm to include only grazers from certain size-fractions.The < 10 µm fraction was assumed to contain mainly HNF, the < 30 µm fraction was assumed to contain HNF and ciliates, while the < 200 µm fraction was assumed to contain HNF, ciliates and also metazoan microzooplankton like rotifers and copepod nauplii. To prepare the diluent, water was filtered over 0.2 µm pore size filters (MF Millipore) to remove bacteria as well as their grazers. Water of the three size-fractions (< 10, <30 and < 200 µm) was then diluted with 0.2 µm filtered water to obtain dilutions of 1, 0.8, 0.6, 0.4, 0.2 (fraction undiluted water to diluent). Two replicates were prepared for each dilution. Three replicate samples of the diluent and the undiluted water of the three size-fractions were collected at the beginning of the experiment. Initial concentrations of bacteria in diluted treatments were estimated based on the dilution factor and bacterial densities in undiluted water, assuming no bacteria were present in the diluent. Treatments were incubated in situ in 250 ml polycarbonate bottles at 30 cm depth and subsampled for bacteria after 24 hr of incubation.
Growth and grazing losses of HNF and ciliates: Growth rates and grazing losses of HNF and ciliates were measured by size-fractionation as well as an adapted version of the dilution method. For the fractionation experiment, lake water was sequentially filtered through nylon meshes of 200, 30 and 10 µm pore sizes in an attempts to remove macrozooplankton (200 µm), rotifers and copepod nauplii (30 µm), and ciliates (10 µm), respectively. Triplicate incubations of the different size-fractions were incubated in situ in 250 ml polycarbonate bottles during 24 hr. Treatments were subsampled for HNF and ciliates (only in the < 200 µm size-fraction) at the beginning and the end of the incubation. Rotifer growth was estimated in parallel incubations in triplicate 1 l bottles to obtain sufficiently large volume for enumeration of rotifers.
For the dilution experiments, lake water was first filtered over a 200 µm nylon mesh to remove macrozooplankton. Part of the water was then filtered through 2 µm pore size polycarbonate membrane filters, which remove all HNF and ciliates but do not retain picoplankton and bacteria. Duplicate series of dilutions (with fractions of undiluted water 1, 0.8, 0.6, 0.4 to 0.2) were prepared with the 200 µm filtered water and the 2 µm filtered water (as diluent). These dilutions were incubated in 100 ml diffusion chambers. These diffusion chambers were made by closing 100 ml bottles with 45 mm diameter, 2 µm pore size membrane filters. The use of diffusion chambers should allow for concentrations of picoplankton and bacteria to remain constant in all treatments.
The treatments were incubated in situ in the lake during 24 hr. 200 µm filtered water was sampled at the beginning of the experiment in order to estimate initial concentrations of HNF and ciliates in undiluted water. Initial concentrations in diluted treatments were estimated based on the dilution factor and abundances in the undiluted water, assuming no HNF and ciliates were present in the diluent. All treatments were sampled for HNF and ciliates at the end of the incubation.
For all experiments, in situ incubations were carried out at a fixed depth of 30 cm, started at noon and lasted 24 hr. All material used was previously washed with 10% HCl and rinsed three times with deionized water.
Analysis of samples: Bacterial abundance was determined by epifluorescence microscopy. Samples of 50 ml were collected and fixed with 0.2 µm filtered formalin to a final concentration of 3%. 2-10 ml subsamples were stained with Acridine Orange, filtered onto black 0.2 µm polycarbonate membrane filters and examined with a Zeiss Axioscop 50 epifluorescence microscope (blue excitation, 1000X magnification). At least 200 bacteria per sample were counted in random fields. HNF and ciliate samples (100 ml) were fixed with the Lugol-formalin-thiosulphate method. HNF abundance was estimated by epifluorescence microscopy. 2 ml sub-samples were stained with DAPI and filtered onto 2 µm pore size polycarbonate membranes. The sub-samples were counted with an epifluorescence microscope with UV excitation (1000X magnification); we switched to blue light excitation to distinguish heterotrophic from autotrophic nanoflagellates, the latter presenting red fluorescency under blue light. At least ten random transects, corresponding to an average of 100 cells, were counted from each sample. To estimate ciliate densities, a 45 ml subsample of each sample was stained with Bengal Rose and allowed to settle for 24 hr in settling chambers for inverted microscopy. The counts were made with the help of an inverted microscope (Wild M-40, 400X magnification) in random transects. At least ten transects, corresponding to about 50 cells, per sample were counted. For determination of rotifer abundances, a 1 l volume was filtered onto a 30 µm nylon mesh which was stored in 30 µm filtered lake water fixed with formalin to a final concentration of 5%. For enumeration, rotifers were washed off the mesh and the entire sample was enumerated in settling chambers using a binocular (80X).
