











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkEl jaguar (Panthera onca) es el felino neo tropical de mayor tamaño y el mayor depredador que habita en el neo trópico (Seymour, 1989; Sanderson et al., 2002). Pertenece al grupo de los pantheridos, que está compuesto por los cuatro felinos más grandes del mundo; el tigre (Panthera tigris), el león (Panthera leo), el leopardo (Panthera pardus) y el jaguar en América (Panthera onca) (Seymour, 1989; Garla, 2001). Su ámbito de distribución se extiende desde el norte de México hasta el sureste de Argentina (Carrillo et al., 2000a; Aranda, 2000). Cumple un importante papel ecológico en los ecosistemas, por su posición en la cúspide de la cadena trófica que convierte esta especie en un elemento esencial para mantener el equilibrio y la diversidad de los ecosistemas (Aranda, 1994; Sunquist & Sunquist, 2002). Pese al alto valor de esta especie, al igual que la mayoría de carnívoros de gran tamaño en el mundo, se encuentra amenazada o en peligro de extinción a causa de presiones antropogénicas (Ceballos et al., 2005). Por lo que el descenso en el número de presas silvestres disponibles, así como la fragmentación de hábitats y la eliminación de jaguares depredadores de animales domésticos, han mermado sus poblaciones (Sáenz & Carrillo, 2002; Sanderson et al., 2002; Hoogesteijn & Hoogesteijn, 2005; Amit et al., 2010). En lo que respecta al jaguar y otros felinos en el neo trópico su grado de amenaza es evidente (Carrillo et al., 2000b; Ceballos et al., 2005), además que sus hábitos conductuales han dificultado su estudio a lo largo de su área de distribución, conociendo muy poco de sus poblaciones (densidad, abundancia relativa, interacciones inter específicas, áreas de actividad, así como requerimientos de hábitat) (Rabinowitz & Nottigham, 1986; Rabinowitz & Zeller, 2010). Sin embargo, pese a estas dificultades se han desarrollado esfuerzos por determinar tamaños poblacionales y aspectos ecológicos con la finalidad de implementar medidas de manejo con información generada dentro y fuera de áreas protegidas (Carrillo et al., 2000b; Silver et al., 2004, Novack et al., 2005, Weckel et al., 2006; Soisalo & Cavalcanti, 2006; Salom et al., 2007). Los métodos comúnmente usados en el neo trópico para el estudio de poblaciones silvestres han sido: la telemetría para estudios de comportamiento (Schaller, 1980; Rabinowitz & Nottigham, 1986; Carrillo, 2000b); el foto trampeo para determinar abundancia absoluta y relativa (Karranth, 1995; Silver et al., 2004; Carazo, 2009) y el uso de conteo de rastros para la estimación de índices de abundancia poblacional (Chinchilla, 1997; Aranda, 2000; Carrillo et al., 2000a; Ramírez 2003). La principal limitante para el desarrollo de estas investigaciones en áreas protegidas, son los elevados costos que incurre la implementación de la telemetría y el foto trampeo (Balme et al., 2009). Por tanto es necesario implementar métodos de bajo costo y fiables para evaluar el estado de las poblaciones dentro de las área protegidas y así verificar el nivel de cumplimiento de sus objetivos de conservación. En este estudio se utilizaron índices de abundancia obtenidos a través del conteo sistemático de rastros para determinar cambios en las abundancias de felinos y sus presas en el período del 2001-2011, en una red de siete senderos dentro del Parque Nacional Santa Rosa (PNSR). Nuestra hipótesis fueron: en estadios sucesión más avanzados las poblaciones de grandes felinos van a tender a un repunte, y ese aumento va a regular la abundancia de los distintos gremios de presas potenciales en el tiempo.
