










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.52 n.2 San José Jun. 2004
Nacional de Helmintos del Instituto de
Biología, UNAM, México
Jorge Falcón-Ordaz & Luis García-Prieto
Laboratorio de Helmintología, Instituto de Biología, UNAM, A.P. 70-153; C.P. 04510, México, D.F.; fax: 5 550 01 64; jfalcon@ibiologia.unam.mx; gprieto@ibiologia.unam.mx
Recibido 02-V-2002. Corregido 18-II-2004. Aceptado 25-II-2004.
Abstract
The present study analyses the taxonomic status of eleven species of trichostrongylins that parasitize rodents and lagomorphs deposited in the Colección Nacional de Helmintos, Instituto de Biología, UNAM, Mexico. This analysis is based on the morphology of the synlophe, characteristic that had not been studied for most of these nematode species and at present, it has a very important taxonomic value. As a result of this study, the identity of five species is ratified (Trichostrongylus calcaratus, Obeliscoides cuniculi, Carolinensis hue-huetlana, Stilestrongylus peromysci and Nippostrongylus brasiliensis), the transference suggested previously for two more (Vexillata convoluta and Vexillata vexillata) is confirmed, Trichostrongylus chiapensis is synonymized with Boehmiella willsoni, and finally Lamothiella romerolagi is redetermined as Teporingonema cerropela-doensis and Stilestrongylus atlatilpinensis as Stilestrongylus hidalguensis.
Key words: Nematoda, Trichostrongylina, Synlophe, Taxonomy, Rodents,
Lagomorphs, Mexico. Los nemátodos pertenecientes al suborden Trichostrongylina se caracterizan por la reducción de la cápsula bucal y su distintiva bolsa caudal; en su mayoría, exhiben ciclos de vida monoxenos, infectando al hospedero vía la ingestión de alimentos o a través de la piel. Son parásitos del estómago o intestino delgado de anfibios, reptiles, aves y mamíferos, alcanzando su máxima diversificación en estos últimos (Lichtenfels 1983). Hasta la fecha, en México se han descrito 29 especies pertenecientes a este suborden, 24 de las cuales se encuentran depositadas en la Colección Nacional de Helmintos del Instituto de Biología de la UNAM (CNHE) (Lamothe et al. 1997, Falcón-Ordaz et al. 2001). Como parte del proceso de actualización y depuración del acervo que se desarrolla en dicha colección, el presente trabajo analiza la situación taxonómica de once especies de Trichostrongylina parásitas de roedores y lagomorfos albergadas en la misma, incorporando el estudio del synlophe, estructura que no había sido descrita para la mayoría de este material y a la que se le confiere un peso taxonómico elevado (Durette-Desset y Chabaud 1981, Lichtenfels 1983, Durette-Desset 1983, 1985).
Materiales y métodos
Se analizaron ejemplares de once especies de tricostrongilinos parásitos de roedores y lagomorfos de México, depositadas en la CNHE (Cuadro 1). El material fue aclarado con lactofenol de Amman, efectuando preparaciones temporales para su estudio al microscópio óptico. La observación del synlophe se realizó de acuerdo con el método propuesto por Durette-Desset (1985): los ejemplares (1 macho y 1 hembra de cada especie en todos los casos, excepto en los que se señale un número diferente) se cortaron transversalmente en los niveles anterior, medio y posterior del cuerpo, mediante una navaja de bisturí, obteniendo cortes con un grosor de aproximadamente 0.110 mm.

