










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.49 n.3-4 San José Dec. 2001
plantations in Northern Chiapas, Mexico
Lorena Soto-Pinto1, Yolanda Romero-Alvarado1, Javier Caballero-Nieto2, Gerardo Segura Warnholtz3
1 El Colegio de la Frontera Sur (ECOSUR). Apdo. Postal 63, San Cristóbal Chiapas, México, 29290, Tel. (967) 8-18-83 ext. 4202, Fax: (967) 8-23-22. Email: lsoto@sclc.ecosur.mx2 Jardín Botánico, Universidad Nacional Autónoma de México, Circuito Exterior, Ciudad Universitaria, México, D.F. 04510. Tel. 56-22-90-64. Email: jcnieto@servidor.unam.mx
3 Secretaría del Medio Ambiente, Recursos Naturales y Pesca, Av. Progreso No. 5, Viveros de Coyoacán, Col. El Carmen, Coyoacán, C.P. 04100, México D.F. Tel. y FAX: 55-54-39-52. Email: gsegura@semarnap.gob.mx
Abstract
Shade-grown coffee is an agricultural system that contains some forest-like characteristics. However, structure and diversity are poorly known in shade coffee systems. In 61 coffee-growers plots of Chiapas, Mexico, structural variables of shade vegetation and coffee yields were measured, recording species and their use. Coffee stands had five vegetation strata. Seventy seven woody species mostly used as wood were found (mean density 371.4 trees per hectare). Ninety percent were native species (40% of the local flora), the remaining were introduced species, mainly fruit trees/shrubs. Diametric distribution resembles that of a secondary forest. Principal Coordinates Analysis grouped plots in four classes by the presence of Inga, however the majority of plots are diverse. There was no difference in equitability among groups or coffee yields. Coffee yield was 835 g clean coffee per shrub, or ca. 1668 kg ha-1. There is a significant role of shade-grown coffee as diversity refuge for woody plants and presumably associated fauna, as well as an opportunity for shade-coffee growers to participate in the new biodiversity-friendly-coffee market.
Key words: Diversity, shade-grown coffee, Coffea arabica, vegetation structure, woody plants, agroecosystems.
Diversity loss concerns researchers and development agents who are studying the causes of diversity loss and trying to propose alternative solutions. In some countries researchers survey existing diversity, but many of these resources are accelerately being lost, it is estimated that around 150 species go extinct daily (Reid y Miller in Pimentel et al., 1992).
The loss of diversity has occurred mainly due to agricultural, industrial, and urban expansion that has penetrated the agricultural frontier, in addition to other factors such as use of pesticides, introduction of exotic species and forest fragmentation (Pimentel et al. 1992, Brown and Lugo 1994, Lugo 1995).
Some agricultural partially resemble to the natural environment; shaded coffee plantations, maintain soil, climatic and biological conditions similar to surrounding natural forests (Jiménez-Avila and Martínez 1979, Perfecto et al. 1996, Moguel and Toledo 1999, Zúñiga and Calvo unpublished). However, in many countries, coffee growers have used single genus combinations for shade. In other cases, they have cultivated only one species, and in the most extreme cases, they have used unshaded coffee. As a result, this simplification process has caused diversity loss and other problems associated with it, such as erosion, pest attacks, diseases and loss of resource availability in the coffee stand (especially for subsistence producers) (Jiménez-Avila and Martínez 1979, Moguel and Toledo 1999).
Recent studies have shown the significance of managed forests for bio-diversity conservation (Hansen et al. 1991), diverse shade-grown coffee plantations can be considered managed forests. It has been reported that shade-grown coffee is very important as habitat for birds (Aguilar-Ortiz 1982, Greenberg et al. 1997, 1997a), mammals (Estrada et al. 1993, Gallina et al. 1996), reptiles (Seib, cited by Perfecto et al.1996), insects (Torres 1984, Perfecto et al. 1997), arachnids (Ibarra-Núñez 1990), woody plants (Purata and Meave 1993) and orchids (Nir 1988), which find refuge within coffee stands. Additionally, shade-grown coffee plantations can be considered as corridors for fauna and flora, since they are part of the scarcely vegetated areas within fragmented zones (Moguel and Toledo 1999). However, there are few studies that describe structural characteristics and woody plant species diversity in coffee stands that make possible to define criteria for shade-grown coffee plantations. Such studies have acquired importance due to the recent developing bio-diversity friendly coffee markets in the United States and Europe. This market gives producers the opportunity to increase their income by selling "ecological friendly coffee"; additionally, it benefits consumers who prefer specialty coffee, and helps to maintain environmental health.
The present study describes and analyzes coffee stand structure and woody plant diversity in shade-grown coffee systems in Northern Chiapas, Mexico.
