










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.49 n.3-4 San José Dec. 2001
Crecimiento y madurez sexual de una población de Saccostrea palmula
(Mollusca: Bivalvia), Costa Rica
Jorge H. Cabrera Peña, Maurizio Protti Quesada, Mario Urriola Hernández y Osvaldo Sáenz Vargas.
Laboratorio de Recursos Naturales y Vida Silvestre (LARNAVISI), Escuela de Ciencias Biológicas, Universidad Nacional; Heredia 86-3000, Fax (506) 2376427, Costa Rica, jcabrer@samara.una.ac.cr o mprotti@samara.una.ac.cr
Recibido 27-X-2000 Corregido 30-III-2001 Aceptado 6-IV-2001
Abstract
Growth and sexual maturity in Saccostrea palmula were studied between May 1998 and May 1999, in Morales estuary, Punta Morales, Puntarenas, Costa Rica (n=540 individuals according to sample size equation). The weight-length relationship for the population was Pt = 2.13 x 10-2 Lt1.6602 and von Bertalanffy growth function was Lt = 68.2 (1- e -0.1577 t). The mean sex ratio was 1.00 male: 0.92 females with sexually mature individuals found throughout the year. Spawning activity was highest between November and January. The mean condition index (IC = PCF/PV x 100) for the population was 121.53 ± 2.24 and the mean meat yield was 10.32 ± 1.64 %.
Key words: Saccostrea palmula, oyster, growth, sexual maturity, condition index, Costa Rica.
Saccostrea palmula (Dolfuss y Dautzenberg, 1920), conocida comúnmente como ostión u ostra, se distribuye desde la laguna de San Ignacio, Baja California hasta Panamá, encontrándose también en Islas Galápagos, Ecuador e Isla del Coco, Costa Rica. Es una especie relativamente abundante en las zonas supralitoral y mesolitoral superior en rocas expuestas y raíces de mangle, presenta una gran variabilidad morfológica en la Provincia Panámica (Cruz y Jiménez 1994, Jiménez 1994) y es una especie intensivamente explotada y sometida a cultivos experimentales en México (Baqueiro 1984).
Las especies de la subfamilia Ostreinae en Centroamérica que han sido estudiadas son Crassostrea rhizophorae, por sus posibilidades de ser cultivada en forma exitosa a gran escala (Pacheco et al. 1983, Cabrera et al. 1983, Alfaro et al. 1985, Quesada et al. 1985, Madrigal et al. 1985a , b, Alarcón y Zamora 1993) y C. columbiensis (Caballero et al. 1997), por su factibilidad de ser explotada en su medio.
En Costa Rica existe una variada y abundante fauna malacológica con importancia comercial como alimento humano rico en proteínas (Cabrera et al. 1983); sin embargo, la producción actual de moluscos está basada en la explotación de los bancos naturales y la poca actividad en acuicultura está aún en etapa preliminar.
A la fecha no existen estudios sobre aspectos biológicos ni biométrico en S. palmula en Centroamérica, por lo que este trabajo tiene por objetivo, determinar el crecimiento, biometría, madurez sexual, rendimiento e índice de condición de esta especie en el manglar del estero Morales, Punta Morales, Puntarenas, Costa Rica.
Materiales y métodos
Se determinó que el número de individuos a extraer mensualmente fue de 36 y 52 (p < 0.05) según el método bimuestreal de Stein (Azorín 1972), para un total de 540 ejemplares de S. palmula, rerecolectados entre mayo 1998 a mayo 1999, por muestreo aleatorio simple en el área de manglar del estuario del estero Morales, Punta Morales, Puntarenas, Costa Rica (10° 4' N y 85° 58' W). A cada ejemplar se le midió: longitud total (Lt), altura (A) y espesor (E) con una precisión de 0.01 mm y peso total (Pt), peso carne fresca (Pcf) y peso valvas (Pv) con una precisión de 0.05 g, según lo recomendado por Cáceres et al. (1992) y Cruz (1989). El sexo y el estado de madurez sexual se determinó por frotis de gónada y observación macro y microscópica, según la tabla de madurez sexual de Lucas (1965). Los individuos fueron agrupados arbitrariamente en cinco clases de tallas: T1 (< 14.00 mm), T2 (14.01-27.25 mm), T3 (27.26-40.50mm), T4 (40.51-53.76 mm) y T5 (53.77-67.01 mm).