Área de estudio: El Parque Nacional Santa Rosa (PNSR) (10°53'1" N, 85°46'30" O) se ubica 35 kilómetros al norte de Liberia en la provincia de Guanacaste (Boza, 1992). Tiene una superficie de 38 628 ha, compuesto por un mosaico de bosque geográficamente continuo de 163000 hectáreas. Dentro de estos bloques de vegetación algunos estuvieron dedicados a la ganadería y agricultura en el pasado (Janzen, 1988). No obstante ahora son tierras en proceso de regeneración ecológica precedidas por manejo de hábitat desde mediados de los ochentas (Janzen, 1986). El PNSR protege uno de los últimos parches de bosque seco mejor conservado en Mesoamérica, caracterizado por una estacionalidad bien marcada, donde el periodo seco se extiende de Diciembre a Abril. La media anual de temperatura oscila entre los 22 y 33 C° con una baja humedad relativa. En cuanto a la estación lluviosa se extiende de mayo a noviembre con una media anual de precipitación de 1 696 mm que va de los 900 mm a los 2 000 mm, según la Estación Automática de Santa Rosa.
Métodos
Se realizaron dos muestreos de Mayo a Septiembre en el 2001 y en el 2011, empleando el método de conteo de rastros en siete senderos dentro del PNSR para la construcción de índices de abundancia de grandes felinos y sus presas potenciales.
Registro de rastros de jaguar, otros felinos y sus presas potenciales. Para determinar los índices de abundancia relativa se realizaron 2 recorridos por mes sobre cada sendero. Recorriendo a paso lento (1 km/h) en busca de signos de presencia animal como huellas, excretas y observaciones directas (Aranda, 2000), registrando como un rastro animal cualquier signo (huella, excretas y marcas) o conjunto continuo de pistas sobre el camino.
Cálculo de abundancia relativa de jaguar, otros felinos y presas potenciales mediante conteo de rastros. Para estimar la abundancia relativa de jaguar y sus presas potenciales. Se consideraron como presas potenciales especies que han sido reportadas dentro de la dieta del jaguar en la bibliografía (Sunquist & Sunquist, 2002).
Posteriormente para el cálculo del índice de abundancia (Ec. 1), se dividió el número total de pistas registradas (fs) entre los kilómetros recorridos durante el muestreo (n), expresando los resultados como número de pistas por kilómetro (IAR).
∑ fs / n = IAR
En el análisis de datos se agruparon los IAR de las especies registradas en el 2001 y el 2011 en gremios alimenticios (Cuadro 1). Para comparar los cambios en los índices de abundancia del año 2001 y el 2011, se aplicó un análisis de varianza (ANDEVA) por gremio y por año, evaluando previamente el supuesto de homogeneidad de varianza mediante una prueba de Bartlett, y la normalidad mediante una prueba de Shapiro-Wilk, ejecutando todas las pruebas estadísticas mediante el software estadístico R 2.6.12 (R Development CoreTeam, 2011).
Resultados
Se recorrieron un total de 142,32 km en el año 2011 y 172,32 Km en el año 2001, detectando un total de 17 especies de mamíferos en ambos puntos de muestreo (Cuadro 1), observando que únicamente la zorra gris (Urocyon cinereoargenteus) no fue detectada en el muestreo del 2011.
Cuadro 1: Resultado ANDEVA para el IAR (rastros/km) por gremio alimenticio y por periodos. Parque Nacional Santa Rosa, Guanacaste, Costa Rica, 2001 y 2011

Registrando una taza de avistamiento para rastros en el 2001 de 9,005 registros/Km y 6,644 registros/Km para el 2011. En cuanto a los índices de abundancia relativa (IAR) por gremio para carnívoros y sus presas, cumplieron los supuestos de homogeneidad de varianza (Gl=3; P= 2,2 e-16) y normalidad (P= 2,2 e-16). En lo que respecta a los resultados del ANDEVA de acuerdo al criterio de información de Akaike el modelo que mostró mayor evidencia estadística fue el mod1 (CIAc 1620.5), que incluyó la interacción de los gremios por año (Cuadro 2), distando del modelo nulo (modnull) en una magnitud de ΔCIAc 76,2. Demostrando mayor evidencia estadística para afirmar diferencias evidentes entre ambos modelos (Anderson, 2002).