Materials and methods
Study area and selection of sampling sites: this study was carried out in two areas of Northern Chiapas. One in the municipality of Chilon and the other in the municipality of Jitotol. Both zones are located in the coffee belt of Northern Chiapas, Mexico, within the subtropical zone characterized by a warm and humid climate (A)C(m) (García 1988). The area presents a gradient between 800 to 1200m above sea level, having an annual rainfall of 1200 to 1500 mm in the higher altitudes; and 2000 mm at the lower altitudes. Average annual temperature is 22° C. Soils are recent, thin, and stony. Typical vegetation in the highest areas is the montane rain forest (bosque mesófilo de montaña), and in the middle and lowest zones semi-evergreen seasonal forest (bosque tropical subperennifolio (Rzedoswki 1978, Breedlove 1981).
The population of the Jitotol area belongs to the the Tzotzil ethnic group and in Chilon they are Tzeltales. Both of them are Mayan descendants whose main activity is agricultural production of corn, beans, squash and coffee along with some cattle raising.
A characterization of main environmental and technical conditions in which coffee is grown was carried out. A questionnaire was given to 102 producers from Chilon and Jitotol to identify the main variants in regional coffee systems. Sixty-one sampling sites were chosen, controlling for coffee plant age, variety, and type of shade and management (organic vs. chemical). In each of the 61 plots a permanent sampling area of 10m x 10m was selected, where measurements of the studied variables were conducted. The square size was defined in order to relate community structure and canopy cover (Anderson 1966) based on the assumption that resulting image from hemispherical photographs gives an almost 180 degree view in all directions, with the zenith at the center and the horizon at the edges of the photograph (Roxburgh and Kelly 1995).
Measurements of coffee stand structure and diversity: Coffee density, slope and plot aspect were measured. In each plot a forest inventory with the following variables was carried out: number of individuals of each shade species, separated into nine diametric classes (1-10cm, 10.1-20cm, 20.1-30cm, 30.1-40cm, 40.1-50cm, 50.1-60cm, 60.1-70cm, 70.1-80 cm, and > 80 cm in breast height diameter (breast height being 1.3m); life forms (tree, shrub, palm and tall herbs). The number of strata in coffee stand as well as height, use of the species and qualitative presence of woody plant seedlings were recorded. With the above-mentioned information the following variables were estimated: density, frequency, abundance, species richness, diversity index and importance value (Mueller-Dumbois 1974, Whittaker 1975). All trees and shrubs in the plots were mapped in order to define number of strata. Light density class, crown form, and number of dead trees were also recorded (Hutchinson 1988). Illumination classes considered by the position of tree crown with respect to the canopy were the following: 1) emergent, 2) full overhead light, 3) some overhead light, 4) mainly oblique light, and 5) indirect light. Crown form classes included: 1) complete circle, 2) irregular circle, 3) half circle, 4) less than half circle, 5) few branches, 6) mainly sprouts, 7) alive without crown. Voucher specimens were collected for identification and deposited in the ECOSURs herbarium (El Colegio de la Frontera Sur) and at the National Herbarium of Universidad Nacional Autónoma de México (MEXU).
Hemispherical photos were taken in each plot (for methodology see Soto-Pinto, 2000). Hemiphot (a computer program) was used to calculate canopy cover and photon flux density below the canopy (Steege 1996).
A Principal Coordinates Analysis (Everitt and Dunn 1991) based on the Jaccards Similarity Index (Sneath and Sokal 1973) was carried out in order to identify groups of plots by floristic composition though the computational program NTSYS. Unidentified species were not included in the analysis. For each group, plot equitability was estimated through the following formula:
E= BA/R
Where:
E= plot equitability
BA= average basal area
R= species richness in 100m2
Results
Coffee producers are indigenous people, which possess in general less than 5 hectares in total; about one fifth of this land is devoted to coffee production. Shade-grown coffee is managed as a low-input system. In Jitotol there are organically-grown coffee, chemical-grown-coffee and natural-grown coffee (no inputs), while in Chilon the natural-coffee prevails. Producers do not control any pest or disease besides the berry borer, what is controlled by cultural methods. Weeds are handling removed twice a year. Cultural practices, except for harvesting are based upon family labor.
Coffee stands present a mixture of varieties Bourbon, Caturra, Mundonovo and Typica in an average density of 1500 shrubs per hectare. Shade trees are densely non-uniformly distributed in the plots, since producers tolerate seedlings growing naturally in the coffee plantation.