A los datos se les aplicó un análisis de estadística descriptiva, distribución de tallas y sexos y distribución de estadios de madurez sexual por mes, calculándose el índice de condición (IC = Pcf/Pv x 100) y el rendimiento (R = Pcf / Pt x 100) (Cabrera et al. 1983). La relación longitud total-peso total (Pt = a Lt b) se determinó según Ricker (1975) y la crecimiento según la ecuación de von Bertalanffy (Gayanilo et al. 1989).
Resultados
El peso carne fresca (Pcf), peso valva (Pv), espesor (E) y peso total (Pt) fueron los parámetros que presentaron las mayores desviaciones estándar (Cuadro 1), en relación con el respectivo valor promedio. Los coeficientes de correlación para las variables biométricas consideradas fluctuaron entre 0.7959 y 0.9854 y fueron significativamente altos, a un valor crítico de ± 0.55108, a un p < 0.05, para las variables dependientes.
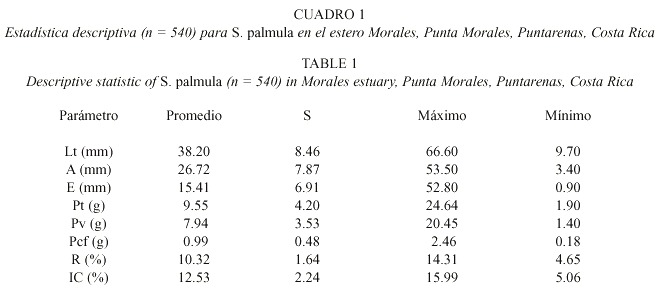
La distribución de sexos por clase de talla (Fig.1) reveló que los machos son más abundantes en las clases 2 y 3; las hembras fueron más abundantes en la clase 4 y 5 y el grupo indeterminados como el sector predominante en la clase 1. La estructura poblacional por tallas fue de un 78.46 % de individuos entre 27.26 y 53.76 mm, mientras que por sexos ésta estuvo compuesta por 13.57 % de indeterminados, 44.88 % de machos y 41.57 % de hembras.
Punta Morales, Puntarenas, Costa Rica.
Fig. 1. Sex and size group distribution of S. palmula in Morales estuary,
Punta Morales, Puntarenas, Costa Rica.
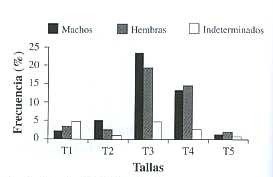
La distribución de sexos por mes (Fig. 2), evidenció a los machos como grupo dominante en mayo, agosto y noviembre 1998 y febrero y abril 1999; mientras que las hembras dominaron el resto de los meses a excepción de octubre 1998, donde los porcentajes fueron similares en ambos sexos. El grupo indeterminados se presentó durante todo el periodo de estudio sin llegar a superar el 20 % del la población total mensual. La proporción sexual promedio para la población durante el periodo de estudio fue de 1.00 machos: 0.92 hembra.
estudio en el estero Morales, Punta Morales, Puntarenas, Costa Rica.
Fig. 2. Distribution of males, females and indeterminates of S. palmula during the study in
Morales estuary, Punta Morales, Puntarenas, Costa Rica.
La relación Lt-Pt para hembras se rige por la ecuación Pt = 2.89 x 10-2 Lt1.5878, con un coeficiente de correlación (r) de 0.6653 (p < 0.05). Para los machos la ecuación fue Pt = 8.1 x 10-3 Lt 1.9068 (r = 0.6297; p < 0.05); y para la población total Pt = 2.13 x 10-2 Lt1.6602 (r = 0.6316; p < 0.05). La ecuación de crecimiento de Von Bertalanffy para S. palmula en el área de estudio fue Lt = 68.2 (1- e -0.1577 t), lo que indica que la especie presenta una longevidad de 23.8 meses.
