










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.51 n.3-4 San José Sep. 2003
Pacific of Costa Rica (Serpentes: Viperidae)
William W. Lamar 1 & Mahmood Sasa 2
1 College of Sciences, University of Texas, 3900 University Blvd., Tyler, Texas 75799, USA.
2 Instituto Clodomiro Picado, Universidad de Costa Rica. San José, Costa Rica, and Organization for Tropical Studies, San José, Costa Rica.
Received 12-IV-2003. Corrected 29-VIII-2003. Accepted 01-IX-2003.
Abstract
A new species of terrestrial pitviper, Porthidium porrasi, is described from mesophytic forests of the Península de Osa and surrounding area of the Pacific versant of southwestern Costa Rica. It is most similar to P. nasutum and is characterized by a pattern of bands, persistence of the juvenile tail color in adults, and a high number of dorsal scales. Analysis of mtDNA sequences confirms its distinction from P. nasutum. The existence of this species reinforces the notion of elevated herpetofaunal endemism in southwestern Costa Rica.
Key words: Porthidium porrasi, Porthidium nasutum, Osa Peninsula, Costa Rica, Central America.
The pitviper genus Porthidium has undergone several revisions since its conception by Cope (1871). The genus has included two to 17 species of terrestrial snakes, most of them less than a meter in total length, and most with middle American distributions (Amaral 1927, 1929, 1944, Boulenger 1896, Campbell and Lamar 1989, Cope 1871, Dunn 1928). They share cryptic patterns and colors, and a sharply defined canthus rostralis, presumably adaptations necessary for the sedentary lifestyle of ambush predators (Campbell and Lamar 1989).
Gutberlet (1998) placed Porthidium melanurum (Muller 1923) into Ophryacus. The genus Bothrocophias now includes P. hyoprora (Gutberlet and Campbell 2001). Recently, phylogenetic relationships among the remaining species have been reviewed using morphological (Werman 1992, 1999), and molecular (Parkinson 1999, Parkinson et al. 2002) characters, and analyzed with powerful new computational algorithms. Porthidium (sensu lato) comprises three genera: the jumping pitvipers, genus Atropoides (Werman 1992); the montane pitvipers, genus Cerrophidion (Campbell and Lamar 1992); and the hognosed pitvipers, genus Porthidium (Campbell and Lamar 1989, in press). Despite the considerable morphological differences, the three genera form a monophyletic clade that originated and evolved in Central America (Savage 2002).
Porthidium includes eight species of terrestrial snakes ranging collectively from Mexico to Ecuador. In addition to a well defined canthus rostralis, members of Porthidium possess a rostral that is higher than it is wide, and the snout is usually attenuate and may be elevated either moderately or greatly. They are terrestrial, less than a meter in overall length, are medium to moderately stout in build, and inhabit xeric or transitional forest (Lawson 1997). The widely distributed P. nasutum is a notable exception, being restricted to lowland rainforest and lower montane wet forest. It ranges from Chiapas, Mexico, to western Colombia and Ecuador (Wilson and McCranie 1984, Campbell and Lamar 1989). More than twenty years field experience along the Caribbean versant of lower Central America leads us to conclude that P. nasutum is one of the most abundant snakes in that region, and this was recently supported by Castoe (2002). Another species that ranges into mesic forest is P. lansbergii, but it is likely a composite species as currently recognized, and the distribution in Central America is restricted to Panama. In Costa Rica the endemic P. volcanicum is also found in mesic forest (Solórzano 1994), and we herein describe another species, an inhabitant of the isolated rainforests in the southwestern portion of that country. Like its closest relative, P. nasutum, it is rather stout and has a greatly elevated snout.
Specimens used in the descriptions or examined for comparisons are housed in the Instituto Clodomiro Picado, Universidad de Costa Rica (ICP), in the Museo de Zoología, Universidad de Costa Rica (UCR), or in the Vertebrate Collection, University of Texas at Arlington (UTA) (see Appendix 1).