Shade vegetation structure in coffee stands showed a complex vertical profile. There were five strata of shade vegetation: one herbaceous stratum, two shrubby strata and two tree strata. The average profile showed 26.4% of emergent trees in the canopy; 21.5% of individuals with full overhead light, 18.9% of individuals with some overhead or oblique light; and 34.2% with trees or shrubs with indirect light (not including coffee shrubs). Different species were represented in each stratum. The species: Inga pavoniana, I. punctata, Belotia mexicana, Heliocarpus appendiculatus and I. sapindioides were mainly emergent trees with full overhead light, while Heliocarpus appendiculatus, Citrus sinensis, Musa sapientum and Croton draco mainly had some overhead or oblique light. Additionally, the species Oecopetalum mexicanum, Piper auritum, Astrocharium mexicanum, Chamaedorea cataractarum, Musa sapientum and Neurolaena lobata represented the stratum receiving indirect light. Frequency of species by illumination classes is presented in Table 1. Canopy cover ranged between 23 and 80%.
Jitotol, Chiapas, México.
| Species | Illumination Classes (%) | ||||
| Emergent trees | Full overhead light | Some overhead light | Mainly oblique light | Indirect light | |
| Inga pavoniana | 24.7 | 27.9 | 10.0 | 2.9 | 0 |
| Inga punctata | 9.9 | 13.2 | 3.3 | 2.9 | 1.5 |
| Belotia mexicana | 9.9 | 10.3 | 3.3 | 0 | 2.9 |
| Heliocarpus appendiculatus | 6.2 | 2.9 | 11.7 | 8.8 | 1.5 |
| Inga sapindioides | 6.2 | 2.9 | 3.3 | 2.9 | 1.5 |
| Citrus sinensis | 0 | 0 | 8.3 | 2.9 | 4.4 |
| Other species | 43.1 | 57.2 | 60.1 | 79.6 | 88.2 |
The average tree density was 371.4 trees ha-1, varying between 100 and 800 trees ha-1. Of all shade trees or shrubs, 76% was woody trees. Average species richness was 3.2 species per 100m2, varying between one and eight species per 100m2.
Tree height varies between 1 and 29m, diameters vary between 1 and 109 cm. Most of shade components were in the range of < 20cm diameter. Diametric distribution resembles that of the secondary forest. Inga contributes significantly to diameters between 30 and 80 cm (Fig.1).
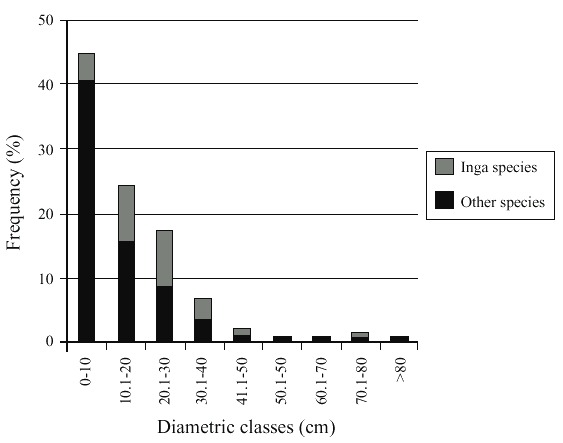
Tree crowns presented different forms: 49% were a complete circle; 14% an irregular circle; 21% a half circle; and the remaining 16% showed less than a half circle, few branches or mainly sprouts. There were no alive without-crown trees or shrubs registered.
Seventy-seven woody species were registered in the sampling plots (Table 2). Of the total number of species, 90% were native ones, the remaining were introduced species, mainly fruit trees. Most shade species of coffee stands belong to tropical forests. The remaining belongs to montane forests or are present in both vegetation types.
of shade-grown-coffee plantations in Northern Chiapas, Mexico.