Results
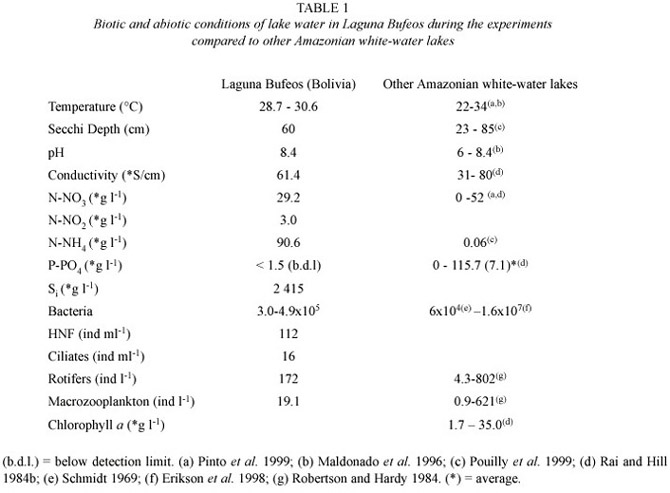
Water column conditions during the experiments are shown in Table 1. Sub-surface water temperature was quite constant with variations of less than 2º C. Transparency was low due to the amber pigmentation typical of waters rich in organic matter. Conductivity was very low and this is also reflected in the low concentration of nutrients. Phosphorus concentrations were below detection limit (1.5 * µg P l-1 ). Bacterial densities in Laguna Bufeos at the time of the experiments ranged from 3.0 to 4.9 x 105 cells ml-1 . HNF and ciliate densities in the lake were 112 and 16 ind ml-1, respectively, while rotifer and macrozooplankton densities were 172 and 19 ind l-1, respectively. The most important rotifer species observed were Brachionus caudatus and Polyarthra sp. The macrozooplankton community was dominated by Moina minuta and Diaphanosoma sp.
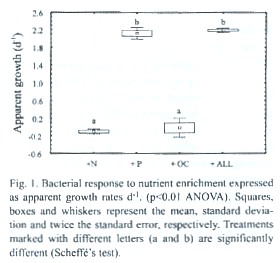
Bacterial growth rates in treatments receiving only nitrogen or only organic carbon were not significantly different during the incubation,whereas they increased strongly and significantly in treatments receiving phosphorus (ANOVA p < 0.01; Fig.1). Bacterial abundances in bottles enriched with phosphorus and in bottles enriched with all nutrients were approximately nine times higher than initial densities and the densities in the other treatments. Bacterial growth in the treatment receiving only phosphorus was not significantly different from the treatment receiving all nutrients (Scheffé s test).