Measurements were made to the nearest 0.1 mm with dial calipers. Twenty-nine morphological characters, including scale counts, head shape, and color pattern commonly used in taxonomic work in pitvipers (Sasa 2002) were recorded. Measurements and scale counts follow Peters (1964): 1) head length; 2) head width; 3) snout length; 4) interorbital length; 5) rostral height, 6) rostral width, 7) supralabial scales; 8) infralabial scales; 9) preocular scales; 10) postocular scales; 11) interocular scales; 12) canthal scales; 13) post-canthals (marginal head scales between the canthal and supraocular); 14) intercanthal scales; 15) foveals; 16) internasals (separated or not); 17) ventrals (Dowling 1951); 18) preventrals; 19) subcaudals; 20) dorsal scale rows; 21) loreals; 22) interrictals (supracephalic scales between last supralabials); 23) nasofrontals (scales anterior to and including the interoculars, Porras et al. 1981); 24) oculabials (rows of scales between suboculars and supralabials), 25) nasorostrals (the scales between nasals and rostrals); 26) dorsal blotches (banded or fragmented); 27) number of dorsal blotches, 28) color of tail tip; 29) Presence of vertebral line. Dorsal scales rows were counted one head length posterior to the interrictals, at midbody, and at the vent (Savage 2002).
In order to evaluate molecular divergence of the new species and other members of the Porthidium clade, total cellular DNA was extracted from the skins of four individuals from Golfito, and Rincón de Osa (Puntarenas Province, Costa Rica) using standard phenolchloroform- isoamyl alcohol methods (Hillis et al. 1996). We also extracted DNA from P. nasutum from Guatemala (UTA R-46138, 46909) and Costa Rica (ASL-068).
The polymerase chain reaction (PCR) was used to amplify ~740 bp fragment of the cytochrome b gene (cyt b) following the protocols of Chippindale et al. (1998) The amplification primers were Gludg:
(5-TGACTTGAARAACCAYCGTTG-3) and ATRCB3:
(5-TGAGAAGTTTTCYGGGTCRTT-3) (Palumbi 1996).
Sequencing was performed using a LICOR© 4200 dual–laser long-read automated sequencer. Sequencing primers were the same as PCR primers (end-labeled in automated sequencing). We used the Sequiterm Excel© kit (Epicentre Technologies), reducing the manufactures reaction volumes to one fourth, and ran them under these conditions: current of 35 mA, and a voltage set to 3000 V for ca. 7 hours. Electropherograms were analyzed for sequence ambiguities and mismatch, and the sequences aligned with a sequence of Crotalus viridis (cytochrome b, GenBank accession number: U65377) using Sequecher©.
Phylogenetic analyses were conducted using PAUP* (Swofford, version 4.0b1-b2; Sinauer Associates), under two optimality criteria: maximum parsimony (MP) and maximum likelihood (ML). For parsimony analyses, changes among character states were differentially weighted using step matrices (Swofford et al. 1996).
Because the methods for phylogenetic inference depend on their underlying sequence divergence model, 10 models were examined to find the one that best fits the data using ModelTest (version 2.1, Posada and Crandall 1998). The models are: Jukes-Cantor, Kimura 2 parameter, Tamura-Nei equal frequencies, Kimura 3 parameter, SYM, Felsenstein81, Hasegawa-Kishino-Yano, Tamura-Nei, Kimura 3 parameter unequeal base frequencies, and General time reversible (see references in Posada and Crandall 1998). These models were tested under four different schemes of among-site variation: 1) equal rates among all sites, 2) a proportion of sites assumed invariant while equal rates are assumed for variable sites (I), 3) rates at all sites assumed to follow a gamma distribution (G), 4) a combination of I + G (some sites invariable, those variable assume gamma distribution). The resulting combination of 40 models allowed for testing four null hypotheses on our data: equal base frequencies, equal rates of transitions and transversions, rates equal among sites, and no invariable sites. Because all models are interrelated, ModelTest incorporates a likelihood ratio test to evaluate the goodness of fit of the data to the models. For ML analyses implemented via PAUP*, input order was randomized and global swapping across all nodes was implemented (using the global option). The ts:tv ratio was estimated empirically from the sequences. In all analyses, shortest trees were obtained using the heuristic search option with stepwise addition and 100 random taxon-sequence addition replicates in each search. Nodal support was assessed by conducting 100 heuristic non-parametric bootstrap replicates (Hillis and Bull 1993).
Results
Porthidium porrasi sp. nov.