| Family | Species | Local Name | Life Form | Origin | Vegetation type | Relative frequency (%) | |
| Anacardiaceae | Mangifera indica L. | Mango | T | I | --- | 1.6 | |
| Araliaceae | Dendropanax arboreus (L.) Dacne & Planchon | Ya can chamel | S | N | TF | 1.6 | |
| Arecaceae | Astrocharium mexicanum Liebm.* | Chapay | P | N | TF | 3.2 | |
| Chamaedorea cataractarum Liebm.* | Chib | P | N | TF | 4.8 | ||
| Chamaedorea tepejilote Liebm.* | Joma | P | N | TF | 3.2 | ||
| Desmoncus schippii Burr.* | Baas | P | N | TF | 1.6 | ||
| Asteraceae | Clibadium arboreum Donn. Sm. | Jaalte | T | N | TF | 1.6 | |
| Eupatorium chiapensis Rob. | Yashajalte | S | N | TF | 3.2 | ||
| Montanoa sp.* | Malacate | T | N | CF | 1.6 | ||
| Neurolaena lobata (L.) R. Br. | Pomte | S | N | BT | 12.9 | ||
| Tithonia rotundifolia (Miller) Blake* | Sun | S | N | TF | 1.6 | ||
| Vernonia deppeana Less.* | Sitit | S | N | TF | 1.6 | ||
| Vernonia patens H.B.K.* | Sitit | S | N | TF | 1.6 | ||
| Bignoniaceae | Amphitecna macrophylla (Seem.) Miers. | Mistel | T | N | TF | 1.6 | |
| Burseraceae | Bursera simaruba (L.) S. | Chacaj | T | N | TF | 3.2 | |
| Caricaceae | Carica pennata Heilb. | Papaya | T | N | TF | 1.6 | |
| Elaeocarpaceae | Muntingia calabura * | Capulin | T | N | CF | 3.2 | |
| Euphorbiaceae | Bernardia aff. interrupta (Schel.) Muell-Arg. | Sac juluchay | T | N | TF | 3.2 | |
| Croton billbergianus Mull Arg. | Tuminte | T | N | TF | 1.6 | ||
| Croton draco Schlecht.* | Chich bat | T | N | TF | 11.3 | ||
| Sapium sp. | Juun | S | N | TF | 1.6 | ||
| Fabaceae | Calliandra sp.* | Shashim | S | N | CF | 1.6 | |
| Erythrina sp.* | Mote | T | N | TF | 3.2 | ||
| Inga pavoniana Donn.* | Coquilte | T | N | TF | 41.9 | ||
| Inga punctata Willd.* | Tzelel | T | N | TF | 24.2 | ||
| Inga sapindioides Willd. | Cok | T | N | CF | 12.9 | ||
| Lonchocarpus sp.* | Shinte | S | N | TF | 4.8 | ||
| Senna multijuga (L. C. Rich.) I. & B. var doylei | Xaxibte | T | N | TF | 1.6 | ||
| Senna papilosa (B. & R.) I. & B.* | Cantelal tzi | T | N | TF | 1.6 | ||
| Fagaceae | Quercus sp. | Tzizte | T | N | CF | 1.6 | |
| Hamamelidaceae | Liquidambar styraciflua L. | Tzoste | T | N | TF | 1.6 | |
| Icacinaceae | Oecopetalum mexicanum Gr. & Th. | Cacaté | T | N | TF | 3.2 | |
| Lauraceae | Nectandra globosa (Aublet) Mez.* | Onte | T | N | TF | 1.6 | |
| Persea schiedeana Nees* | Chinino | T | N | TF | 3.2 | ||
| Marantaceae | Calathea macrochlamys Woodson & Standl.* | Pimil | H | N | TF | 6.5 | |
| Melastomataceae | Conostegia xalapensis (Bonpl.) D.Don | Pem chitom | S | N | CF | 1.6 | |
| Miconia aff. ibaguensis (Bonpl.)Triana* | Chactajmut | T | N | TF | 1.6 | ||
| Meliaceae | Cedrela mexicana Roe | Cedro | T | N | TF | 3.2 | |
| Moraceae | Castilla elastica Cerv. | Hule | T | N | TF | 3.2 | |
| Cecropia obtusifolia Bert* | Guarón | T | N | BT | 3.2 | ||
| Musaceae | Musa sapientum L. | Plátano roatan | T | I | --- | 9.7 | |
| Myricaceae | Myrica cerifera L. | Atsamte | T | N | BT | 1.6 | |
| Myrsinaceae | Myrsine myricoides (Schlecht.) Lundell | Atsamte | T | N | BT | 3.2 | |
| Myrtaceae | Casearia corymbosa Kunth | Tanchit | T | N | TF | 1.6 | |
| Sysingium jambos L.* | Pomarrosa | T | I | --- | 6.5 | ||
| Psidium guajava L.* | Guayaba | T | N | TF | 4.8 | ||
| Piperaceae | Piper auritum Kunth* | Momun | H | N | TF | 1.6 | |
| Piper patulum Bertol. | Momun chitom | S | N | CF | 1.6 | ||
| Rhamnaceae | Rhamnus capraefolia Shclecht. | Kanolte | T | N | TF | 1.6 | |
| Rubiaceae | Blepharidium mexicanum Standl. | Xacaxte | S | N | TF | 1.6 | |
| Rutaceae | Citrus aurantifolia Osb. | Limón | S | I | --- | 1.6 | |
| Citrus reticulata Lour. | Mandarina | S | I | --- | 1.6 | ||
| Citrus sinensis Osb. | Naranja | S | I | --- | 8.1 | ||
| Zanthoxylum aff. microcarpum Griseb* | Pajulte | T | N | TF | 3.2 | ||