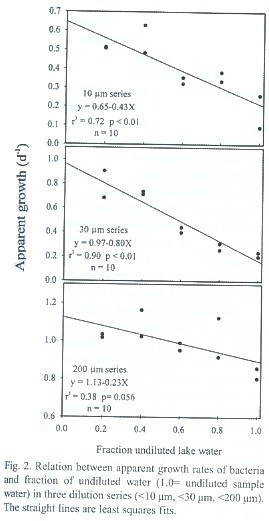
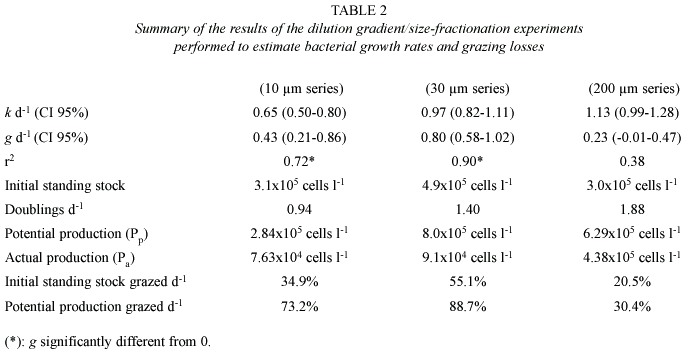
A negative relation between observed growth rates and dilution factor was observed in the three dilution series (< 10 µm, < 30 µm and < 200 µm; Fig.2) (Least squares regression). The calculated growth rate k in the absence of predation increased from 0.65 d-1 in the < 10 µm dilution series, over 0.97 d-1 in the < 30 µm dilution series up to 1. 13 d-1 in the < 200 µm dilution series (Fig.2, Table 2). The estimated grazing rate g was 0.43 d-1 in the 10 µm series and increased to 0.80 d-1 in the <30 µm treatment. In the <200 µm treatment, the slope of the regression line was marginally different from zero (p =0.056) and a grazing rate corresponding to 0.23 d-1 was calculated. Average densities of HNF did not differ significantly among the three size-fractions (ANOVA p<0.01) and varied between 61 and 104 cells ml-1. Initial numbers of ciliates increased from 4 cells ml-1 in the < 10 µm size fraction over 7 cells ml-1 in the < 30 µm fraction to 15 cells ml-1 in the <200 µm fraction. Rotifers were only observed in the < 200 µm size fraction. Initial rotifer abundance in the experiment was 172 individuals l-1.
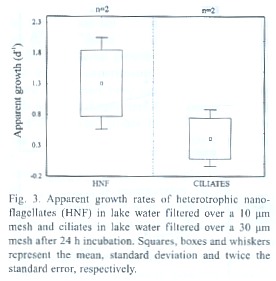
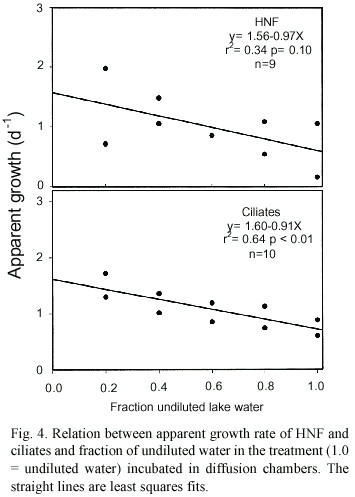
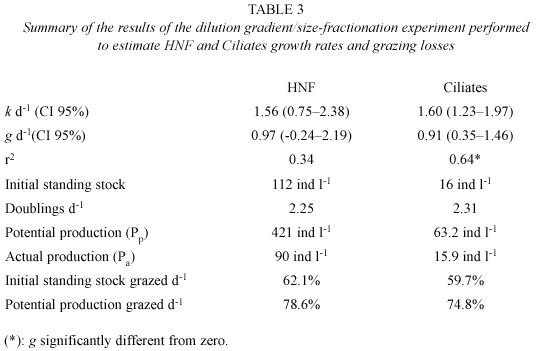
Growth rates k estimated in the size fractionation experiment were 1.31 d-1 for HNF and 0.42 d-1 for ciliates (Fig. 3). Grazing rates (of organisms between 10 and 200 µm in size) were 0.65 d-1 on HNF and 0.60 d-1 on ciliates. Growth rates k estimated by the dilution technique (Fig. 4, Table 3) were slightly higher when compared to the size fractionation estimates for HNF ( 1.56 d-1) and significantly higher for ciliates (1.60 d-1 ). The slope of the regression line was significantly different from zero in the case of ciliates only. Estimated grazing rates by organisms < 200 µm were 0.97 d-1 for grazing on HNF and 0.91 d-1 for grazing on ciliates.
Discussion
Nutrient concentrations as well as microbial and zooplankton densities were very low in Laguna Bufeos. Phosphorus concentration was particularly low (less than 1.5 µg l-1 ) but nitrogen was also present at relatively low concentration (but not limiting). Although floodplain lakes typically present low conductivity and nutrient content (Rai and Hill 1984b), nutrient concentrations in Laguna Bufeos were at the lower end of the range reported for this type of lakes (see Table 1). pH is normally neutral to slightly acid, but it varies greatly seasonally (5 -8.4 in floodplain lakes of River Ichilo), (Maldonado et al.1996). During the experiments, pH was at the higher end of this range.