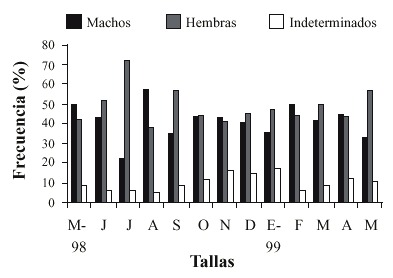
La variación mensual del desarrollo gonadal de S. palmula (Fig. 3) mostró individuos en los diferentes estadios de maduración sexual durante todos los meses, con un mayor incremento de madurez entre julio y septiembre y dos picos máximos de desove en noviembre y enero, con un promedio de desove mensual de 17.04 ± 6.78 % de la población reproductivamente activa.
Puntarenas, Costa Rica.
Fig. 3. Monthly variation of the gonadal development of S. palmula in Morales estuary, Punta Morales,
Puntarenas, Costa Rica.
El índice de condición promedio fue de 12.53 ± 2.24 % y el mayor correspondió a ejemplares de tallas entre 53.77 y 67.01 mm (13.10 ± 1.98 %) (Cuadro 2). El rendimiento promedio para la especie fue de 10.32 ± 1.64 %, encontrándose el mayor valor para las tallas entre 53.77 y 67.01 mm (10.64 ± 1.50 %).

Discusión
La talla máxima registrada para S. palmula (66.60 mm de Lt) en este estudio, fue superior al informado para la misma especie por Cruz y Jiménez (1994) (45.00 mm). Estas diferencias pueden ser atribuidas a factores de exposición del substrato, corrientes y masas de aguas propias del estero Morales, coincidiendo con Caballero et al. (1997).
Con relación a otras especies comerciales, los resultados de este estudio indican un menor tamaño que Crassostrea corteziensis (150 mm) (Cruz y Jiménez 1994), C. rhizophorae (110.2 mm) (Quesada et al. 1985) y C. columbiensis (89.6 mm) (Caballero et al. 1997).
La distribución por clase de tallas difiere de lo informada por Madrigal et al. (1985a) para C. rhizophorae en el estero Vizcaya, Limón, Costa Rica, donde el 45.50 % de individuos tuvieron tallas superiores a 59.90 mm, por Vélez (1982) para C. rhizophorae en Venezuela y por Caballero et al. (1997) para C. columbiensis en estero Morales, Costa Rica, lo cual puede ser atribuido a factores de exposición del sustrato, corrientes y calidad de agua propia de la zona de estudio.
La talla mínima sexualmente reproductiva se registró en ejemplares mayores a los 10 mm de Lt; valor menor que el informado para C. rhizophorae por Pacheco et al. (1983) (> 13 mm) en el estero Vizcaya y por Nascimiento y Da Silva (1980) (20-25 mm) y para C. columbiensis por Caballero et al. (1997) (> 11 mm), en la misma zona del presente estudio. En Venezuela, Vélez (1982) encontró para C. rhizophorae tallas mínimas similares a las encontradas en esta investigación a pesar de ser especies que alcanzan tallas máximas diferentes.
La relación Lt-Pt mostró un crecimiento del tipo alométrico, lo cual coincidió con lo informado por Pacheco et al. (1983) y Quesada et al. (1985) para C. rhizophorae en el Caribe y Pacífico costarricense respectivamente y por Caballero et al. (1997) para C. columbiensis en la misma zona de este estudio. Al analizar el incremento de longitud en función del tiempo, se obtuvo una correlación inversa (p< 0.05), lo cual concordó con los resultados de Brousseau (1984), quien informó que los moluscos bivalvos en zonas tropicales reducen su incremento en la tasa de crecimiento con el tiempo, debido a la compensación metabólica y a la poca variabilidad estacional.
El hecho que los machos predominen en las tallas menores a 40.5 mm coincidió con lo informado para C. rhizophorae por Nascimento y Da Silva (1980) en Brasil, por Vélez (1982) en Venezuela, por Alarcón y Zamora (1993) en el Caribe de Costa Rica y para C. columbiensis por Caballero et al. (1997) en el estero Morales.