| Zanthoxylum aff. kellermanii P. Wilson* | Ichilte | T | N | BT | 1.6 | ||
| Sapindaceae | Cupania dentata D.C. | Tojposte | T | N | TF | 1.6 | |
| Sapotaceae | Pouteria zapota Merr. | Zapote | T | N | TF | 1.6 | |
| Chrysophyllum caimito L. | Caimito | P | N | CF | 1.6 | ||
| Chrysophyllum mexicanum (Brand) Standl.* | Chiit | T | N | BT | 9.7 | ||
| Solanaceae | Solanum aphyodendron Knapp* | Ashinte | S | N | TF | 1.6 | |
| Sterculiaceae | Theobroma cacao L. | Cacao | S | N | TF | 4.8 | |
| Tiliaceae | Belotia mexicana Shum.* | Ikbat | T | N | TF | 12.9 | |
| Heliocarpus appendiculatus Turcz.* | Saquil Bat | T | N | BT | 16.1 | ||
| Heliocarpus donnell-smithii Rose* | Jono ha | T | N | TF | 3.2 | ||
| Heliocarpus mexicanus (Turcz) Sprague* | Sajal Bat | T | N | TF | 1.6 | ||
| Heliocarpus reticulatus Nash* | Xochbat | T | N | TF | 1.6 | ||
| Urticaceae | Myriocarpa longipes Liebm. | Bacte | T | N | CF | 3.2 | |
| Verbenaceae | Lippia myriocephala Schlech. & Cham.* | Sac Mumus | T | N | TF | 1.6 | |
| Violaceae | Orthion subssesile (Standl.) Steyerm. & Stadl. | Weel | T | N | TF | 1.6 | |
| Non identified | Jititul | T | - | TF | 1.6 | ||
| Non identified | Netamte | T | - | CF | 4.8 | ||
| Non identified | Pisisnich | S | - | CF | 1.6 | ||
| Non identified | Suchalanal | S | - | CF | 3.2 | ||
| Non identified | Tzajal tzit | S | - | CF | 1.6 | ||
| Non identified | Tzerey | T | - | CF | 4.8 | ||
| Non identified | Ujchum | S | - | TF | 1.6 | ||
| Non identified | Yaxte | S | - | CF | 1.6 | ||
| Non identified | Zon | S | - | CF | 1.6 | ||
Origin key: N= native, I= introduced; vegetation type: TF=tropical forest, CF=cloud forest, BT=both types; life form: H=herb, S=shrub, T= tree
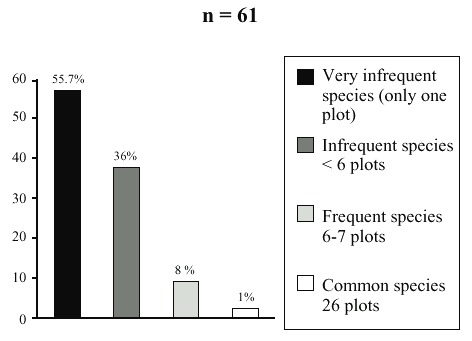
Of the total of shade species, 61.5% are trees, 29.5% shrubs, 6.4% palms, and 2.6% tall herbs. Thirty-three families and 56 genera were identified. The most numerous families are Fabaceae (legumes), followed by the Tiliaceae, Asteraceae, Arecaceae, Euphorbiaceae and Rutaceae. Most of the species of these families (55%) were infrequent, occurred in only one plot. Other group of species (35.9%) is more frequent (10% of the total the plots). A smaller group (7.7%) is the most frequent species (10-12% of the total the plots) (Fig. 2). The most frequent-species were: I. pavoniana, I. punctata, Heliocarpus appendiculatus, Neurolaena lobata, Belotia mexicana, Inga sapindioides, and Croton draco. Although coffee shade is diverse in woody species, 38% of the plots presented high importance values for any of the Inga species. The rest of the plots were dominated by other 26 species.
In most plots (83.3%), seedlings of woody species growing from the natural vegetation were registered. These seedlings are tolerated by coffee growers due to their function as useful or protection (shade) plants. Seedlings of 34 useful species were registered. Most salients were Nectandra globosa, Chrysophyllum mexicanum, Chamaedorea cataractarum, and Inga pavoniana (Table 2).
Ninety percent of the species present in the coffee stands are of some use to the producers. One hundred and twenty-three products or services were registered, most of them being represented by firewood (27.6%), followed by food (23.6%) and construction materials (9.8%). The remaining are represented by plants recognized as good shade for coffee (16%), poles used for living fences (5%), prime material for handcrafts and labor instruments (3.3%), home remedies (3.3%), fibers (1%), gums (1%), fodder (1%) and other domestic uses (8.4%). Of food species, 29 different products were registered: fruits, stems and edible flowers, with fruits the most frequent. Other organisms observed in the coffee stands are mushrooms, and plant species of the families Orchidaceae, Bromeliaceae, Cicadaceae and Araceae. Three percent of the total individuals are dead trees or shrubs.