Holotype: A male 276 mm SVL, UTA R-30829 collected at Drake Bay, Península de Osa, Puntarenas Province, Costa Rica, on 10 July 1990 (Fig. 1 and 2).
Paratypes. UCR 3310, adult female (261 mm SVL); UCR 3359, adult male (341 mm SVL); ICP 1131, adult female (338 mm SVL) from Rincón de Osa, Península de Osa, Puntarenas Province, Costa Rica. UCR 11479, juvenile male (173 mm SVL) from Golfito, Puntarenas Province, Costa Rica.

Diagnosis: Porthidium porrasi may be distinguished from all congeners by a combination of the following features: snout extremely elevated, tail tip white in both juveniles and adults, ventrals fewer than 145, and usually 25-27 midbody dorsal scale rows.
Comparison with other species: Porthidium porrasi is not sympatric with any congeners, but its range closely approaches those of P. ophryomegas, P. nasutum, and P. volcanicum. From P. ophryomegas it may be distinguished by the presence of only one canthal scale per side; from P. volcanicum it may be distinguished by its elevated snout, banded pattern, white tail tip, and lower number of ventrals (fewer than 155); from P. nasutum it may be distinguished by usually possessing 25-27 dorsal scale rows (vs 23), a white tail tip in adults, and a banded pattern rather than juxtaposed blotches.
Description of the Holotype: An adult male, 305.7 mm in total length; tail length 29.7 mm, comprising 9.7% of total length; head length 23.12 mm from front face of rostral to posterior end of mandible; head width 13.83 mm at broadest point (posterior end of mandible); snout elevated, 6.68 mm in length (from front edge of supraocular to rostral); rostral greatly elevated, about 1.65 times higher than broad; internasals 1.5 times longer than wide, not separated; nostrils small, dorsally oriented; one pre-ocular; three postoculars; interocular scales 6; supralabials 9/10, 1 contacting anterior nasal, 4 and 5 are the largest; mental small, 1.1 times broader than long; canthal enlarged; no post canthal, five to six intercanthal scales; internasals in contact; dorsal scales keeled and disposed in 23 rows in neck, 25 at midbody and reduced to 19 one head length from vent; dorsal scales in 6 rows at level of tenth subcaudal; no apical pits apparent; ventrals 143; two preventrals; anal not divided; 30 subcaudals; 24 rows of interrictals; 17 nasofrontals; one row of oculabials between suboculars and supralabials.
Color of holotype: In life the color is orange tan. The pattern consisted of 18 dorsal blotches, the anterior nine are juxtaposed at each side of vertebral line, the others form bands; vertebral line is pale, and almost 38% of the tail is cream including the tip. In preservative (ethanol after formaldehyde) color is dull brown, with darker dorsal blotches, and white tail tip. Chin heavily pigmented. The vertebral line persists after preservative.
Natural History: Porthidium porrasi inhabits mesic forest where it lies coiled by day in leaf litter. One specimen was coiled on a small log by day, and another was crawling slowly on a forest trail by night. Captives feed readily on anoline lizards and rodents. A female (UTA R-30830) from Bahía Drake on the Osa Peninsula gave birth to six young (UTA R-30831-36) ranging from 167 to 175 mm in total length on 24 June 1991. Another from La Gamba, near Golfito, gave birth to nine young in March.
Distribution: As currently recognized, the species is distributed in the lowland Pacific wet forest southwestern Costa Rica, including Península de Osa, and the lands surrounding the Golfo Dulce. It is found below 1000 m above sea level.
Etymology: The name porrasi is a patronym honoring Louis W. Porras, a Costa Rican born herpetologist who was the first to recognize the distinctiveness of the hognosed pitvipers inhabiting the Península de Osa. It is masculine in gender.