Bacterial densities were also among the lowest compared to data published on other Amazonian lakes, which vary in a wide range (see Table 1). To our knowledge, no data have been published on HNF and ciliates densities in Neotropical floodplain lakes. Bacterial, HNF and ciliate densities in Laguna Bufeos were in the lower end of the range of values published for temperate lakes, where bacterial numbers range between 0.3 and 49 x 106 ml-1 (Letarte and Pinel Alloul 1991, Gasol et al. 1995). HNF densities vary between 190 and 30 000 cells ml-1 (Jürgens and Stolpe 1995, Pace et al. 1998) and ciliate densities range between 3 and 250 cells ml-1 (Sherr et al.1992,Gilbert and Jack 1993). Overall, abundances of components of the microbial food web as well as nutrient concentrations observed in Laguna Bufeos were comparable to abundances observed in oligotrophic temperate lakes.
The nutrient limitation assay clearly demonstrated phosphorus limitation of bacterial growth in Laguna Bufeos: when phosphorus was added, a strong stimulation of bacterial growth was observed. Early studies emphasised the importance of organic carbon in controlling bacterial populations, but more recently, bacterial growth limitation by phosphorus has been described in oligotrophic freshwater lakes (Sterner et al.1995) and marine environments (Elser et al. 1995). In humic lakes, which are somehow similar to Amazonian floodplain lakes due to their richness in organic matter, it has also been observed that growth of bacterioplankton can be nutrient limited rather than carbon limited. Since the C/N and C/P ratios of bacterial cells are usually substantially lower than that of the organic matter they use as a substrate, bacteria are dependent on external, inorganic sources of nutrients (Fenchel et al. 1998). The identity of the limiting nutrient will, hence, depend on the N/P ratio of the dissolved inorganic nutrient pool. Limitation of bacterial growth rates by the supply of phosphorus in this tropical lake is logical as the N/P ratios of planktonic bacterial cells (N/P =10; Fagerbakke et al. 1996) are lower than those of the available inorganic pool (N/P > 20).
The dilution experiment showed that HNF were the main grazers on bacterioplankton despite their low densities. Grazing by HNF accounted for 73.2% of the bacterial production (<10 µm series). The combined grazing activity of HNF and ciliates accounted for 88.7% of the production (<30 µm series). The fraction of the bacterial production that was grazed in the <200 µm series decreased to 30.4% (see table 2). We see two potential causes for this pattern: (1) a top-down effect of rotifers on HNF and ciliates reducing grazing rates in the <200 µm series;and (2) nutrients not constant with dilution, resulting from phosphorus recycling by rotifers grazing on HNF, ciliates and phytoplankton. However, our calculated grazing rates can not be interpreted straight-forward, phosphorus limitation has important implications for the dilution experiments set up to measure growth rates and grazing losses of bacteria since one of the assumptions of the dilution technique is that intrinsic bacterial growth rates are independent of dilution (Landry 1993). When phosphorus is limiting, one can expect intrinsic growth rates of bacteria to depend on dilution as more recycled phosphorus will be available in the least diluted treatments and more external phosphorus will be available per cell in the most diluted treatments. This problem can be avoided by adding nutrients in excess to the treatments. Since this was not done,the results of our experiments should be interpreted cautiously.
The fact that no nutrients were added to the dilution series should, however, allow us to make some considerations on nutrient cycling: bacterial growth rates estimated from the dilution experiments (k =0.65 –1.13 d-1 ) were lower than those found when phosphorus was added in the nutrient-limitation experiment (2.1-2.2 d-1 )but higher than when no phosphorus was added (~0), this suggests that in our experimental treatments, a phosphorus source stimulated bacterial growth. A plausible source of phosphorus is regeneration by microzooplankton (rotifers as well as ciliates or HNF) grazing on phytoplankton (Sterner et al.1995). Another possible source of phosphorus is that released from cell destruction during filtration (Landry 1993). This is, however, unlikely since no bacterial growth was observed in the nutrient-limitation experiment when water was filtered over 2 µm filter. Recycling by microzooplankton is therefore, the most likely source of phosphorus in the dilution series, and may explain why bacterial growth was most pronounced in the <200 µm series (k =1.13 d-1 ) and decreased from the <30 µm series (k =0.90 d-1 ) to the <10 µm series (k =0.65 d-1 ).