Average yield in sampling plots is 835 g of clean coffee per bush or approximately 1668 kg per ha-1.
The Principal Coordinate Analyses shows four groups of plots: the first group includes plots (57.6% of the total of plots) with diverse composition of species; the following group comprises plots (24.5% of the total of the plots) with diverse species and I. pavoniana; the third group contain plots (9.8% of the total the plots) with I. punctata and other diverse species; and finally, the last group comprises plots (8.1% of the plots) including I. punctata, I. pavoniana and other diverse species (Fig. 3). Equitability and yields were not significantly different among groups (p < 0.05).
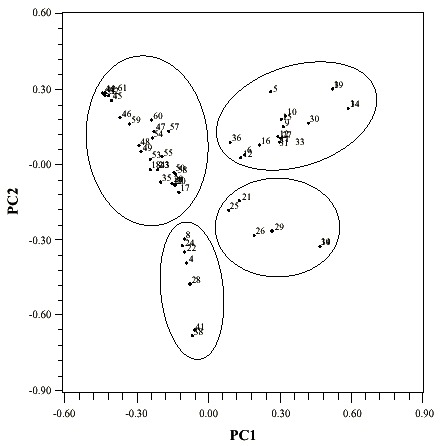
resulting from the Principal Coordinate Analyses based on a Jaccard index
similarity matrix.
Discussion
Most of the coffee shade species are native species and correspond to 40% of the species reported for the coffee belt in northern Chiapas (Miranda1953, Breedlove 1986, Del Amo et al. 1992). This highlights the salient role of shade-grown coffee system in the conservation of woody flora, as has been pointed out by Purata and Meave (1993). Shade-grown coffee systems play an important role for conservation, specially for species with narrowed distribution, as in the case of species restricted to montane forests or tropical forest as the majority of species in this study are. Even when the results showed four groups of coffee stands in relation to their woody species composition, there was no difference in equitability among groups. Each coffee stand is practically unique in plant composition, except for the presence of Inga species, whose dominance was corroborated in 38% of plots. However, the majority of plots were diverse.
The dominance of Inga species could be due to the intervention of INMECAFE (Instituto Mexicano del Café), a government institution with wide impact on coffee activities in Mexico. This institute promoted a technological package that included the use of improved coffee varieties, use of inorganic fertilizers, pesticides and the use of species of Inga as shade trees. Actually, producers of several zones in Mexico still maintain the idea that Inga is the best choice for shade, and although the INMECAFEs dissemination programs have finished, producers still are interested in shifting diversified shade by shade dominated by one or two species of Inga. Producers tolerate seedlings of Inga species, one of the most notable species in secondary regeneration, and even, in some places they grow seeds for sowing. The fact that plots were grouped in four classes, depending on the presence of Inga, responds to this technological innovation occurred in last three decades. However, Romero-Alvarado (pers. comm. 2000) and Peeters (pers. comm. 2000) could not find significant differences in yields comparing Inga-dominated plantations versus rustic plantations. This mosaic diversity acquires importance for conservation of associated flora and fauna (Aguilar-Ortiz 1982, Torres 1984, Brash 1987, Nir 1988, Pimentel et al. 1992, Purata and Meave 1993, Perfecto and Vandermeer 1994, Perfecto and Snelling 1995, Greenberg et al. 1997a, b, and others summarized by Perfecto et al. (1996). The heterogeneity of vegetation can play an important role in habitat selection for associated fauna, e.g. bird species (Greenberg et al. 1997). This heterogeneity is maintained by the following factors: the different crown forms, mainly complete and irregular circles, the presence of several vegetation strata, an irregular (non-systematic) disposition of shade trees, a high number of different taxa, the presence of dead trees, epiphytes, mushrooms and herbs. This species mosaic probably contributes to biological diversity. Unfortunately, the unavailability of studies in natural forest in this life zone do not permit to compare ecological and structural features between these and coffee plantations.
Although we did not quantify secondary regeneration, the presence of seedlings of many of the woody species reported was found in the plots. Consequently, since growing of diverse woody native species is promoted through the tolerance of seedlings, the role of producer in bio-diversity conservation is significant.
The majority of species have some useful value. Most notables are those that offer products that can not be substituted and are of primary necessity such as firewood, food and materials for construction. Other uses are less generalized, such as the use of Heliocarpus donnell-smithii (bark inhabits the larva of an edible butterfly), Belotia mexicana (bark is used to make rope) and Calathea macroclamys (leaves used to wrap "tamales", a traditional Mexican dish made of corn and wrapped in natural leaves). In this sense, it is very important to note the important role of producers knowledge in adding ecological and socioeconomic benefits to local and global scopes.