Variation: Total length of adult specimens examined ranged from 288 mm to 385 mm. Tail lengths of males (UCR 3359 and UCR 11479) are 44 (11.4% body length) and 22 mm (11.3%) respectively; whereas females have shorter tails: 27 (9.3%), and 39 mm (10.2%). The rostral is higher than wide in all specimens: observed ratios (height/width) were 1.17, 1.43, 1.32, and 1.21. All specimens have 10 strongly pigmented supralabials, except UCR 3310 that has nine. Eleven infralabials, usually pigmented, are present, together with 1 or 2 preoculars and similar number of postoculars. Interocular scales range from 5 to 6. Canthals are elongated and slightly pigmented. One individual (UCR 3359) lacks a postcanthal, the others possess one. Intercanthal count differs among individuals, ranging from 6 to 9 scales. In all examined specimens, internasals are not separated totally, although one scale is usually inserted
between them. Ventral scales are not different between males and females examined, as 138 and 144 were observed for both sexes. On the other hand, undivided subcaudals in males are 30 and 33, whereas in both females 26 sub-caudals were counted. Interrictals range from 25 to 29. Dorsal scales rows are consistently higher in the neck and midbody than at the vent: observed formulas are 25-25-20, 25-24- 19, 27-24-19, 25-25-19. Nasofrontals range from 23 to 29 scales. One or two oculabials were observed. The number of dorsal blotches in the body is 15 and 16, and in all specimens more than 70% of the blotches are not fragmented.
In all specimens, tip of the tail is cream, covering a third to a quarter of the overall length of the tail. Also, a cream vertebral line is present in all cases (see Fig. 2).
Divergence at the molecular level: An analysis of cyt b sequences of Porthidium porrasi and P. nasutum reveal seventy-one parsimony informative characters. Nucleotide sequences departed from the equal base frequency hypothesis, as well as from the equal rates of transitions and transversions, and equal ratio among sites hypotheses (p< 0.00001 in all cases). However, the null hypotheses of equal rate transitions and equal rate transversions were accepted (p> 0.133 in both cases). Thus, the evolutionary model that best fit the divergence of nucleotide sequences in both parameter economy and sequence divergence was Hasegawa-Kishino-Yano (1985, HKY + G).
Divergence within cyt b sequences Porthidium porrasi ranged from 0.0028 to 0.0048 substitutions per site, whereas observed divergence between sequences of P. porrasi and P. nasutum ranged from 0.049 to 0.066 substitutions per site (mean = 0.055). Parsimony analyses retrieved a single parsimony trees (215 steps, consistency index CI = 0.898, retention index RI = 0.861). In this tree, P. porrasi samples form a monophyletic group sister to the clade that includes samples from P. nasutum from Costa Rica and Guatemala (Fig. 3).
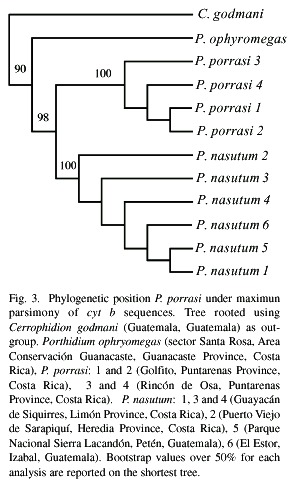
The topology retrieved under maximum parsimony analysis was not in conflict with that obtained using maximum likelihood under the HKY + G model (-ln L = 1948.759, ti/tv = 4.2082, a (gamma shape parameter) = 0.2488). Thus both methods of phylogenetic reconstruction placed Porthidium porrasi as a distinct clade sister to P. nasutum, and the observed divergences are well supported with mtDNA.
Discussion
In their analysis of P. nasutum morphological variation, Porras et al. (1981) noticed that populations from Península de Osa and adjacent areas could be distinguished from other populations of the species. However, no taxonomic changes were suggested at that time, and individuals from the southwestern populations were regarded as simple variants of the P. nasutum morphotype. Our study reveals however that, despite the extensive overlap in scale counts with other related species, P. porrasi can be distinguished from other members of the genus both at the morphological and molecular level.
Most of the diagnostic features of P. porrasi might be described as neotenic characters that are expressed early on in other related members of the group. For instance, Porras et al. (1981) stated that juveniles of P. nasutum differ from adults in that they are "noticeably brighter in color and more distinct in pattern than are the adults" (pp. 99). All adult P. porrasi collected by us are also brightly colored and banded. Juvenile P. nasutum are also characterized by the following combination: usually with head markings, pale vertebral line present, well defined dorsal blotches, and a conspicuous cream to yellowish coloration in the tip of the tail, features that are also present in juveniles of other terrestrial pitvipers (Solórzano 1990, Solórzano et al. 1998). All these characters are present in adult P. porrasi of both sexes. Thus, it seems that retention of neotenic characters was a byproduct of the process of speciation that resulted in P. porrasi. Speciation through neoteny might not be an uncommon pattern, as several examples of neotenic lineages are known among lower vertebrates (Chippindale et al. 2000).