Resultados
Apartir del análisis morfométrico efectuado al material de tricostrongilinos depositados en la CNHE, las especies estudiadas se dividieron en dos grupos; para las del primero se ratificó su identificación o su transferencia a otros géneros y para las del segundo proponemos modificaciones a su posición taxonómica. El primer grupo está compuesto por Obeliscoides cuniculi (Graybill, 1923) Graybill, 1924, Carolinensis huehuetlana Falcón & Sanabria, 1996, Nippostrongylus brasiliensis Travassos, 1914, Stilestrongylus peromysci Falcón & Sanabria, 1999, Trichostrongylus calcaratus Ransom, 1911, Vexillata convoluta (Caballero & Zerecero, 1943) Durette-Desset, 1971 y V. vexillata (Hall, 1916) Durette-Desset, 1970. Para este grupo de especies se adiciona información sobre el arreglo del synlophe a los niveles anterior y posterior del cuerpo: en ambos niveles, la disposición de las espinas del synlophe de O. cuniculi (Fig. 1 a-b) es semejante a la descrita por Durette-Desset (1978) para la región media. De manera similar, en este trabajo presentamos por primera vez la descripción de las regiones anterior y posterior del synlophe de C. huehuetlana (Fig. 1 g-i), ya que la región media fue estudiada previamente por Falcón y Sanabria (1996); para ambos sexos la región anterior presenta 11 espinas, con tres ventrales de mayor tamaño; en la región posterior, el número de espinas aumenta a 17 en machos y hembras. Los machos mantienen las tres espinas ventrales de mayor tamaño, mientras que en las hembras, a la altura del oviyector, exhiben un tamaño homogéneo.
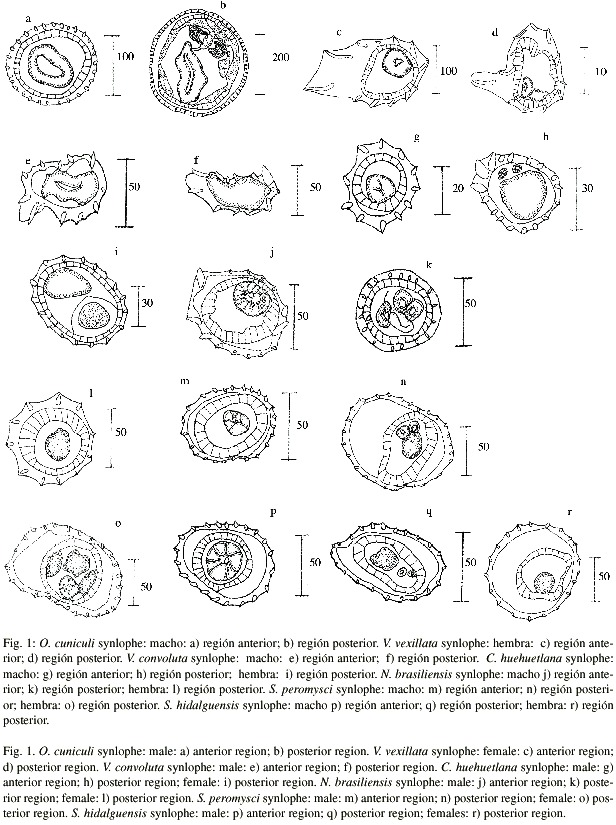
Para N. brasiliensis (Fig. 1 j-l) se confirmó el arreglo de las espinas del synlophe reportado por Lichtenfels (1974) en machos y hembras: en la región anterior este número va de diez a 14 en ambos sexos y de 15 a 18 y de 11 a 13 en la región posterior de machos y hembras, respectivamente. Por su parte, la región anterior del synlophe de los dos sexos de S. peromysci (Fig. 1 m-o) posee 24 espinas de tamaño diferente, las más pequeñas localizadas en el cuadrante ventral derecho de la región media, presentando el arreglo referido por Falcón y Sanabria (1999); a nivel de las espiculas se contaron 27 espinas (15 dorsales y 12 ventrales), siendo de menor tamaño las de los cuadrantes ventral y dorsal izquierdo; las hembras, a nivel del oviyector tienen 26 espinas de tamaño semejante.
Finalmente, la carencia de synlophe en T. calcaratus, determinó que la corroboración de su identidad se efectuara con base en la estructura de las espículas, que exhiben una curvatura apical, así como en el origen asimétrico del rayo dorsal y la ausencia de división en sus ramas, conjunto de rasgos diagnósticos de la especie de acuerdo con Ransom (1912).
El material de V. convoluta y V. vexillata (Fig. 1 c-f) depositado en la CNHE, fue asignado por Caballero y Zerecero (1943) y Caballero (1958), respectivamente, al género Longistriata Schulz, 1926; posteriormente, Durette-Desset (1972, 1978) transfirió ambas especies al género Vexillata (Travassos, 1937). Nuestro trabajo ratifica su transferencia a partir del estudio del synlophe en la región central del cuerpo. En el caso de V. convoluta se estudia por primera vez esta estructura en las regiones anterior y posterior del cuerpo, mismas que mantienen la disposición y número descritos para la porción central por Durette-Desset (1972, 1978); es decir, el arreglo de las 12 espinas sigue un eje frontal, con el carene dispuesto a lo largo del cuerpo.