Our results agree with the general belief that a large proportion of the bacterioplankton production may be removed by predation; many studies have implicated HNF and occasionally ciliates as the major bacterivores (Sanders and Wickham 1993). Some authors conclude that these organisms are the main actors controlling bacterial densities in natural waters (Jiménez-Gómez et al.1994). Although this paper deals only with microzooplanktonic grazing, we should keep in mind that cladocerans are probably important grazers on bacteria even tough they represent 7% of the zooplanktonic community (in number of individuals). Further experimental work needs to be performed in order to determine the effect of macrozooplankton on the microbial community.
Concerning the dilution experiment in diffusion chambers, the results suggest that both HNF and ciliates feed on particles smaller than 2 µm (bacteria, possibly also autotrophic picoplankton) as, for both groups, apparent growth increased with dilution. The dominant predators on both groups are probably rotifers. The predation losses rate on HNF and ciliates were relatively high (g =0.97 d-1 for HNF and g =0.91 d-1 for ciliates). Even though growth rates were also high (1.5 d-1 for HNF and 1.6 d-1 for ciliates), the predation losses accounted for a major part of the HNF (78.6%) and ciliates (74.8 %) daily production (see Table 3), suggesting a strong top-down regulation of protistan populations.
Growth rates and grazing losses as measured by the size-fractionation and the dilution techniques were similar for HNF, meaning that cannibalism and predation by other organisms in the same size-fraction was not important. In the case of ciliates, the growth rate calculated by the fractionation technique was significantly lower than that calculated by the dilution method. This may be due to higher predation rates among members of the size-class 10-30 µm, but it is also likely that ciliates (which are more sensible to handling) have been damaged during filtration.
Due to their size, ciliates can be preyed upon by larger microzooplankton (mainly rotifers), macrozooplankton and even small fish, and they have the potential to be an important link from microbial to "traditional" food webs (Sanders and Wickham 1993). Unfortunately, the fate of ciliate production in freshwater plankton communities is poorly understood, even though it is assumed that much of it is ingested by cladocerans, copepods, and some rotifers (Gilbert and Jack 1993). Rotifers feed on nearly all the organisms that are part of the microbial loop. Like ciliates, rotifers may also constitute an important link between the microbial and "traditional" food webs.
In summary, we can conclude from our data that Laguna Bufeos is an oligotrophic system, with low microbial biomass and rich in organic matter as deducted from the dark colour of lake water causing low transparency (Secchi depth =60 cm). Decomposition of organic matter is mainly controlled by phosphorus, which is limiting bacterial growth. HNF and ciliates graze nearly all bacterial production and their densities are in turn controlled by rotifers. It seems apparent that protozoa and rotifers play a major role recycling nutrients and stimulate organic matter breakdown. The presence of grazers thus, both alleviates phosphorus limitation for bacteria (reducing bottom-up control) as well as strongly reduces bacterial densities (top-down control). The overall effect of microzooplankton on bacterial densities is positive, since bacterial growth under phosphorus limiting conditions and in the absence of grazers (Fig.1) was lower than in the presence of HNF, ciliates and rotifers (see apparent bacterial growth in undiluted water; Fig.2).
Acknowledgments
This research was supported by a grant in the framework of the Own Initiative "Limnological studies in the department of Cochabamba (PROLIMCO)", sponsored by the Flemish Inter-University Council (VLIR). We thank Frans Ollevier, Wim Vyverman and Hanne Degans for their logistic support.