Se encontraron individuos maduros y desovados durante todo el año, lo que coincidió con lo informado por Giese (1959) y Villalobos (1980), quienes señalan que en los trópicos, los invertebrados se caracterizan por tener desoves prolongados a lo largo del año y, por Caballero et al. (1997) para C. columbiensis en estero Morales; pero difiere de lo encontrado por Alarcón y Zamora (1993) para C. rhizophorae en el estero Vizcaya en el Caribe de Costa Rica, ya que en este estero, contrario a lo observado en este estudio, las ostras no se reproducen durante todo el año, debido a las fluctuaciones en la salinidad entre la época seca y la lluviosa.
El rendimiento promedio para S. palmula fue de 10.32 ± 1.64 %, valor menor a los encontrados por Squires et al. (1978) para Anadara tuberculosa en Colombia (15 %) y por Cruz y Palacios (1983) en Costa Rica (ámbito entre 16.31 y 20.34 %); por Cabrera et al. (1983) para C. rhizophorae (18.17 %) y por Cabrera et al. (1995) para Modiolus capax en Playa Ocotal (27,20%) y semejante al mayor valor informado por Caballero et al. (1997) para C. columbiensis. (10.37%).
Los mayores índices de condición obtenidos en S. palmula, corresponden a individuos mayores a 14.01 mm y menores a 27.25 mm (12.68 ± 1.69 %) y mayores a 53.77 mm (13.1 ± 1.98 %) lo que puede ser atribuido a que las ostras jóvenes o las de tallas mayores fueron usualmente planas y de poco espacio intervalvar. Esto coincidió con lo informado para C. rhizophorae por Nascimento y Da Silva (1980) y Cabrera et al. (1983) y para C. columbiensis por Caballero et al. (1997).
Agradecimientos
Se agradece a Tatiana Láscaris, Universidad Nacional por el apoyo brindado para realizar este trabajo en la Estación de Ciencias Marinas de Punta Morales (ECMAR), así como, a los revisores anónimos por sus comentarios y sugerencias al manuscrito.
Resumen
Se determinó la estructura de la población, crecimiento, madurez sexual, índice de condición y rendimiento de Saccostrea palmula en el estero Morales, Punta Morales, Puntarenas, Costa Rica, entre mayo de 1998 y mayo de 1999. La longitud total y peso total máximo encontrado fue de 66.66 mm y 24.64 g respectivamente. El crecimiento para la especie de acuerdo a la ecuación de von Bertalanffy, se rigió por Lt =68.2 (1- e -0.1577 t). La relación longitud total–peso total para la población indicó un crecimiento del tipo alométrico y se rigió por la ecuación Pt = 2.13 x 10-2 Lt1.6602. La proporción sexual fue de 1.00 macho: 0.92 hembras, con una talla mínima reproductiva en ejemplares mayores a los 10 mm de Lt y una actividad reproductiva durante todo el año. El mayor desove ocurre entre noviembre y enero. El índice de condición promedio para la población fue de 12.53 ± 2.24 % y el rendimiento de 10.32 ± 1.64 %.
Referencias
Alarcón, F. & E. Zamora. 1993. Ciclo de maduración sexual y hermafroditismo en las poblaciones de Crassostrea rhizophorae (Guilding, 1828) en estero Negro y estero Vizcaya, Limón, Costa Rica. Simposio Investigación Acuícola en Centroamérica, Heredia, Costa Rica. p. 19-36. [ Links ]
Cabrera, J., E. Zamora & O. Pacheco. 1983. Determinación del tamaño comercial de la ostra de manglar, Crassostrea rhizophorae (Guilding, 1828) en sistema de cultivo suspendido en estero Vizcaya, Limón, Costa Rica. Rev. Biol. Trop. 31: 257-162
[ Links ]Cabrera, J., R.A. Cruz, Y. Solano & M. Protti. 1995. Biometría de Modiolus capax (Bivalvia: Mytilidae) en Playa Ocotal, Guanacaste, Costa Rica. Rev. Biol. Trop. 43: 173-176.
[ Links ]Cáceres, C., C. Ruiz & D. Ramírez. 1992. Experimental collection of pearl oyster, Pinctada mazatlanica and Pteria sterna, spat on a filament substrate. J. World Aquacult. Soc. 23: 232-240.