Para el segundo grupo de especies que conforma este análisis, se proponen modificaciones de su posición taxonómica, las cuales se presentan a continuación:
Boehmiella wilsoni
Lucker, 1943
Sin. Trichostrongylus chiapensis
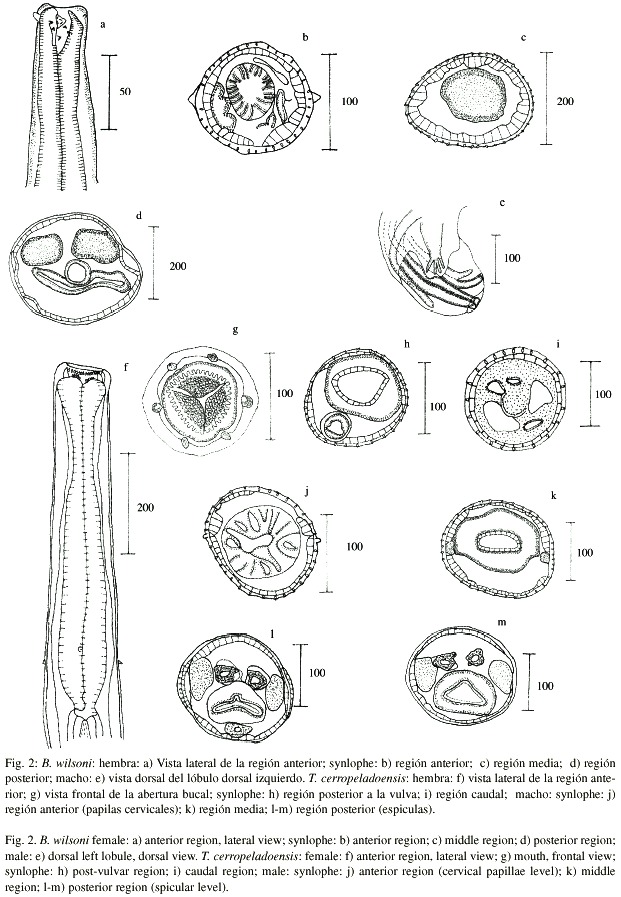
Synlophe: los cortes se realizaron en una hembra, observándose espinas pequeñas y perpendiculares al cuerpo, localizadas entre la región cervical y la región media. A nivel de las papilas cervicales se cuentan 34 espinas y al inicio del intestino 47, reduciéndose a 46 en la región preecuatorial; en la parte media solo aparecen 22 espinas hialinas. En el macho el synlophe se limita a la región media del cuerpo.
Comentarios: el material contenido en los lotes 2339 y 2340 de la CNHE fue identificado por Caballero (1958) como T. chiapensis; sin embargo, al reexaminar dicho material observamos que los rayos laterales de la bursa caudal están altamente quitinizados y que la boca en ambos sexos exhibe un diente dorsal, rasgos diagnósticos del género Boehmiella, de acuerdo con Gibbons y Khalil (1982). Adicionalmente, Trichostrongylus carece de synlophe, mientras que en Boehmiella si se presenta, pero restringido a la región anterior del cuerpo, tal y como se observa en el material de Caballero. Se asignaron a B. wilsoni por presentar las espiculas divididas en dos ramas, la abertura bucal provista de un diente dorsal, así como los rayos laterales quitinizados, rasgos similares a los descritos originalmente para la especie por Lucker (1943). Esta es la primera descripción del synlophe para la especie, además del primer reporte de la misma en México.
Teporingonema cerropeladoensis
Synlophe: se estudio en ocho ejemplares de ambos sexos; se inicia a 40-70 micrómetros del extremo anterior, exhibiendo espinas pequeñas y perpendiculares al cuerpo. En la región media de los dos sexos, las espinas laterales están agrupadas; en la región preespicular no se presentan espinas dorsales ni ventrales, únicamente laterales, que se desvanecen a nivel de la región media de las espiculas. El número de espinas en machos y hembras antes de las papilas cervicales va de 26 a 28, en la unión esófago-intestino de 28 a 43 y en la región media de 35 a 47. En los machos, el número disminuye hacia la región posterior, hasta llegar a 19 en la zona espicular; en las hembras también se observa una reducción, alcanzando un número mínimo de 24 posteriores al ano, con dos espinas hialinas laterales a nivel caudal.