Resumen
Que el bacterioplancton juegue básicamente un papel de reciclaje de nutrientes, o sea una fuente directa de nutrientes, depende de varios factores que afectan su producción y abundancia. En los lagos tropicales, la baja concentración de nutrientes es posiblemente el principal factor limitante del crecimiento bacteriano, y suele suponerse que la mayor pérdida poblacional de bacterias se debe a depredación por parte del microzooplancton. Estudiamos la regulación ascendente ("de abajo hacia arriba") y descendente ("de arriba hacia abajo") de abundancia bacteriana mediante seis experimentos in situ de limitación de nutrientes y de fraccionamiento de la dilución tamaño- gradiente. Bacterias, nanoflagelados heterotróficos (NHT), ciliados y rotíferos tienen densidades relativamente bajas. Las pérdidas por depredación de ciliados y de NHT explicaron la mayoría de la producción diaria, lo que sugiere que las poblaciones de protistas son reguladas por los rotíferos de forma descendente. El fósforo resultó ser un limitante fuerte del crecimiento bacteriano, pero no se detectó efecto del enriquecimiento con nitrógeno ni con "DOC". Los NHT fueron los principales depredadores de bacterias (g=0.43 d-1 ). El coeficiente de ramoneo aumentó al agregar los ciliados (g=0.80 d-1 ) pero disminuyó al agregar los rotíferos (g=0.23 d-1 ) probablemente debido a reciclaje de nutrientes o a control descendente de los NHT y ciliados por parte de los rotíferos.
Palabras clave: Bacterioplancton, microzooplancton, ramoneo, consumo, nutriente, reciclaje, lago tropical.
References
Anesio, A. M., P. C. Abreu & F. D. Esteves. 1997. Influence of the hydrological cycle on the bacterioplankton of an impacted clear water Amazonian lake. Microb. Ecol. 34:66-73. [ Links ]
Arndt, H. 1993. Rotifers as predators on components of the microbial web (bacteria, heterotrophic flagellates, ciliates)-a review. Hydrobiologia 255/256:231-246. [ Links ]
Currie, D. J. 1990. Large-scale variability and interactions among phytoplankton, bacterioplankton and phosphorus. Limnol. Oceanogr. 35:1437-1455. [ Links ]
Elser, J. J., L. B. Stabler & R. P. Hassett. 1995. Nutrient limitation of bacterial growth and rates of bacterivory in lakes and oceans: a comparative study. Aquat. Microb. Ecol. 9:105-110. [ Links ]
Erikson, R., K. Vammen, A. Zelaya & R. T. Bell. 1998. Distribution and dynamics of bacterioplankton production in a polymictic tropical lake (Lago Xolotlan, Nicaragua). Hydrobiologia 382:27-39. [ Links ]
Fagerbakke, K. M., M. Heldal & S. Norland. 1996. Content of carbon, nitrogen, oxygen, sulfur and phosphorus in native aquatic and cultured bacteria. Aquat. Microb. Ecol. 10:15-27. [ Links ]
Fenchel, T., G. M. King & T. H. Blackburn. 1998. Bacterial biogeochemistry:the ecophysiology of mineral cycling. Academic, San Diego, California. [ Links ]
Gasol, J., A. Simons & J. Kalff. 1995. Patterns in the top-down versus bottom-up regulation of heterotrophic nanoflagellates in temperate lakes. J. Plankton Res. 17:1879-1903. [ Links ]
Gifford, D. J. 1988. Impact of grazing by microzooplankton in the northwest arm of Halifax Harbour, Nova Scotia. Mar. Ecol. Prog. Ser. 47:249-258. [ Links ]
Gilbert, J. J. & J. D. Jack. 1993. Rotifers as predators on small ciliates.Hydrobiologia 255:247-253. [ Links ]
Jiménez-Gómez, F., V. Rodríguez & B. Bautista. 1994. Trophic interactions in the microbial food web at a coastal station in the Alboran Sea (Western Mediterranean) in winter. (II). Size selective flagellate feeding on bacteria and its implication on the microbial loop size-structure. Sci. Mar. 58:153-159. [ Links ]
Jürgens, K. & G. Stolpe. 1995. Seasonal dynamics of crustacean zooplankton, heterotrophic nanoflagellates and bacteria in a shallow, eutrophic lake. Freshwater Biol. 33:27-38. [ Links ]