Cuadro 2: Registro de especies detectadas mediante conteo de rastros sobre principales senderos dentro del Parque Nacional Santa Rosa, Guanacaste, Costa Rica, 2001 y 2011
*Detectado únicamente en el 2001
El promedio de IAR de 2001 a 2011 para los felinos (Figura 1) mostró un incremento 0,425 IAR (IC: 0,348 - 0.513), 0.851 IAR (CI: 0,600 - 1,351), misma tendencia que mostró el gremio omnívoro (Fig.2) para el periodo del 2001 al 2011 donde los promedios de los IAR aumentaron 0,434 IAR (IC: 0,335-0,526), 0,912 IAR (CI: 0,642 - 1.244). El IAR del gremio frugívoro en el periodo del 2001 al 2011 indica un descenso 1.191 IAR (IC: 0,945-1,516); 0,822 IAR (CI: 0,592 - 0,908), patrón similar al que muestra el gremio herbívoro donde las medias del IAR del 2001 al 2011, tendieron a disminuir 5,456 IAR (IC: 3,966 -7.313); 3,530 IAR (CI: 2,794 - 4,320).
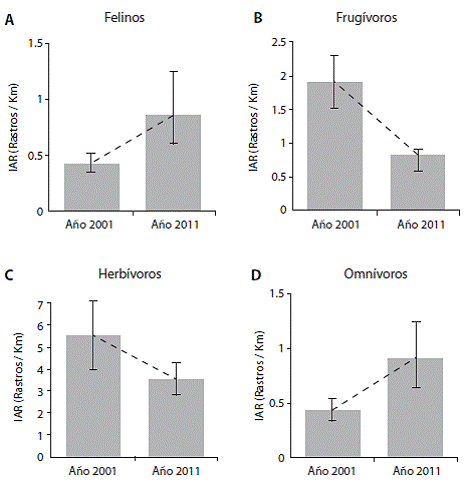
Figura 1: Grafico interacción IAR (rastros/km) por ano de acuerdo al gremio alimenticio, promedio e intervalo de confianza al 95%. Parque Nacional Santa Rosa Guanacaste, Costa Rica, 2001 y 2011.
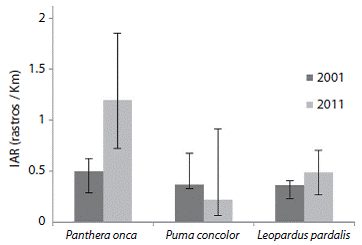
En cuanto al grupo felinos (Figura 2), el jaguar registró un incremento en el IAR promedio de 0,497 rastros/km (IC: 0,286 - 0,622) a 1,196 rastros/km (IC: 0,721 -1,853), con el puma el IAR disminuyó de 0,366 rastros/km (IC: 0,182 - 0,527) a 0,22 rastros/km (IC: 0,156 - 0,693), finalmente el IAR para ocelote aumentó de 0,359 rastros/km (IC: 0,227- 0,402) a 0,485 rastros/km (IC: 0,266 - 0,705).