Comentarios: El material de los lotes CNHE 2304 y 2305 fue establecido por González (1984) como especie tipo del género Lamothiella González, 1984 (L. romerolagi), parásito de Romerolagus diazi (Ferrari-Pérez, 1893), recolectado en Tlalpan, Ciudad de México. Por su parte, Harris (1985) con base en material proveniente de la misma especie de hospedero y localidad, erigió al género Teporingonema Harris, 1985, designando como especie tipo a T. cerropeladoensis, única descrita para el mismo hasta la fecha. Al reestudiar el material correspondiente al holotipo y paratipos de L. romerolagi, encontramos que exhiben las características diagnósticas del género Teporingonema: synlophe simple, cabeza truncada con borde muscular, boca con abertura hexagonal rodeada por cuatro papilas y dos anfidios, cavidad bucal con un anillo de dientes en forma de gancho, esófago claviforme, papilas cervicales prominentes, poro excretor que abre aproximadamente en el primer tercio de la longitud del cuerpo, espículas altamente esclerosadas, de igual tamaño, con una muesca en su región proximal y una punta compleja (Harris 1985). A partir de lo anterior, redeterminamos este material como T. cerropeladoensis debido a que el nombre específico asignado por González (1984) para el mismo no fue publicado.
Stilestrongylus hidalguensis
Synlophe: previo a este estudio, el arreglo y número de las espinas de esta especie se conocía exclusivamente a nivel medio del cuerpo (Falcón y Sanabria 1999): las espinas se disponen en un eje oblícuo, con las de mayor tamaño situadas en el cuadrante ventral derecho; su número oscila de 21 a 26 (24) en ambos sexos. En este estudio, se realizaron cortes del cuerpo de seis machos y dos hembras a dos niveles: en la región anterior, mantienen un arreglo y número similar al descrito anteriormente; en la región posterior, los machos exhiben de 19 a 22 (21) espinas y las hembras (a nivel del oviyector) presentan 20. Para ambos sexos éstas son pequeñas, de igual tamaño y perpendiculares a la pared del cuerpo.
Comentarios: Falcón (1993) describió dos especies del género Stilestrongylus Freitas, Lent & Almeida, 1937 procedentes de la misma localidad y hospedero: S. atlatilpinensis y S. hidalguensis, a las que diferenció con base en el arreglo de los rayos bursales del lóbulo izquierdo y en el número de espinas del synlophe. Sin embargo, el reestudio de este material (CNHE 2293 a 2295 y 2297), permitió corroborar que la disposición de los rayos bursales dos y tres es idéntica en ambas especies, separándose desde su inicio con una ligera curvatura y siendo relativamente más largo el rayo dos que el tres. El otro rasgo diferencial citado por Falcón (1993), con referencia al número de espinas del synlophe en la región media, no puede mantenerse como diagnóstico de S. atlatilpinensis, ya que la realización de nuevos cortes reveló que dicho número queda incluido en la variación que ahora establecemos para S. hidalguensis. Por estas razones, reasignamos el material determinado como S. atlatilpinensis por Falcón (1993) a la especie S. hidalguensis.
Discusión
La clasificación de los Trichostrongylina ha variado a lo largo del tiempo paralelamente a la adición de caracteres para su determinación taxonómica; no obstante, dos rasgos han sobresalido por su amplia aceptación: el primero, relacionado con el número, arreglo, tamaño y orientación de las espinas que conforman al synlophe y el segundo, referente a la disposición de los rayos bursales. Dichos rasgos constituyen los principales caracteres diagnósticos en los niveles subfamilia, género e inclusive especie.