Landry, M. R. & R. P. Hassett. 1982. Estimating the grazing impact of marine microzooplankton. Mar. Biol. 67: 283-288. [ Links ]
Landry, M. R. 1993. Estimating rates of growth and grazing mortality of phytoplankton by the dilution method. p. 715-722. In P. F. Kemp, B. F. Sherr, E. B. Sherr & J. J. Cole (eds.). Handbook of methods in aquatic microbial ecology. Lewis, Boca Ratón, Florida. [ Links ]
Letarte, Y. & B. Pinel-Alloul. 1991. Relationships between bacterioplankton production and limnological variables: Necessity of bacterial size considerations. Limnol Oceanogr. 36:1208-1216. [ Links ]
Maldonado, M., E. Goitia, F. Acosta, M. Cadima & D. Castellón. 1996. Caracterización limnológica de lagunas en la llanura aluvial del Río Ichilo, Cochabamba (Bolivia). Rev. Bol. Ecol. 1:29-37. [ Links ]
Neill, W. E. 1994. Spatial and temporal scaling and organization of limnetic communities.In P. S. Guiller, A. G. Hildrew & D. G. Raffaelli (eds.). Aquatic ecology: scale, patterns and process. Blackwell Scientific, Oxford, New York. [ Links ]
Pace, M. L., J. J.Cole & S. R. Carpenter. 1998. Trophic cascades and compensation:differential responses of microzooplankton in whole-lake experiments. Ecology 79:138-152. [ Links ]
Pomeroy, L. R. & W. J. Wiebe. 1988. Energetics of microbial food webs. Hydrobiologia 159:7-18. [ Links ]
Pouilly, M, M. Gutierrez & T. Yunoki. 1999. Funcionamiento ecológico de las lagunas de la zona de inundación del río Mamoré (Beni-Bolivia). Rev. Bol. Ecol.6:41-54. [ Links ]
Rai, H. & G. Hill. 1984a. Microbiology of Amazonian waters. p.413-442.In H.Sioli (ed.) Amazon, limnology and landscape ecology of a mighty tropical river and its basin. Dr. W. Junk, The Hague, The Netherlands. [ Links ]
Rai, H .& G. Hill. 1984b. Primary production in the Amazonian aquatic ecosystem.p.311-336.In H. Sioli (ed.). Amazon, limnology and landscape ecology of a mighty tropical river and its basin. Dr. W. Junk, The Hague, The Netherlands. [ Links ]
Robertson B. & E. Hardy. 1984. Zooplankton of Amazonian lakes and rivers. p. 337-352.In H. Sioli (ed.). Amazon, limnology and landscape ecology of a mighty tropical river and its basin. Dr. W. Junk, The Hague, The Netherlands. [ Links ]
Sanders R. W. ,K. G. Porter, S. J. Bennet & A. E. DeBiase. 1989. Seasonal patterns of bacterivory by flagellates, ciliates, rotifers and cladocerans in a freshwater planktonic community. Limnol. Oceanogr. 34:673-687. [ Links ]
Sanders, R. W., D. A. Caron & U. G. Berninger. 1992. Relationships between bacteria and heterotrophic nanoplankton in marine and freshwaters: an inter-ecosystem comparison. Mar. Ecol. Prog. Ser. 86:1-14. [ Links ]
Sanders R. W. & S. A. Wickham. 1993. Planktonic protozoa and metazoa: predation, food quality and population control. Mar. Microb. Food Webs. 7:197-223. [ Links ]
Schmidt G. W. 1969. Vertical distribution of bacteria and algae in a tropical lake. Int. Revue ges Hydrobiol. 54:791-797. [ Links ]
Sherr, B., E. Sherr & G. A. Paffenöfer. 1986. Phagotrophic protozoa as food for metazoans:a "missing" trophic link in marine pelagic food webs. Mar. Microb. Food Webs.1:61-80. [ Links ]
Sherr, E. & B. Sherr. 1988. Role of microbes in pelagic food webs: a revised concept. Limnol. Oceanogr. 33: 1225-1227. [ Links ]
Sherr, B., E. Sherr & J. McDaniel. 1992. Effect of protistan grazing on the frequency of dividing cells in bacterioplankton assembages. Applied Environ. Microbiol. 58:2381-2385. [ Links ]
Sterner, R. W., T. H. Chrzanowski, J. J. Elser & N. B. George. 1995. Sources of nitrogen and phosphorus supporting the growth of bacterio-and phytoplankton in an oligotrophic Canadian Shield lake. Limnol. Oceanogr. 40:242-249. [ Links ]
Zimmermann, H. 1996. Interactions between planktonic protozoans and metazoans after the spring bloom of phytoplankton in a Eutrophic Lake, the Belauer See, in the Bornhoveder Seenkette, North Germany. Acta Protozool. 35:215-221. [ Links ]