The phylogenetic affinities of P. porrasi are not understood to this date, but it seems that the species is closely related to P. nasutum (Porras et al. 1981). To estimate time of divergence between these forms form our mDNA sequences, we employed the Brown et al. (1982) primate molecular clock based on cyt b sequences. Using the gamma-HKY model, Arbogast and Slowinski (1998) corrected the primate clock and estimated a rate of 0.0278 substitutions per site per lineage per million years (Mya). Thus, at that rate, the divergence between the P. porrasi and P. nasutum lineages might have occurred in the last 1.7 million years, coinciding with events that began during the late Pliocene. In late Pliocene and throughout the Pleistocene, repeated cooling and warming episodes affected the region. During Glacial periods, temperatures dropped so that mountain vegetation mixed with low-land. However, as Colineaux (1996) suggested, interglacial periods were probably wet periods in Middle America. Thus, isolation of the wet lowland forest in the Southwestern Pacific of Costa Rica might be produced by other factors. It seems that the uplift of the major mountain ranges in the country emphasized the rainshadow effect of the Pacific slope and fragmented the faunas at each side of the mountains. The last episodes of volcanism in the Talamancas, the southern mountain range that divides Costa Rica, coincide with the P. nasutum – P. porrasi separation timeframe (Denyer et al. 2000).
In recent years, two other species of vipers have been described from the southwestern region of Costa Rica: Porthidium volcanicum (Solórzano 1994) and Bothriechis supraciliaris (Solórzano et al. 1998). The presence of P. porrasi on the Península de Osa adds to the notion that these wet forests contain not only a great diversity, but also are highly rich in endemics.
Agknowledgements
We thank Jonathan A. Campbell and Federico Bolaños for kindly providing material for this study. Erika Deinert, Louis W. Porras, and Alejandro Solórzano made valuable comments about the manuscript. Paul T. Chippindale and Todd Castoe kindly help us with the molecular analyses. Luis Diego Gómez discussed the biogeography of the mesic forest in southern Costa Rica. The Miranda family and staff of Marenco Biological Station helped us with logistics and housing while working in Osa. This study was supported in part by Vicerrectoría de Investigación, Universidad de Costa Rica (Project 741-A1-021).
Resumen
Una nueva especie de víbora terrestre, Porthidium porrasi, es descrita de los bosques húmedos de la Península de Osa y áreas adyacentes en la vertiente Pacífica del suroeste de Costa Rica. La especie es más similar a P. nasutum y se caracteriza por la persistencia en adultos de caracteres neoténicos como patrón de bandas, línea vertebral pálida, punta de cola crema y la presencia de un número alto de filas de escamas dorsales. Los análisis de secuencias de ADN mitocondrial confirman su distinción de P. nasutum. La existencia de esta especie refuerza la noción de un fuerte endemismo de herpetofauna en el suroeste de Costa Rica.