En la mayoría de las especies, el conocimiento del synlophe se ha limitado a determinar su arreglo a la mitad del cuerpo (Lichtenfels 1983), aspecto que reduce su utilidad diagnóstica. El empleo generalizado que ha tenido este carácter, se debe principalmente a que ha demostrado ser estable, a pesar de que el parásito proceda de hospederos diferentes. Lo anterior fue observado en el presente trabajo, al estudiar los lotes de N. brasiliensis colectados en Rattus norvergicus Erxleben, 1777, cuyo synlophe, a todos los niveles, coincide con el descrito por Lichtenfels (1974) para la misma especie parasitando tres especies de hospederos (ratas, ratones y hámster). De la misma forma, para V. vexillata y V. convoluta constantemente se han descrito 12 espinas en la región media del cuerpo, aún cuando se han registrado parasitando hospederos de familias diferentes (Durette-Desset 1972, 1978; Falcón 1993 y el presente estudio). De la misma forma, autores como Lichtenfels (1977) y Lichtenfels y Pilitt (1983 a y b), han utilizado al synlophe como una herramienta para la separación de especies morfológicamente muy similares, pero que exhiben diferencias características a este nivel (por ejemplo en los géneros Nematodirella Yorke & Maplestone, 1926 y Nematodirus Ransom, 1907). Asimismo, el empleo del synlophe en la taxonomía del grupo, posibilita la identificación de las especies a partir de hembras en las cuales el synlophe sea idéntico al de los machos (Lichtenfels 1983); entre el material que ahora estudiamos, S. hidalguensis y N. brasiliensis exhiben un arreglo y número de espinas común para ambos sexos, lo que confirma el planteamiento anterior. Finalmente, caracteres del synlophe como número, tamaño y forma de las espinas, arreglo de las mismas a lo largo y alrededor del cuerpo y su ángulo de orientación, han mostrado su utilidad en los diferentes estudios filogenéticos realizados sobre el grupo (Durette-Desset y Justine 1991, Hoberg y Lichtenfels 1994, Durette-Desset et al. 1999, Pérez-Ponce de León et al. 2000).
La forma, arreglo y tamaño de los rayos bursales cuentan también con una amplia aceptación entre los especialistas del grupo (Durette-Desset y Chabaud 1977, 1981, Guerrero 1984). En el material depositado en la CNHE que analizamos, esta herramienta permite incluir a O. cuniculi y T. cerropeladoensis en la subfamilia Libyostrongylinae, ya que ambos exhiben el arreglo característico de la misma (rayo tres más largo que el dos, formando una "pinza"). A nivel genérico, las características de la bursa también juegan un papel importante; un ejemplo de ésto lo constituyen Libyostrongylus Lane, 1923 y Paralibyostrongylus Ortlepp, 1939, que p0resentan el rayo dorsal corto, mientras Obeliscoides Graybill, 1924 lo exhibe largo, aspecto que utiliza Durette-Desset (1983) para la separación de los géneros. De la misma forma, la utilidad de la bursa en la diferenciación de especies puede constatarse en nuestro material, al separar a V. vexillata de V. convoluta con base en la presencia de ramas anexas en el rayo dorsal de la primera, de las cuales carece la segunda; otro ejemplo lo constituye el arreglo de los rayos bursales de S. hidalguensis y S. peromysci, ya que en la primera, el lóbulo derecho exhibe un arreglo dos-dos-uno y el izquierdo dos-tres, mientras que en la segunda se presenta un mismo arreglo en ambos lóbulos (dos-dos-uno).
De los puntos referidos previamente se desprende que el número de especies representantes del suborden Trichostrongylina depositadas en la CNHE, se reduce de once a siete, manteniendo su determinación original únicamente cinco especies. Lo anterior ratifica la necesidad de realizar actualizaciones periódicas del acervo resguardado en las colecciones científicas, a la luz de metodologías y de caracteres taxonómicos importantes que no se hayan utilizado en sus descripciones originales, con el fin de preservar su valor sistemático y garantizar los beneficios educativos y de investigación que éstas deben proveer.
Agradecimientos
A Rafael Lamothe Argumedo, Curador de la CNHE, por las facilidades otorgadas para la realización de este estudio; a Gerardo Pérez Ponce de León, por la lectura crítica del manuscrito.
Resumen
El presente trabajo analiza la situación taxonómica de once especies de tricostrongilinos parásitos de roedores y lagomorfos depositadas en la Colección Nacional de Helmintos del Instituto de Biología, UNAM, a partir del estudio del synlophe, rasgo que no había sido estudiado para la mayor parte de este material y al que en la actualidad se le confiere un elevado peso taxonómico. Con base en dicho análisis, se ratifica la identidad de cinco de especies (Trichostrongylus calcaratus, Obeliscoides cuniculi, Carolinensis huehuetlana, Stilestrongylus peromysci y Nippostrongylus brasiliensis), se confirman las transferencias propuestas previamente para dos más (Vexillata convoluta y Vexillata vexillata), se sinonimiza a Trichostrongylus chiapensis con Boehmiella wilsoni, y se redetermina a Lamothiella romerolagi como Teporingonema cerropeladoensis y a Stilestrongylus atlatilpinensis como Stilestrongylus hidalguensis.