Although producers maintain a high tree density in their plots which provides the aforementioned products or services, this density does not avoid the maintenance of coffee yields even above the Mexican average. According to Soto et al. (2000), coffee yield behaves in a quadratic form in relation to the percentage of shade cover, with the highest yields nearer to 50% cover, with an average of 463 shade trees. In addition, Romero-Alvarado (pers. comm., 2000) also reported that shade tree density had no effect on crop yields.
These results lead us to think that coffee growers working on shade-grown coffee plantations have the opportunity to have a place in the bio-diversity friendly coffee production and in the specialty coffee market without a reduction in yields. Some estimations suggest that shade-grown coffee represents roughly one to two percent of the specialty coffee market, with total sales estimated at US $ to $60 million per year (Commission for the Environmental Cooperation Web Page, 2000. //www.cec.org). At the same time, shade-grown coffee can contribute to the preservation of natural resources and wildlife habitats in Mexico.
Acknowledgements
We thank coffee growers form the following localities: Alan Kantajal, Segundo Cololteel and Muquenal from Chilón Municipality, as well as Francisco I. Madero, from the Jitotol Municipality for their interest and participation in this study. We thank Manuel Martínez Gómez for field support, and Mario Ishiki and Juan Castillo Hernández for species identification. Field work was supported by Sistema de Investigación Benito Juárez (SIBEJ 98-01-021), Consejo Nacional para el Conocimiento y Uso de la Biodiversidad-MacArthur Foundation (CONABIO M018) ) and El Colegio de la Frontera Sur (ECOSUR). Kristen Nelson (University of Minnesota) and three anonymous reviewers provided valuable comments on a draft of this manuscript.
Resumen
El café bajo sombra es un sistema agrícola que contiene algunas características de los bosques. Sin embargo, las características estructurales y de diversidad de la sombra del café son poco conocidas. En 61 parcelas de productores del norte de Chiapas, Mexico, se midieron variables estructurales de la vegetación de sombra y los rendimientos de café, registrando las especies y sus usos. Los cafetales presentaron cinco estratos de vegetación. Se encontraron 77 especies leñosas, la mayoría de uso maderable (densidad promedio de 371.4 árboles por hectárea). Noventa por ciento fueron especies nativas (40% de la flora local), el porcentaje restante fueron especies introducidas, principalmente árboles o arbustos frutales. La distribución diamétrica se asemeja a la distribución típica de bosques secundarios. El Análisis de Coordenadas Principales distinguió cuatro grupos de parcelas por la presencia de Inga, sin embargo las parcelas son diversas. No hubo diferencias en equitabilidad entre grupos y tampoco en rendimientos de café. Los rendimientos fueron de 835 g de café pergamino por arbusto o aproximadamente 1668 Kg por hectárea. Se discute el importante papel del café de sombra como refugio de plantas leñosas y como hábitats para fauna asociada así como la oportunidad de los productores de participar en el nuevo mercado de café amable con la biodiversidad.
References
Aguilar-Ortíz, F. 1982. Estudio ecológico de las aves del cafetal, p. 103-128. In E. Jiménez-Avila & A. Gómez-Pompa (eds.). Estudios ecológicos en el sistema cafetalero. CECSA, México. [ Links ]
Anderson, M.C. 1966. Some problems of the simple characterizationof the light climate in plant communities,p. 77-90. In G.C. Evans (ed.). Light as an ecologicalfactor. British Ecol. Soc. Symposium,Blackwell Scientific Pub., Oxford, U.K. Brash, A.R. 1987. The history of avian extinctions and forest conversion on Puerto Rico. Biol. Conserv. 39: 97-111. [ Links ] Breedlove, D.E. 1981. Flora of Chiapas, Part I: Introduction of the flora of Chiapas. The California Acad. of Sci., San Francisco, California. 35 p. [ Links ]
Breedlove, D.E. 1986. Listados florísticos de México. IV. Flora de Chiapas. Universidad Nacional Autónoma de México, Instituto de Biología, México, D.F. 246 p. [ Links ]
Brown, S. & A.E. Lugo. 1994. Rehabilitation of tropical lands: Akey to sustaining development. Restor. Ecol. 2: 97-111.