References
Arbogast, B.S. & J.B. Slowinski. 1998. Pleistocene speciation and the mitochondrial DNA clock. Science 282: 1955. [ Links ]
Amaral, A. do. 1927. Studies of neotropical ophidia. V. Notes on Bothrops lansbergii and B. brachystoma. Bull Antivenin Inst. America 1: 22. [ Links ]
Amaral, A. do. 1929. Studies of neotropical ophidia. XII. On the Bothrops lansbergii group. Bull. Antivenin Inst. America 3: 19-27. [ Links ]
Amaral, A. do. 1944. Notas sôbre a ofiología neotrópica e brasílica. II. Sôbre Bothrops lansbergii lansbergii (Schlegel 1841); Trimeresurus dunni Hartweg and Oliver 1938; T. lansbergii annectens Schmidt 1936. Pap. Avulsos. Dep. Zool., São Paulo 5: 7-12. [ Links ]
Boulenger, G.A. 1896. Catalogue of the snakes in the British Museum (Natural History). Vol. 3. Taylor and Francis, London, 727 p. [ Links ]
Brown, W.M., E.M. Prager, A. Wang & A.C. Wilson. 1982. Mitochondrial DNA sequences of primates: Tempo and mode of evolution. J. Mol. Evol. 18: 225-239. [ Links ]
Campbell, J.A. & W.W. Lamar. 1989. The venomous reptiles of Latin America. Cornell Univ., Ithaca, New York, 425 p. [ Links ]
Campbell, J.A. & W.W. Lamar. 1992. Taxonomic status of miscellaneous Neotropical viperids, with the description of a new genus. Occas. Pap. Mus. Texas Tech. Univ. 153: 1-31. [ Links ]
Campbell, J.A. & W.W. Lamar. [in press]. The venomous reptiles of the Western Hemisphere. Vol. 2. Cornell University, Ithaca, New York. [ Links ]
Castoe, T.A. 2002. Microhabitat selection in Porthidium nasutum (Serpentes: Viperidae) in Costa Rica, with comments on ontogenetic variation. Herpetol. Rev. 33: 174-175. [ Links ]
Chippindale, P.T., L.K. Ammerman, & J.A. Campbell. 1998. Molecular approaches to phylogeny of Abronia (Anguidae: Gerrhonotinae), with emphasis on relationships in the subgenus Auriculabronia. Copeia 1998:883-892 [ Links ]
Chippindale, P.T., A.H. Price, J.J. Wiens & D.M. Hillis. 2000. Phylogenetic relationships and systematics revision of central Texas hemidactyliine plethodontid salamanders. Carnagie Museum Publications in Zoology. [ Links ]
Colinveaux, P.A. 1996. Quaternary environmental history and forest diversity in the Neotropics. pp. 359-405. In J.B.C. Jackson, A.F. Budd & A.G. Coates. Evolution and environment in tropical America. The University of Chicago, Chicago. [ Links ]
Cope, E.D. 1871. Ninth contribution to the herpetology of tropical America. Proc. Acad. Nat. Sci. Philadelphia 23: 200-224. [ Links ]
Denyer, P., G.E. Alvarado & T. Aguilar. 2000. Historia Geológica. pp. 155-167. In P. Denyer & S. Kussmaul (eds.). Geología de Costa Rica. Editorial Tecnologica de Costa Rica, San José. 513 p. [ Links ]
Dowling, H.G. 1951. A proposed standard system of counting ventrals in snakes. British J. Herpetol. 1: 97-99. [ Links ]
Dunn, E.R. 1928. Notes on Bothrops lansbergi and Bothrops ophryomegas. Bull. Antivenin Inst. America 2: 29-30. [ Links ]
Gutberlet, R.L. Jr. 1998. The phylogenetic position of the Mexican black tailed pitviper (Squamata: Viperidae: Crotalinae). Herpetologica 54: 184-206. [ Links ]
Gutberlet, R.L. Jr. & J.A. Campbell. 2001. Generic recognition for a neglected lineage of South American pitvipers (Squamata: Viperidae: Crotalinae), with a description of a new species from the Colombian Chocó. Amer. Mus. Novitates. 3316: 1-15. [ Links ]
Hasegawa, M., Kishino, K. & T. Yano. 1985. Dating the human-ape splitting by a molecular clock of mitochondrial DNA. J. Mol. Evol. 22: 160-174. [ Links ]
Hillis, D.M. & J.J. Bull. 1993. An empirical test of boot-strapping as a method for assessing confidence in phylogenetic analysis. Syst. Biol. 42: 182-192. [ Links ]
Hillis, D.M., B.K. Mable, A. Larson, S.K. Davis & E.A. Zimmer. 1996. Nucleic acids. IV. Sequencing and clonning. pp. 321-381. In D.M. Hillis, C. Moritz & B.K. Mable (eds.). Molecular Systematics. Sinauer Associates, Sunderland, Massachusetts. [ Links ]