Referencias
Bravo, H.M. 1950. Estudio de nemátodos parásitos de los Lepóridos del Distrito Federal. Anales Inst. Biol. Univ. Nac. Autón. Méx. 21: 103-118. [ Links ]
Caballero, C.E. 1958. Estudios Helmintológicos de la región oncocercosa de México y de la República de Guatemala. Nematoda 10a parte. Anales Esc. Nac. Cienc. Biol. 9: 64-66. [ Links ]
Caballero, C.E. & M.C. Zerecero. 1943. Longistriata convoluta n. sp. (Nematoda: Trichostrongylidae) parásito del intestino de una "tuza" Crateogeomys merriami (Thomas). Anales Esc. Nac. Cienc. Biol. Méx. 14: 201-205. [ Links ]
Durette-Desset, M.C. 1972. Compléments morphologiques à létude de quelques Nématodes Héligmosomes, parasites de rongeurs américains. Ann. Parasitol. Hum. Comp. Paris. 47: 243-249. [ Links ]
Durette-Desset, M.C. 1978. Nouvelles donnèes morphologiques sur des nèmatodes trichostrongylides des collections du United States National Museum. Bull. Mus. Nat. Hist. Nat. Paris. 42: 501-508. [ Links ]
Durette-Desset, M.C. 1983. Keys to genera of the superfamily Trichostrongyloidea, p. 1-86. In R.C. Anderson & A.G. Chabaud (eds.). CIH key to the nematode parasites of vertebrates. Commonwealth Agricultural Bureaux, Farnham Royal, Bucks, Inglaterra. [ Links ]
Durette-Desset, M.C. 1985. Trichostrongyloids nematodes and their vertebrate hosts, pp. 239-306. In J.R. Baker & R. Muller (eds.). Advances in Parasitology. Academic, Londres. [ Links ]
Durette-Desset, M.C. & A.G. Chabaud. 1977. Essai de classification des Nématodes Trichostrongyloidea. Ann. Parasitol. Hum. Comp. Paris 52: 539-558. [ Links ]
Durette-Desset, M.C. & A.G. Chabaud. 1981. Nouvel essai de classification des Nématodes Trichostrongyloidea. Ann. Parasitol. Hum. Comp. Paris. 56: 539-558. [ Links ]
Durette-Desset, M.C., J.P. Hugot, P. Darlu & A.G. Chabaud. 1999. Acladistic analysis of the Trichostrongyloidea (Nematoda). Int. J. Parasitol. 29: 1065-1086. [ Links ]
Durette-Desset, M.C. & J.L. Justine. 1991. A cladistic analysis of the genera in the Subfamily Pudicinae (Nematoda, Trichostrongyloidea, Heligmonellidae). Int. J. Parasitol. 21: 579-587. [ Links ]
Falcón-Ordaz, J. 1993. Estudio taxonómico de los nemátodos parásitos de roedores de los géneros Peromyscus y Liomys, de Huehuetla y Atlatilpan, Estado de Hidalgo. Tesis de licenciatura, Universidad Nacional Autónoma de México, Tlalnepantla, Estado de México, México. 100 p. [ Links ]
Falcón-Oradaz, J., S.L. Gardner & G. Pérez-Ponce de León. 2001. Vexillata liomyos n. sp. (Nemata: Ornithostrongylidae) from Liomys pictus (Rodentia: Heteromyidae) from Mexico, with comments on the synlophe of Vexillata armandae. J. Parasitol. 87: 656-659. [ Links ]
Falcón-Ordaz, J. & M.A. Sanabria E. 1996. Especie nueva del género Carolinensis (Nemata: Heligmonellidae) de Peromyscus difficilis de Hidalgo, México. Anales Inst. Biol. Univ. Nac. Autón. Méx. 67: 67-75. [ Links ]
Falcón-Ordaz, J. & M.A. Sanabria E. 1999. Dos nuevas especies de Stilestrongylus (Nematoda: Heligmonellidae) parásitos de Peromyscus (Rodentia: Cricetidae) de México. Rev. Biol. Trop. 47: 929-937. [ Links ]