Del Amo, S., A.V. Cárdenas & A.L. Anaya. 1992. Manual de actividades de conservación y recuperación de especies para los comités municipales, Chiapas. Gobierno del Estado de Chiapas, Tuxtla Gutiérrez, Chiapas, México. 174 p. [ Links ]
Estrada, A., R. Coates-Estrada & D. Merrit Jr. 1993. Bat species richness and abundance in tropical rain forest fragments and in agricultural habitats at Los Tuxtlas, México. Ecography 16: 309-318. [ Links ] Everitt, B.S. & G. Dunn. 1991. Applied multivariate data analysis. Edward Arnold, New York. 304 p. [ Links ] García, E. 1988. Modificaciones al sistema de clasificación climática de Köppen. México. 217 p. [ Links ] Greenberg, R., P. Bichier, A.C. Angon & R. Reitsma. 1997a. Bird populations in shade and sun coffee plantations in central Guatemala. Conserv. Biol. 11: 448-459. Greenberg, R., P. Bichier & J. Sterling. 1997b. Bird populations in rustic and planted shade coffee plantations of Eastern Chiapas, México. Biotropica 29: 501-514. Hansen, A.J., T.A. Spies, F.J. Swanson& J.L. Ohmann. 1991. Conserving biodiversity in managed forests. BioScience 41: 382-392. Hutchinson, I.D., 1988. Points of departure for silviculture in humid tropical forests. Commonw. For. Rev. 67: 223-230. Ibarra-Nuñez, G. 1990. Los artrópodos asociados a cafetos en un cafetal mixto del Soconusco, Chiapas, México. Variedad y abundancia. Folia Entomol. Mex. 79: 207-231. Jiménez-Avila, E.& V.P. Martínez. 1979. Estudios ecológicos del agroecosistema cafetalero. II. Producción de materia orgánica en diferentes tipos de estructura. Biótica 4: 109-126. [ Links ] Lugo, A. 1995. Management of tropical biodiversity. Ecol. Appl. 5: 956-961. Miranda, F. 1953. La vegetación de Chiapas. Ediciones del Gobierno del Estado de Chiapas, Tuxtla Gutiérrez, Chiapas, Mexico. 425 p. [ Links ] Moguel, P. & V.M. Toledo. 1999. Biodiversity conservation in traditional coffee systems of Mexico. Conserv. Biol. 13: 1-11. Mueller-Dumbois D. & H. Ellenberg. 1974. Aims and methods of vegetation ecology. Wiley, New York. 499 p. Nir, M., A. 1988. The survivors: Orchids on a Puerto Rican coffee finca. Amer. Orchid Soc. Bull. 57: 989-995. Perfecto, I. & R. Snelling. 1995. Biodiversity and the transformation of a tropical agroecosystem: Ants in coffee plantations. Ecol. Appl. 5: 1084-1097. Perfecto I., J. Vandermeer, P. Hanson & V. Cartin. 1997. Arthropod biodiversity loss and the transformation of a tropical agroecosystem. Biodiv. Conserv. 6: 935-945. Perfecto, I. & J. Vandermeer. 1994. Understanding biodiversity loss in agroecosystems: Reduction of ant diversity resulting from transformation of the coffee ecosystem in Costa Rica. Entomol. (Trends Agril. Sci.) 2: 7-13. Perfecto, I., R. Rice, R. Greenberg & M.E. Van der Voort. 1996. Shade coffee: A disappearing refuge for biodiversity. BioScience 46: 598-608. Pimentel, D., U. Stachow, D.A. Takacs, H.W. Brubaker, A.R. Dumas, J.J. Meaney, J.A.S. ONeil, D.E. Onsi & D.B. Corzilius. 1992. Conserving biological diversity in agricultural/foresty systems. BioScience 42: 354-362. Purata, S. & J. Meave. 1993. Agroecosystems as an alternative for biodiversity conservation of forest remnants in fragmented landscapes, p. 9. In Smithsonian Migratory Bird Center (ed.) Symposium Abstracts. Forest remnants in the tropical landscapes: Benefits and policy implications. Smith. Migrat. Bird Center, Washington D.C. Roxburgh, J.R. & D. Kelly. 1995. Uses and limitations of hemispherical photography for estimating forest light environments. N.Z. J. Ecol. 19: 213-217. Rzedowski, J. 1978. Vegetación de México. Limusa, México, D.F. 432 p. Sneath, P.H.A. & R.R. Sokal. 1973. Numerical taxonomy. Freeman, San Francisco. 573 p. [ Links ] Soto-Pinto, L., I. Perfecto, J. Castillo, H.& J. Caballero-Nieto. 2000. Shade effect on coffee production at the northern tzeltal zone of the state of Chiapas, Mexico. Agric. Ecosyst. Environ. 80 (2000): 61-69. Steege, H., 1996. Hemiphot, Steege-Tropenbos. University of Wageningen, Netherlands. 43 p. [ Links ] Torres, J.A. 1984. Diversity and distribution of ant communities in Puerto Rico. Biotropica 16: 296-303. Whittaker, R.H. 1975. Communitites and ecosystems. MacMillan, New York. 385 p. [ Links ]