Discusión
El grupo felinos (jaguar, puma y ocelote), mostró un aumento significativo en su IAR del 2001 al 2011 según datos colectados mediante conteo de rastros. Esta tendencia a principios del 2001 indica como la abundante disponibilidad de presas ha ido favorecido las condiciones necesarias para que estos carnívoros puedan costear la demanda energética que incurre la reproducción (Sandell, 1989; Karranth et al., 2004). De manera que esta estrecha relación combinada con las condiciones ecológicas existentes en el PNSR han beneficiado las poblaciones de grandes felinos. La abundancia del jaguar indica un incremento en esta especie, misma tendencia observada mediante cámaras trampa, en dos muestreos puntuales con 5 años de diferencia donde se reportó una mayor proporción en el número de individuos juveniles (Montalvo, 2012). Las abundancias de pumas y jaguares en el 2001 fueron similares, no obstante diez años después cuando la abundancia de jaguares muestra un repunte, se observa como el número de registros de puma en senderos decreció. Lo que probablemente pueda deberse a la existencia de un patrón entre estas dos especies a evadir co-específicos (Harmsen et al., 2009), ya que en sitios donde conviven ambas especies se ha demostrado que pueden permanecer evitándose mutuamente de manera temporal y espacial para impedir encuentros (Scognamillo et al., 2003). Otra posible explicación a este repunte podría ser la presencia de sitios de anidamiento de tortuga lora (Lepidochelys olivacea) y negra (Chelonia mydas), dentro del PNSR (Carrillo et al., 2009, Fonseca et al., 2009), lo que asegura una potencial presa para el jaguar inaccesible para puma. Debido a que a que el jaguar es capaz de explotar un umbral de presas mayor que el puma (Seymour, 1989; Moral-Sachetti et al., 2011), muy probablemente el aumento en la población de jaguares este limitando aún más la disponibilidad del recurso alimenticio para el puma, de manera que estén siendo marginados (Dayan & Simberloff, 1996). En cuanto a los ocelotes sus tendencias permanecieron constantes sin reportar ningún cambio estadísticamente evidente. Según Oliveira et al., (2010) esta tendencia puede ser explicada por la presencia de carnívoros mayores como el jaguar y el puma, que mantienen marginadas las poblaciones de felinos de menor tamaño. Sin embargo dentro del PNSR es muy probable que ante la ausencia de presas como el chancho de monte, felinos como el jaguar tiendan a traslapar mayormente su dieta con los pumas, lo que podría estar reduciendo su disponibilidad, desplazando la dieta de los pumas a otras presas de menor tamaño también consumidas por ocelotes. Los IAR generales para los grupos de presas potenciales no mostraron cambios estadísticamente significativos con ambos métodos. Sin embargo al agruparlos por gremios alimenticios se observó como en 2011, con respecto al 2001, los grandes felinos aumentaron, los herbívoros tendieron a disminuir y el gremio compuesto por los omnívoros reportó un incremento en sus abundancias. Siendo muy probable que el manejo de hábitat que se ha dado dentro del PNSR (Jansen, 1988) haya limitado la dominancia de especies arbóreas dispersadas por viento, lo que ha favorecido la recolonización de otras especies de árboles que ofrecen una gama de frutos y semillas carnosos (Jansen, 1983), destacando por ser más atractivos para frugívoros y herbívoros que al mismo tiempo actúan como dispersores, acelerando los procesos de regeneración y aumentando la disponibilidad de alimento para estos gremios (Ribeiro et al., 2012). Aumentando así la abundancia de herbívoros y frugívoros, por lo que en estadios más avanzados las poblaciones de carnívoros van a disponer de una mayor cantidad de presas, promoviendo condiciones idóneas para la reproducción, aumentando así la abundancia de este gremio que va a tender a regular poblaciones de herbívoros y frugívoros disminuyendo la presión sobre los organismos vegetales y liberando de esta forma espacios para organismos pertenecientes al gremio omnívoro, que es más generalista (Román, 2000; Ripple et al., 2007). Finalmente concluyendo que el método de conteo sistemático de rastros permitió darle seguimiento al dinamismo de las poblaciones en proceso de recuperación dentro del PNSR. Donde estos procesos de restauración están abriendo nuevos espacios con condiciones idóneas de alimentación y refugio para albergar nuevos depredadores clave como el jaguar. Experimentando una dinámica en donde interacciones y procesos biológicos interrumpidos por actividades antrópicas previas estén tendiendo a estados estables donde se demuestra la influencia positiva de un depredador clave como el jaguar a lo largo de la estructura de ese ecosistema (Estes et al., 2011).