Gibbons, M.L. & L.F. Khalil. 1982. Akey for the identification of genera of the nematode family Trichostrongylidae Leiper, 1912. J. Helminthol. 56: 185-233. [ Links ]
González, O.M. 1984. Estudio taxonómico de algunos parásitos de roedores y lagomorfos mexicanos. Tesis de licenciatura, Universidad Nacional Autónoma de México, D.F., México. 56p. [ Links ]
Guerrero, R. 1984. Nematoda: Trichostrongyloidea parasites of venezuelan wild mammals III. The genus Vexillata Travassos, 1937. Ann. Parasitol. Hum. Comp. Paris. 59: 253-261. [ Links ]
Harris, E.A. 1985. Some helminths of the volcano rabbit Romerolagus diazi, including a description of the nematode Teporingonema cerropeladoensis gen. nov., sp. nov. (Trichostrongylidae: Libyostrongylinae). J. Nat. Hist. 19: 1239-1248. [ Links ]
Hierro, H.P. 1992. Helmintofauna de la "rata de alcantarilla" Rattus norvegicus Erxleben, 1777, de la Ciudad de Morelia, Michoacán, México. Tesis de licenciatura, Universidad Nicolaita de San Nicolás de Hidalgo, Morelia, Michoacán, México. 72 p. [ Links ]
Hoberg, E.P. & J.R. Lichtenfels. 1994. Phylogenetic systematic analysis of the Trichostrongylidae (Nematoda), with an initial assessment of coevolution and biogeography. J. Parasitol. 80: 976-996. [ Links ]
Lamothe-Argumedo, R., L. García-Prieto, D. Osorio-Sarabia & G. Pérez- Ponce de León. 1997. Catálogo de la Colección Nacional de Helmintos. UNAM-CONA-BIO, México, D.F. 211 p. [ Links ]
Lichtenfels, J.R. 1974. Number and distribution of ridges in the cuticle of Nippostrongylus brasiliensis (Travassos, 1914) (Nematoda: Heligmosomatoidea). J. Parasitol. 60 : 285-288. [ Links ]
Lichtenfels, J.R. 1977. Differences in cuticular ridges among Cooperia spp. of North American rumiants with an illustrated key to species. Proc. Helminthol. Soc. Wash. 44: 111-119. [ Links ]
Lichtenfels, J.R. 1983. The synlophe and species determination of Trichostrongyloidea, pp. 273-291. In A.R. Stone, H. M. Platt & L. F. Khail (eds.). Concepts in Nematode Systematics. Academic, Londres. [ Links ] Lichtenfels, J.R. & P.A. Pillit. 1983a. Cuticular ridge patterns of Nematodirus (Nematoda: Trichostrongyloidea) of North American rumiants, with a key to species. Syst. Parasitol. 5: 271-285. [ Links ]
Lichtenfels, J.R. & P.A. Pillit. 1983b. Cuticular ridge patterns of Nematodirella (Nematoda: Trichostrongyloidea) parasitic in domestic rumiants of North America, with a key to species. Proc. Helminthol. Soc. Wash. 50: 261-274. [ Links ]
Lucker, J.T. 1943. A new trichostrongylid nematode from the stomachs of American squirrels. J. Wash. Acad. Sc. 33: 75-79. [ Links ]
Ortega, M.L. 1976. Los parásitos más frecuentes en conejos domésticos en el Valle de México. Tesis de licenciatura, Universidad Nacional Autónoma de México, México, D.F., México. 100 p. [ Links ]
Pérez-Ponce de Léon, G., S.L. Gardner & J. Falcón. 2000. Phylogenetic relationships among species of Stilestrongylus Freitas, Lent and Almeida, 1937 (Trichostrongyloidea: Heligmonellidae: Nippostrongylinae), parasites of Myomorph rodents (Rodentia: Muridae) in the neotropics. J. Parasitol. 86: 1326-1335. [ Links ]
Ransom, B.H. 1912. Two new species of parasitic nematodes. Proc. U. S. Nat. Mus. 41: 363-369. [ Links ]