[ Links ]Cruz, R.A. 1989. Variación mensual de la condición, composición bioquímica (proximal) y ciclo reproductivo del mejillón Mytella guayanensis (Bivalvia: Mytilidae). Tesis de grado, Escuela de Ciencias Biológicas, Universidad Nacional, Heredia, Costa Rica. 66 p.
[ Links ]Cruz, R. & J.A. Palacios. 1983. Biometría del molusco Anadara tuberculosa (Pelecypoda: Arcidae) en Punta Morales, Puntarenas, Costa Rica. Rev. Biol. Trop. 31: 175-179.
[ Links ]Cruz, R.A. & J.A. Jiménez. 1994. Moluscos asociados a las áreas de manglar de la costa Pacífica de América Central. Fundación UNA, Heredia, Costa Rica. 182 p.
[ Links ]Giese, A.C. 1959. Comparative physiology : Annual reproductive cycle of marine invertebrates. Ann. Rev. Physiol. 21: 547-576.
[ Links ]Gayanilo, F.C., J. Soriano & D. Pauly. 1989. A draf guide to Complet ELEFAN. INCLARM software 2.70. International Center for Living Aquatic Resource Management, Manila. Contribution. 435: 70 p.
[ Links ]Jiménez, J.A. 1994. Los manglares del Pacífico de Centroamérica. EFUNA. 336 p.
[ Links ]Lucas, A. 1965. Recherche sur la sexualitè des mollusques bivalves. Tr. Rennes: 135 p.
[ Links ]Madrigal, E., J. Alfaro, R. Quesada, O. Pacheco & E. Zamora. 1985a. Estructura de la población y distribución de tallas del ostión de manglar (Crassostrea rhizophorae Guilding, 1828) en el estero Vizcaya, Limón, Costa Rica. Rev. Biol. Trop. 33: 61-62
[ Links ]Madrigal, E., O. Pacheco, E. Zamora, R. Quesada & J. Alfaro. 1985b. Tasa de filtración del ostión de manglar (Crassostrea rhizophorae Guilding, 1828), a diferentes salinidades y temperatura. Rev. Biol. Trop. 33: 77-79.
[ Links ]Nascimento, I.A. & E.M. Da Silva. 1980. Desenvolvimento da gónada primaria em ostras de mangue Crassostrea rhizophorae idade e tamanho mínimo de maturacao sexual. Cien. Cult. 32: 735-742.
[ Links ]Pacheco, O., J. Cabrera & E. Zamora. 1983. Crecimiento y madurez sexual de Crassostrea rhizophorae (Guilding, 1828) cultivada en sistema suspendido en estero Vizcaya, Limón, Costa Rica. Rev. Biol. Trop. 31: 277-282.
[ Links ]Quesada, R., E. Madrigal, J. Alfaro, O. Pacheco & E. Zamora. 1985. Crecimiento y supervivencia del ostión de manglar (Crassostrea rhizophorae Guilding, 1828) trasladado de estero Vizcaya, costa del Caribe a estanques de cultivo de camarones en Chomes, Costa Pacífica de Costa Rica. Rev. Biol. Trop. 33: 7-12
[ Links ]Ricker, W.E. 1975. Computation and interpretation of biological statistic of fishes population. Bull. Fish. Res. Bd., Canadá. 382 p.
[ Links ]Squires, M., M. Estevez, O. Barona & O. Mora. 1978. Mangrove cockles, Anadara sp. (Mollusca: Bivalvia) of the Pacific coast of Colombia. Veliger 18: 57-68.
[ Links ]Vélez, A. 1982. Hermafroditismo en la ostra de mangle Crassostrea rhizophorae. Bol. Inst. Oceanogr. Univ. Oriente. 21: 129-132.
[ Links ]Villalobos, C.R. 1980. Variation in population structure in genus Tetratlita (Crustacea: Cirripedia) between temperate and tropical populations. IV The age structure of T. stalactifera and concluding remarks. Rev. Biol. Trop. 28: 353-359. [ Links ]