Lawson, D. 1996/1997. Sexual dimorphism and reproduction of the pitviper Porthidium ophryomegas (Serpentes: Viperidae) in Guatemala. Rev. Biol. Trop. 44 (3)/45(1): 671-674. [ Links ]
Palumbi, S.R. 1996. Nucleic acids II: The polymerase chain reaction. pp. 205-247. In D.M. Hillis, C. Moritz & B.K. Mable (eds.). Molecular Systematics. Sinauer Associates, Sunderland, Massachusetts. [ Links ]
Parkinson, C.L. 1999. Molecular systematics and biogeographical history of pitvipers as determined by mitochondrial ribosomal DNA sequences. Copeia 1999: 576-586. [ Links ]
Parkinson, C.L., J.A. Campbell & P.T. Chippindale. 2002. Multigene phylogenetic analysis of pitvipers with comments on their biogeography. pp. 93-110. In G. W. Schuett, M. Höggren, M.E. Douglas & H.W. Greene (eds.). Biology of the Vipers. Eagle Mountain Publishing, Eagle Mountain, Utah. 580 p. [ Links ]
Peters, J.A. 1964. Dictionary of Herpetology. Hafner Publishing, New York and London. [ Links ]
Porras, L., J.R. McCranie, & L.D. Wilson. 1981. The systematics and distribution of the hognose viper Bothrops nasuta Bocourt (Serpentes: Viperidae). Tulane Stud. Zool. Bot. 22: 85-107. [ Links ]
Posada, D. & K.A. Crandall. 1998. Modeltest: Testing the model of DNA substitution. Bioinformatics 14: 817- 818. [ Links ]
Sasa, M. 2002. Morphological variation in the lancehead pitviper Bothrops asper (Garman) (Serpentes: Viperidae) from Middle America. Rev. Biol. Trop. 50(1): 259-271. [ Links ]
Savage, J.M. 2002. The amphibians and reptiles of Costa Rica. A herpetofauna between two continents, between two seas. The University of Chicago, Chicago and London, 933 p. [ Links ]
Solórzano, A. 1990. Reproduction in the pitviper Porthidium picadoi Dunn (Serpentes: Viperidae) in Costa Rica. Copeia 1990: 1154-1157. [ Links ]
Solórzano, A. 1994. Una nueva especie de serpiente venenosa terrestre del genero Porthidium (Serpentes: Viperidae), del sureste de Costa Rica. Rev. Biol. Trop. 42: 695-701. [ Links ]
Solórzano, A., L.D. Gómez, J. Monge & B. Crother. 1999 [dated 1998]. Redescription and validation of Bothriechis supraciliaris (Serpentes: Viperidae). Rev. Biol. Trop. 46: 453-62. [ Links ]
Solórzano, A., J. M. Gutiérrez & L. Cerdas. 1988. Botrhops ophryomegas Bocourt (Serpentes: Viperidae) en Costa Rica: distribución, lepidosis, variación sexual y cariotipo. Rev. Biol. Trop. 36: 187-190. [ Links ]
Swofford, D.L. 1996. PAUP*: Phylogenetic analysis using parsimony. Version 4.0. Sinauer Associates, Sunderland, Massachusetts. [ Links ]
Swofford, D.L., G.J. Olsen, P.J. Waddell & D.M. Hillis. 1996. Phylogenetic inference. pp. 407-514. In Hillis, D.M., C. Moritz & B.K. Mable (eds.). Molecular Systematics. 2nd ed. Sinauer Associates, Sunderland, Massachusetts. [ Links ]
Werman, S.D. 1992. Phylogenetic relationships of Central and South American pitvipers of the genus Bothrops (sensu lato): Cladistic analyses of biochemical and anatomical characters. pp. 21-40. In Biology of the Pitvipers, J.A. Campbell & E.D. Brodie, Jr., (eds.). Selva, Tyler, Texas. [ Links ]
Werman, S.D. 1999. Molecular phylogenetics and morphological evolution in Neotropical pitvipers: An evaluation of mitochondrial DNA sequence information and the comparative morphology of the cranium and palatomaxilary arch. Kaupia 8: 113-126. [ Links ]
Wilson, L.D. & J.R. McCranie. 1984. Bothrops nasuta. Cat. Amer. Amph. Rept. 349: 1-2. [ Links ]
Specimens examined
Porthidium porrasi – COSTA RICA: Puntarenas, Golfito (UCR 11479, personal collection); Península de Osa, Rincón de Osa (UCR 3310, UCR 3359, ICP 1131).
P. nasutum – COSTA RICA: Guayacán, Siquirres (ASL-068); Heredia, Puerto Viejo de Sarapiquí (personal collection); – GUATEMALA: Petén, La Libertad, Parque Nacional Sierra Lacandón (UTA R-46138); Izabal, Sierra de Santa Cruz, El Estor, Aldea Santa María (UTA R-46909).

